Бентофауна и ихтиофауна переката средней равнинной реки, населяемого русской быстрянкой Alburnoides rossicus Berg

Автор: Котельникова В.С.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 3, 2017 года.
Бесплатный доступ
Изучен видовой состав рыб переката р. Валы, где русская быстрянка занимает лидирующие позиции в ихтиофауне по биомассе и доминирует по численности, превосходя все прочие компоненты ихтиоценоза. В бентофауне реки в районе исследований зарегистрировано 46 видов и форм, принадлежащих 4 классам: малощетинковые черви (4 вида), двустворчатые моллюски (5), брюхоногие моллюски (1) и насекомые (36). Наибольшая доля в биомассе зообентоса в период исследований принадлежала двустворчатым моллюскам (48.2%), малощетинковым червям (21.7%) и ручейникам (8.4%о), по численности преобладали хирономиды (54.4%) и ручейники (11.6%>). Содержимое кишечников русской быстрянки состоит из 33 компонентов, из которых около 9% - водоросли и высшие растения, 88%> - беспозвоночные животные, 3%> - неорганические компоненты. Среди насекомых отмечены бентосные и наземно-воздушные формы. По характеру питания русская быстрянка может быть отнесена к типичным полифагам.
Питание рыб, зообентос рек, ихтиофауна рек
Короткий адрес: https://sciup.org/147204837
IDR: 147204837 | УДК: 574.584+587
Benthofauna and ichthyofauna of the riffle of the medium lowland river inhabited by Albburnoides rossicus Berg
The species composition of fish of the riffle of the Vala River was studied, in which a Russian spirlin was present. It occupies a leading position in the ichthyofauna for biomass and dominates in abundance, exceeding all other components. In benthic community we had found 46 taxa of 4 classes: Oligochaeta (4 species), Bivalvia (5), Gastropoda (1), Insecta (36). The largest share in zoobenthos biomass during the study period belonged to bivalved mollusks (48.2%), oligochaetes (21.7%) and caddis flies (8.4%); chi-ronomids (54.4%) and caddis flies (11.6%) predominated in abundance. The food spectrum of Russian spirlin Alburnoides rossicus from Vala River in the Udmurt Republic was studied. It consists of 33 components, among them 9% are algae and higher plants, 88%> - invertebrates, 3%> - mineral components. One types of insects were found: benthic forms, and land-air ones. According to food spectrum Russian spirlin may be considered as a typical polyphage.
Текст научной статьи Бентофауна и ихтиофауна переката средней равнинной реки, населяемого русской быстрянкой Alburnoides rossicus Berg
Род Alburnoides Jeitteles, 1861 со значительным количеством видов относится к широко распространенным рыбам Европы [Ручин, 2013]. Быстрянка выступает важнейшим компонентом экосистем текучих вод, образуя большие скопления в реках, в которых обитает, превосходя все прочие компоненты ихтиоценозов. Таким образом, представители этого рода являются важным элементом трофической цепи рек. Кроме того, эти виды успешно используются как ин- дикаторы чистоты водотоков [Raikova-Petrova et al., 2011].
Русская быстрянка Alburnoides rossicus Berg, 1924 выделена как самостоятельный вид не так давно, и данные по её морфологии, экологии и биологии в литературе довольны ограничены. Изученная нами ранее русская быстрянка A. rossicus из верховьев р. Чепцы (территория Удмуртской Республики) по характеру питания относится к по-ли(эври)фагам. В условиях отсутствия зоопланктона или его низкой концентрации, молодь быстрян-
ки использует в пищу дрифтующих донных беспозвоночных животных. При достижении половозре-лости быстрянка в большей степени питается зоо-и, особенно, фитобентосом (фитоперифитоном). [Котельникова, 2016]. Также в литературе присутствуют данные по питанию наиболее близкого вида – обыкновенной быстрянки Alburnoides biрunctatus . Считается, что обыкновенная быст-рянка питается растительной и животной пищей: личинками насекомых, а также наземными формами, которые попадают на поверхность воды. Кроме насекомых, обыкновенная быстрянка употребляет мелких ракообразных, червей и диатомовые водоросли [Lelek, 1987; Treer et al., 2006].
