Белки теплового шока и фотосинтетическая активность хвои сосны обыкновенной в постпирогенный период

Автор: Коротаева Н.Е., Гетте И.Г., Косов И.В., Пахарькова Н.В., Боровский Г.Б.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 10, 2017 года.
Бесплатный доступ
Формирование растительного покрова Си-бири, как ранее, так и на современном этапе, происходит под воздействием лесных пожа-ров. Температурный стресс при прохождении низового пожара вызывает в растениях опре-деленные физиологические изменения и, как следствие, влияет на динамику леса. Во время прохождения лесного пожара температурный градиент в кроне очень крутой, и тепловые потоки могут существенно влиять на фото-синтетические свойства растений. Для опре-деления способности к восстановлению и со-хранению акклимационных эффектов было про-ведено полуколичественное определение со- держания белков теплового шока (heat shock proteins, Hsp) и фотосинтетической активно-сти хвои сосны обыкновенной (Pinus sylvestris L.) как критериев в оценке общего физиологиче-ского статуса в постпирогенный период. Ис-следования проводились в сосновых молодня-ках зеленомошно-разнотравной группы типов леса, располагающихся в лесостепной зоне Южной Сибири. На опытном участке в 2014 году проводилось экспериментальное контро-лируемое выжигание, имитирующее низовой пожар средней силы. Проведенные экспери-ментальные исследования выявили различия в относительном параметре замедленной флуоресценции и в содержании Hsp, свиде-тельствующие о модификации некоторых фи-зиологических процессов в растениях после экспериментального выжигания. Возможно, благодаря накоплению Hsp 70 и Hsp 101 хвоя сосны обыкновенной, которая была сформиро-вана после искусственного выжигания, обла-дает большей устойчивостью к повторному действию высоких температур.
Гипертермия, флуорес-ценция, белки теплового шока, лесные пожары
Короткий адрес: https://sciup.org/140224154
IDR: 140224154 | УДК: 574.24:582.475:630*43
Heat shock proteins and photosynthetic activity of needles of ordinary pine in post-fire period
The forest-formation process in Siberia both earlier and at the present stage occurs under the influence of forest fires. Heat stress due to fire ef-fects is exerting strong effects on plant physiology and resultant forest dynamics. During forest fires the temperature gradient in the crown layer is very steep and heat events can have major impact on photosynthetic features of plants. For the evalua-tion of ability to restoration and preservation of ac-climatization effects semi-quantitative definition of the content of proteins of thermal shock (heat shock proteins, Hsp) and photosynthetic activity of needles was carried out in ordinary pine (Pinus sylvestris L.) as criteria in the assessment of gen-eral physiological status during after fire period. The researches were conducted in pine saplings of green-motley grass group of the types of the wood settling down in the forest-steppe zone of Southern Siberia. In 2014 on experimental site testing con-trolled burning imitating local low-intensity fire was carried out. Conducted pilot studies revealed the distinctions in relative parameter of slowed-down fluorescence and in the maintenance of Hsp, testi-fying to the modification of some physiological pro-cesses in plants after experimental burning out. Perhaps, due to the accumulation of Hsp 70 and Hsp 101 the needles of ordinary pine formed after controlled burning, were more resistant to repeated high temperature stress.
Текст научной статьи Белки теплового шока и фотосинтетическая активность хвои сосны обыкновенной в постпирогенный период
Введение . Одним из основных экологических факторов, влияющих на лесные экосистемы Сибири и существенно корректирующих продуктивность древостоев, следует признать лесные пожары [1]. Устойчивость древостоев к пирогенным повреждениям оценивается по ле-соводственно-таксационным характеристикам (тип насаждения, возраст, диаметр древостоя и др.) [1, 2], тогда как пожары вызывают также существенные физиологические изменения древесных растений [3, 4] .
Одним из основных лесообразующих видов хвойных Сибири является сосна обыкновенная ( Pinus sylvestris L.), которая многими исследователями признана одной из пород, наиболее устойчивых и хорошо адаптирующихся к огневому воздействию. Не раз отмечалось, что сосновые насаждения, которые подвергались воздействию пожара слабой или средней интенсивности, по классификации Курбатского Н.П. [5], в дальнейшем становятся более устойчивыми к огню [6]. В частности, это достигается за счет уменьшения густоты древостоев, увеличения среднего диаметра деревьев, уменьшения запасов горючих материалов, исчезновения ярусности древостоя, что повышает их естественную по-жароустойчивость [7, 8]. Физиологические механизмы, которые могут лежать в основе приобретенной пожароустойчивости, изучались в меньшей степени.
В настоящее время большое внимание уделяется изучению адаптивных реакций и механизмов устойчивости растений в ответ на действие различных стрессовых факторов. Пожар воздействует на различные органы деревьев: ветви, ствол и корни [2-4]. Конвективный поток низового пожара оказывает непосредственное влияние на вегетативные органы растений, что впоследствии сказывается на состоянии организма в целом. Известно, что под влиянием температур, превышающих физиологический оптимум, в растениях происходят многочислен- ные структурные и функциональные изменения, важное место среди которых занимают изменения в фотосинтетическом аппарате [3, 4]. Ряд работ, рассматривающих влияние прогрева на отдельные части растения, указывает на изменения в физиолого-биохимических процессах и показателях, в частности в синтезе стрессовых белков, необходимых для выживания в неблагоприятных условиях [9]. Синтез белков теплового шока (heat shock protein, Hsp) является важной составляющей стрессовой реакции живых систем на воздействия неблагоприятных факторов [10]. Накопление Hsp помогает растениям адаптироваться к неблагоприятным условиям, защищая важнейшие клеточные компоненты [9]. Увеличение содержания Hsp в растениях вызывает интерес у многих исследователей в связи с возможностью использования этого показателя в качестве биохимического маркера защитных реакций растений. Ассимилирующие органы древесных растений являются наиболее чувствительными к изменениям окружающей среды [11]. Исследования, посвященные изучению ответных реакций растений на действие стрессовых факторов, в том числе высоких температур, указывают на проявления определенных защитных механизмов фотосинтезирующих органов, в частности на ускоренный синтез набора Hsp [12]. Однако приведенные данные по накоплению белков-шаперонов отражают результаты кратковременных опытов, тогда как последствия действия температуры наблюдаются в течение нескольких часов. Остается открытым вопрос о длительности перио- да восстановления нормального метаболизма в листьях после стрессового воздействия. Анализ ответных реакций и способности к восстановлению у деревьев после теплового стресса поможет расширить наше понимание механизмов термостабильности. Именно изменение активности фотосинтеза и синтеза стрессовых белков часто является первой реакцией при тепловом стрессе [4, 9, 10, 12].
Цель исследования . Определение различий в накоплении Hsp и в фотосинтетической активности у хвои сосны обыкновенной в постпирогенный период.
Объекты и методы . Объектом исследований служили сосновые молодняки I класса возраста зеленомошно-разнотравной группы типов леса, расположенные на территории экспериментального хозяйства «Погорельский бор» Института леса им. В.Н. Сукачева СО РАН, которая относится к лесостепной зоне Южной Сибири (56°22′ с.ш., 92°57′ в.д.). Исследование проводилось на деревьях экспериментального и контрольного участков размером 0.4 га каждый. В начале вегетационного периода 2014 года на экспериментальном участке (табл.) было проведено контролируемое выжигание, имитирующее низовой пожар средней силы [5], с фиксацией температуры на высоте 4,5 метра. Температура в кроне во время выжигания составила 37 ± 6 ° С, что является ниже предполагаемой для эксперимента температуры. Поэтому образцы для экспериментов отбирались с меньшей высоты, где температура находилась в диапазоне от 40 до 50 ° С.