Цель данной работы – изучение особенностей биоценоза переката средней равнинной реки, населяемого русской быстрянки A. rossicus . Для осуществления этой цели поставлены следующие задачи: установить видовой состав ихтиофауны и бенто-фауны исследуемого переката, определить таксономический состав пищи русской быстрянки и характер её питания; выявить закономерности в питании русской быстрянки.
Материал и методы исследования
Река Вала – приток 2-го порядка р. Вятки, берёт начало на Можгинской возвышенности и протекает по территории Удмуртской Республики и Кировской обл. Длина реки – 196 км, площадь бассейна – 7 360 км² [Ресурсы…, 1967]. По совокупности признаков (длина, площадь водосбора) данный водоток относится к средним рекам [Комлев, Черных, 1984; Нежиховский, 1988].
Материал собирали в р. Вале 16 августа 2015 г. в утренние часы местного времени (8–10 ч. утра). Исследованный участок (N 56.453008º E 52.369820º) расположен в верхнем течении р. Валы на 152-м км от устья (рисунок) в Можгинском р-не Удмуртской Республики и представляет собой перекат с гравийно-песчаным дном и большим количеством высшей водной растительности, преимущественно рдестами. Скорость течения на стрежне переката достигала 0.25 м/с. Русло реки глубоко врезано, берега обрывистые. Средняя глубина составляла 0.6 м, максимальная – 1.5 м.
Орудием лова рыбы был электролов ЭЛЛОР-2. Площадь обловленного участка составила 150 м². Для анализа спектра питания русской быстрянки A. rossicus использованы пробы рыб и зообентоса. Рыб для изучения питания фиксировали 4%-ным раствором формалина [Боруцкий, 1974]. Расчеты и измерения морфометрических признаков с точностью до 0.1 мм при помощи штангенциркуля проводились в соответствии с работой К. Хуббса и К. Лаглера [Hubbs, Lagler, 1958]. Длина тела измерялась до конца прободенных чешуй. Полную массу тела рыб устанавливали с помощью весов WT-300
с точностью до 0.01 г. Пол и стадию зрелости гонад определяли по И.Ф. Правдину [Правдин, 1966]. Для определения возраста с левого бока рыбы отбирали чешую. Всего исследовано содержимое пищеварительных трактов 51 экземпляра русской быстрянки, 11 из исследованных пищеварительных трактов оказались пустыми.
Параллельно сбору рыбы, непосредственно в местах обитания быстрянки, были отобраны пробы зообентоса (6), которые собирали при помощи бен-тометра с площадью захвата 625 см². При промывании проб бентоса использовали газ с размером ячеи 220 мкм [Методика …, 1975].
Для характеристики структуры донных сообществ рассчитывали индекс Шеннона и его стандартную ошибку [Песенко, 1982, Алимов, 2001].
Для выделения доминирующих видов в биоценозах применяли индекс плотности Л.В. Арнольди [Арнольди, 1949] в модификации Г.Х. Щербины [Щербина, 1993]:
ID = XiN X Px В.
где P – встречаемость (%), N – численность вида (%), В – биомасса вида (%).
Для восстановления массы съеденных организмов использованы средние массы тел, полученные в результате взвешивания пищевых компонентов из гидробиологических проб, взятых одновременно с пробами на питание рыб [Боруцкий, 1974]. Массу тела личинок хирономид в питании рыб для видов, отсутствующих в гидробиологических пробах, восстанавливали по ширине головной капсулы по уравнению Е.В. Балушкиной [Балушкина, 1987]:
IV' = 34.68 xdp.
где W' – средняя масса личинки данного возраста, мг, d k – ширина головной капсулы личинки, мм.
Масса компонентов содержимого пищеварительных трактов, имеющих малое значение, либо трудно выделяемых (фрагменты водорослей, высших растений, песок) определялась на глаз в долях от общей фактической массы пищевого комка, либо, по возможности, путём прямого взвешивания [Боруцкий, 1974].
Общий индекс наполнения (ОИН, 0/ 000 ) рассчитывался по формуле
ОИН = -^X 10000
где q ф.п.к – фактическая масса пищевого комка, мг, Q – масса тела рыбы, мг.