Таксационные показатели экспериментального участка
|
Диаметр, см |
5,9 |
|
Высота, м |
5,2 |
|
Протяженность кроны, м |
3,3 |
|
Возраст, лет |
21 |
|
Густота, шт/га |
7300 |
На каждой пробной площади (ПП) было выбрано по пять модельных деревьев, произрастающих в одинаковых условиях увлажнения и освещения, сохранивших жизнеспособность, без видимых повреждений насекомыми-ксилофагами, суховершинностей и т.п. Сбор побегов осуществлялся в декабре 2016 г. С ка- ждого дерева с помощью сучкореза с ветвей нижней части кроны отбирали по 3 ветки (средняя высота отбора побегов 2,5–3 м).
После отбора образцы оставляли в комнатных условиях для акклиматизации на 1 сутки.
Для определения устойчивости древесных растений к высоким температурам и их восста- новления в послепожарный период применяли метод искусственного стрессового воздействия, который заключался в прогревании хвои при температурах, превышающих ее физиологический оптимум. Нагрев срезанных побегов осуществляли в термостате, оборудованном внутренним вентилятором для равномерного прогрева образцов, при сублетальной температуре 45 °С. Длительность нагрева составляла 1 час.
Для оценки устойчивости хвои к послепожар-ным условиям регистрировали относительный параметр замедленной флуоресценции (ОПЗФ), отражающий фотосинтетическую активность хвои. Параметры замедленной флуоресценции определяли на флуориметре «Фотон-10» на базе кафедры экологии и природопользования Сибирского федерального университета, учитывая методические рекомендации для отбора хвои сосны обыкновенной и их анализа [13, 14].
После прогрева побеги оставляли в лабораторных условиях при относительной влажности воздуха ≈60±5 % и температуре воздуха ≈24±2 °С на весь период исследования. Затем в течение 4 суток фиксировалось восстановление их фотосинтетической активности. Для каждого измерения использовалась обобщенная проба хвои с исследуемых побегов. Каждые сутки перед измерением хвоя обрезалась с побегов и помещалась в 24 кюветы (по 5 хвоинок в каждой) для измерения интенсивности замедленной флуоресценции. Таким образом определялась термоустойчивость хвои как способность ее клеток активировать механизмы, защищающие фотосинтез, и противостоять тепловому повреждению, что определяет выносливость дерева во время пожара и после него [15].
Материалом для выделения Hsp послужила хвоя второго года жизни контрольных (К) и экспериментальных (горельник, Г) деревьев после суток акклиматизации побегов в лабораторных условиях (К25 или Г25), а также после повторного теплового воздействия на акклиматизированные побеги в течение одного часа при 45 °С (К45 или Г45). Температурная обработка проводилась в темноте в термостате MIR-154 (Panasonic(Sanyo), Япония). Общий белок выделяли по ранее опубликованной методике [16]. Концентрацию белка определяли с помощью реактива Брэдфорда (“Bio-Rad”, США). Перед электрофорезом количество наносимого белка было выравнено, наносили по 10 мкг белка на трек. После разделения белков с помощью Na-ДДС-электрофореза в 12%-м ПААГ и Western Blot в системе mini–Protean III (“BioRad”, США) проводили инкубирование полученных нитроцеллюлозных мембран с нанесенными белками с антителами против стрессовых белков Hsp 70 (Hsp70/Hsc 70; SPA-820, “StressGen Bioreagents”), HSP 60 (SPA-807, “StressGen Bioreagents”, Канада), Hsp 101 (AS07 253, “Agrisera”, Швеция) или Hsp 17,6 (класс I, AS07 254, “Agrisera”, Швеция). Антитела визуализировали с помощью вторичных антител, конъюгированных со щелочной фосфатазой (“Sigma”, США), в присутствии BCIP и NBT (“Gerbu”, Германия). Интенсивность окрашивания белкового пятна на мембране определяли после сканирования в цифровой формат денситометрически с помощью программы Gel Analysis в условных единицах, которые соответствовали количеству окрашенных пикселей на изображении. Содержание Hsp в каждом образце вычисляли относительно контроля, содержание соответствующего белка в котором было принято за единицу.
Эксперименты по полуколичественному определению Hsp проводились минимум в двукратной аналитической повторности для каждой обобщенной пробы с 5 модельных деревьев.
Анализ данных проводили с использованием Microsoft Excel. Определение ОПЗФ проводилось по средней пробе с побегов каждого модельного дерева в трех проворностях. Статистический анализ проводили с использованием программного обеспечения STATISTICA 9. Статистически значимые различия определяли с помощью t-критерия Стьюдента при р ≤0.05.
Результаты и их обсуждение . В природных условиях продолжительность воздействия экстремальной температуры на растения в случае пожара может сильно варьировать и составлять, как правило, от 2 до 15 минут. Действие конвективного потока и возврат температуры к фоновым значениям может по-разному отразиться на физиологическом состоянии растений, что не раз отмечалось в работах по изучению последействий прогревов на динамику теплоустойчивости побегов и хвои, и дерева в целом [1, 2, 4].
Фотосинтез является одним из наиболее чувствительных физиологических процессов, для которого отмечена высокая способность к репарации. Поэтому по изменениям фотосинте- тической активности в период действия и последействия высоких температур можно сделать заключение о функциональной стойкости ассимиляционного аппарата.
В период отбора проб при среднесуточных температурах декабря -20 °С деревья находились в фазе зимнего покоя, о чем свидетельствуют низкие уровни относительного показателя замедленной флуоресценции (ОПЗФ) (рис. 1). При комнатной температуре (25°С) ОПЗФ не показал существенных различий между пробными площадями (К25 и Г25). Прогрев побегов в течение одного часа при 45 °С приводит к подавлению фотосинтетической активности как у деревьев, произрастающих на контрольной пробной площади (К45), так и у деревьев, подвергавшихся действию низового пожара два года назад (Г45). Для побегов, отобранных с пробной площади после низового пожара 2014 года, уровень фотосинтетической активности снижается незначительно – примерно на 6 % относительно исходного. В то же время для побегов, отобранных с контрольной пробной площади и не подвергавшихся огневому воздействию, 1-часовая экспозиция приводит к снижению
ОПЗФ на 35 % относительно исходного уровня. Таким образом, хвоя 2-го года жизни, сформировавшаяся после экспериментального выжигания, отличается более высоким уровнем первичной теплоустойчивости, сохранившимся вплоть до момента исследования.
На следующие сутки экспозиции в лабораторных условиях наблюдается увеличение показателя ОПЗФ как для побегов без искусственного стрессового воздействия, так и после гипертермии. Однако интенсивность восстановления фотосинтетической активности наблюдается в различной степени. У побегов с контрольной пробной площади без действия низового пожара репарация фотосинтеза происходит более интенсивно. Такая реакция возможна ввиду того, что после стрессового воздействия происходит активный синтез термопротекторов, действие которых направлено на поддержание ростовых процессов, в то время как у деревьев с пробной площади после действия низового пожара могла закрепиться кратковременная физиологическая адаптация, которая послужила основой устойчивости в ответ на повторное стрессовое воздействие.
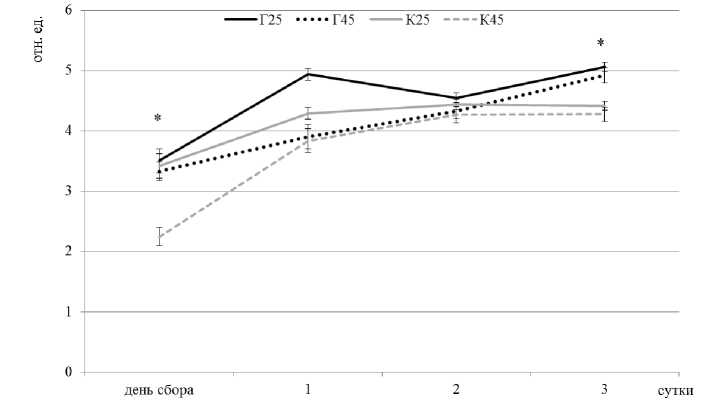
Мера изменчивости представлена стандартным отклонением
* – статистически значимые различия при р≤0.05 (для показателя ОПЗФ между Г45 и К45)
Рис. 1. Изменение интенсивности относительного показателя замедленной флуоресценции (ОПЗФ) хвои сосны обыкновенной после теплового воздействия: К (контроль) - хвоя с деревьев, не подвергавшихся выжиганию; Г (горельник) - хвоя с деревьев после выжигания; К25/Г25 - хвоя без обработки гипертермией; К45/Г45 - хвоя после часового воздействия температурой 45°С. Измерение проводили сразу после тепловой обработки (день сбора) и на 1-, 2- и 3-е сутки после этого
Отечественными и зарубежными специалистами не раз отмечалось, что физиологические функции клеток после снятия действия стрессового фактора могут восстанавливаться [3, 15, 17], в связи с чем в работе регистрировались изменения фотосинтетической активности, происходящие в течение нескольких суток. Наблюдения показали, что на 3-е сутки экспозиции у побегов без искусственного стресса наблюдается рост фотосинтетической активности, что связано с выходом из состояния зимнего покоя в условиях комнатной температуры. ОПЗФ хвои сосны обыкновенной после гипертермии также увеличивается на 3-и экспериментальные сутки, при этом уровень фотосинтетической активности близок к показателям, которые характерны для необработанных высокими температурами образцов. Более высокие значения ОПЗФ зарегистрированы для побегов деревьев с пробной площади после действия низового пожара. Таким образом, низовой пожар 2014 г. привел к повышению первичной термоустойчивости и способности деревьев к репарации, которая проявляется после повторного воздействия повреждающей температурой.
Исследование содержания Hsp проводилось на хвое побегов, отобранных в холодное время года и, судя по показателям ОПЗФ хвои (см. рис. 1), находящихся в фазе зимнего покоя.
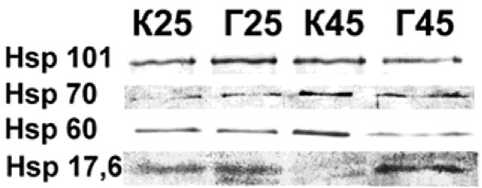
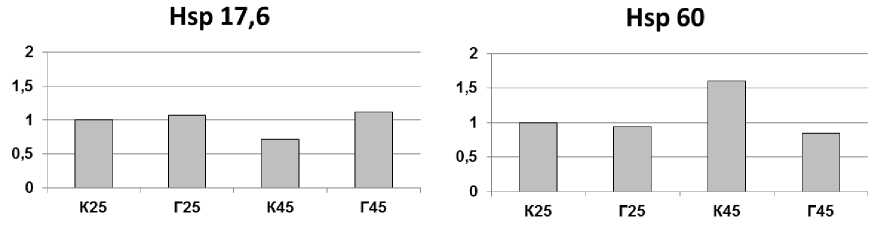
После суток акклиматизации в лабораторных условиях было проведено полуколичественное определение содержания высокомолекулярных белков (Hsp 101, 70 и 60) и низкомолекулярного (Hsp 17,6) для хвои с контрольной пробной площади (К) и хвои с деревьев после контролируемого выжигания (горельник, Г). Термическая обработка вызывала закономерное накопление Hsp 101, Hsp 70 и Hsp 60 в хвое контрольных деревьев, но приводила к снижению содержания Hsp 17,6 в ней, хотя низкомолекулярные Hsp, как правило, активно накапливаются при гипертермии [18]. Сведений об Hsp голосеменных древесных растений крайне мало. Отмечено, что Hsp хвои сосны обыкновенной условно можно разделить на группы: накапливающиеся преимущественно в холодное время года (Hsp 70), в теплое время года (Hsp 17,6), и присутствующие в клетках хвои постоянно, но с небольшими количественными изменениями (Hsp 101, Hsp 60) [16]. Возможно, причиной снижения со- держания Hsp 17,6 под действием гипертермии стало использование в эксперименте зимней хвои, у которой экспрессия гена Hsp 17,6, по-видимому, полностью подавлена в холодное время года [16], и даже предварительная акк-лимация побегов при комнатной температуре, судя по нашим данным, не смогла полностью активировать его тепловую регуляцию.
У хвои с деревьев после выжигания термическая обработка не вызвала накопления Hsp 60 и Hsp 17,6, содержание Hsp 101 снизилось. Учитывая, что в пробах Г25 содержание Hsp оказалось выше, чем в К25 (особенно это касается Hsp 101, для Hsp 17,6 в пробах Г25 была выявлена дополнительная полоса белка более низкой массы, но тоже реагирующая со специфическими антителами), можно предположить, что произведенное в прошлом выжигание уже активировало экспрессию генов этих Hsp, так что дополнительное воздействие не вызвало дополнительной активации. Накопление Hsp70 после тепловой обработки в хвое после выжигания оказалось таким же, как и у контрольных деревьев.
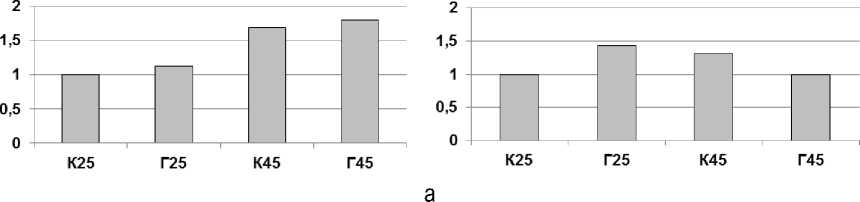
Согласно Даунс и соавт. [19], листья, которые сформировались при действии высоких температур, могут быть устойчивы к тепловому стрессу благодаря наличию Hsp, но как долго сохраняется данный эффект, остается неясным. Результаты, представленные на рисунке 2, свидетельствуют, что содержание белков Hsp 101 и Hsp 70 при экспозиции в лабораторных условиях выше у хвои с послепожарной пробной площади. Накопление Hsp, как правило, сопровождается ростом устойчивости к данному стрессору, возможно, частично благодаря этому обработка побегов температурой в 45 °С не привела к значительному снижению ОПЗФ (см. рис. 1). Стрессирование в лабораторных условиях привело к увеличению содержания всех исследуемых высокомолекулярных белков с обеих пробных площадей. При этом содержание Hsp 101 и Hsp 60 оказалось выше для образцов с контрольной пробной площади, что, возможно, повлияло на более высокие темпы восстановления фотосинтетической активности на следующие сутки после прогрева, о чем говорит показатель ОПЗФ.
Таким образом, полученные нами данные согласуются с работами как отечественных, так и зарубежных исследователей. Так, Титов с со-авт. [17] отмечают, что повышение устойчивости растений может быть вызвано не только длительными (часы, сутки), но и кратковременными (секунды, минуты) воздействиями неблагоприятных температур. В случае контролируемого выжигания кратковременное действие высоких температур на вегетативные органы может также являться экстремальным стрессовым воз- действием. Рост содержания специфических Hsp в хвое с послепожарных деревьев может говорить об их участии в повышении термоустойчивости клеток даже в течение нескольких лет после пожара, что выражается в более высоких темпах репарации фотосинтетической активности хвои после повторного стрессового воздействия.
Hsp 70 Hsp 101
б
Рис. 2. Изменение содержания Hsp в хвое сосны обыкновенной из подвергнутых искусственному выжиганию (Г) и контрольных сосняков (К) после часового теплового воздействия при 45 °С (К45 и Г45) и без него (К25 и Г25). Представлено изображение типичной мембраны после визуализации антител (а) и результат денситометрической оценки интенсивности окрашивания белковых пятен (б). Интенсивность окрашивания каждого белка представлена в условных единицах относительно контроля (К25), принятого за 1
Заключение. Таким образом, кратковременное воздействие повышенных температур при контролируемом выжигании, имитирующем действие низового пожара средней интенсивности, может вызывать повышение общей устойчивости растений. Проведенные экспериментальные исследования выявили повышение устойчивости клеток хвои, которая не подверга- лась непосредственному воздействию неблагоприятной температуры, а сформировалась в период восстановления после пожара. Это может свидетельствовать о том, что на уровне организма, испытавшего воздействие стрессора, сохраняется сигнал, способный вызывать адаптивные (физиологические и биохимические) перестройки, в конечном итоге приводящий к по- вышению резистентности ассимиляционного аппарата. В частности, для сосновых деревьев накопление стрессовых белков в хвое может быть одним из компонентов защитной реакции, определяющих возможность восстановления нормальной жизнеспособности и сохранения устойчивости к повторному действию повышенных температур на протяжении нескольких лет восстановления после пожара.
Список литературы Белки теплового шока и фотосинтетическая активность хвои сосны обыкновенной в постпирогенный период
- Валендик Э.Н., Косов И.В. Влияние теплового излучения лесного пожара на окружающую среду//Сиб. экол. журн. -2008. -№ 4. -С. 517-523.
- Иванова Г.А., Конард С.Г., Макрае Д.Д. Воздействие пожаров на компоненты экосистемы среднетаежных сосняков Сибири. -Новосибирск: Наука, 2014. -232 с.
- Wang R., Hu H.-Q. Physiological response of Betula platyphylla leaves to fire and the restoration after fire//Journal of Beijing Forestry University. -2014. -V. 36(1). -Р. 31-34.
- Гирс Г.И. Физиология ослабленного дерева. -Новосибирск: Наука, 1982. -255 с.
- Курбатский Н.П. Техника и тактика тушения лесных пожаров. -М.: Гослесбумиздадт, 1962. -154 с.
- Верховец С.В. Влияние контролируемых выжиганий на пожароопасность и лесовосстановление на сплошных вырубках: дис.... канд. с.-х. наук: 06.03.03. -Красноярск, 2000. -200 с.
- Фуряев В.В., Фуряев Е.А. Пироэкологические свойства сосны обыкновенной в Средней Сибири//Хвойные бореальной зоны. -2008. -№ 1-2. -С. 103-108.
- Цветков П.А. Пирогенные свойства древесных пород//Лесоведение. -2011. -№ 2. -С. 25-31.
- Wahid A., Gelani S., Ashraf M. . Heat tolerance in plants: An overview//Environmental and Experimental Botany. -2007. -V. 61 (3). -P. 199-223
- Timperio M., Egidi G., Zolla L. Proteomics applied on plant abiotic stresses: role of heat shock proteins (HSP)//Journal Proteomics. -2008. -V. 71(4). -P. 391-411.
- Тужилкина В.В. Реакция пигментной системы хвойных на длительное аэротехногенное загрязнение//Экология. -2009. -№ 4. -С. 243-248.
- Кулаева О.Н. Белки теплового шока и устойчивость растений к стрессу//Соросовский образовательный журнал. -1997. -№ 2. -С. 5-13.
- Григорьев Ю.С., Андреев Д.Н. К вопросу о методике регистрации замедленной флуоресценции хлорофилла при биоиндикации загрязнения воздушной среды на хвойных//Естественные науки. -2012. -№ 2. -С. 36-39.
- Патент № 2069851. Способ определения содержания фитотоксических веществ/Григорьев Ю.С., Фуряев Е.А., Андреев А.А.//Бюл. изобретений от 27.11.1996. -№ 33.
- Ланге О.Л. Исследование изменений теплоустойчивости у растений. Клетка и температура среды. -М.; Л.: Наука, 1964. -С. 91-97.
- Korotaeva N.E., Oskorbina M.V., Kopytova L.D. . Variations in the content of stress proteins in the needles of common pine (Pinus sylvestris L.) within an annual cycle//Journal of Forest Research. -2012. -Т. 17. -№ 1. -С. 89-97.
- Титов А.Ф., Таланова В.В. Устойчивость растений и фитогормоны. -Петрозаводск: Изд-во Карельского НЦ РАН, 2009. -206 с.
- Xu Y., Zhan C., Huang B. Heat Shock Proteins in Association with Heat Tolerance in Grasses//International Journal of Proteomics. -2011.
- Downs C.A., Coleman J.S., Heckathorn S.A. The chloroplast 22-kDa heat shock protein: a lumenal protein that associates with the oxy-gen evolving complex and protects photosystem II during heat stress//J Plant Physiol. -1999. -P. 155:477-487.