Общий индекс потребления (ОИП, 0/ 000 ) рассчитывался по формуле
ОИП =-^x 10000.
где q в.п.к – восстановленная масса пищевого комка, мг, Q – масса тела рыбы, мг [Боруцкий, 1974].
Количественную оценку избирательности питания рыб проводили по формуле В.С. Ивлева [1955]:
— й С — ’
Л "^ A
где xi – доля i-корма у вида х, уi – доля i-корма у вида у. Индекс равен нулю при полном различии пищевых где ri – относительное содержание компонента в пищевом комке (в виде процента от всей массы пищевого комка), pi – относительное содержание того же компонента во внешней среде.
Большее или меньшее совпадение пищевых спектров у рыб разных сравниваемых групп (пол, возраст) свидетельствует о степени сходства состава пищи. Степень перекрывания пищевых ниш возрастных и половых групп рассчитывали по ин- ниш и равен единице при полном их совпадении. Значение индекса >0.6, расценивалось как биологически значимое перекрывание пищевых ниш [Wallace, 1981].
При оценке встречаемости компонентов в питании рыб, за 100% принималось общее количество пищеварительных трактов, в том числе и пустых [Боруцкий, 1974]. Нами была принята сле- дующая классификация встречаемости компонен-
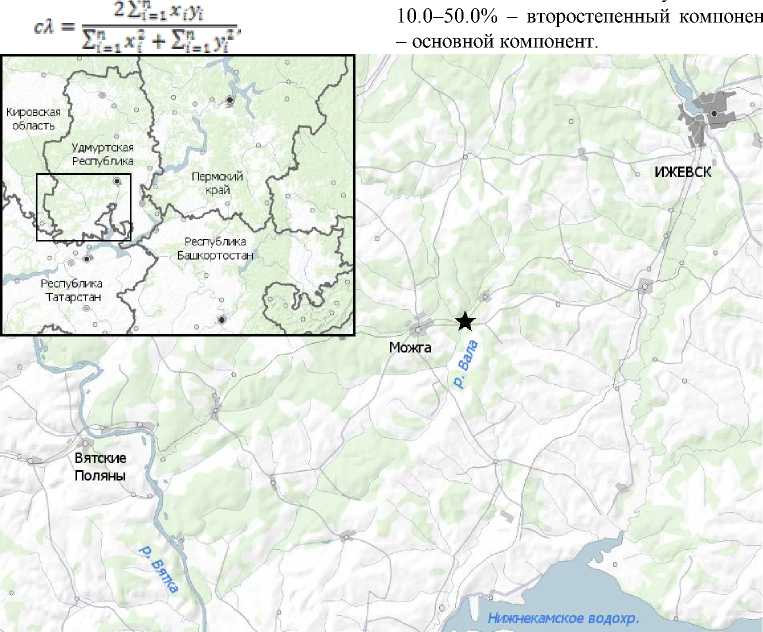
10.0–50.0% второстепенный
– основной компонент.
–
ИЖЕ1
Можга гое водохр.
Республика Татарстан
Удмуртская Республика
Республика Башкортостан..
Кировская область
Пермский край
Вятские Поляны
Карта-схема района исследования.
Звездочкой указано место проведения работ
Для оценки статистической значимости различий половых и возрастных групп рыб использовали однофакторный дисперсионный анализ. Апостериорные сравнения средних значений в ходе дисперсионного анализа проводили с использованием LSD-теста Снедекора-Фишера. Статистически значимыми считали различия при p <0.05. Индекс Мориситы-Хорна рассчитывали в статистической среде R с использованием функции similary [Шитиков, Розенберг, 2013]. Стандартную ошибку индекса находили методом бутстрепа на основе 1000 итераций.
Результаты
Структура ихтиоценоза
На исследованном перекате р. Валы отловлено
29 особей 6 видов рыб, относящихся к 3 семействам: Cyprinidae (4 вида), Cobitidae (1 вид), Esoci-dae (1 вид).
Относительная численность рыб составила 5 403 экз./га, относительная масса – 61.36 кг/га (табл. 1). В структуре рыбного сообщества по численности преобладала русская быстрянка Albur-noides rossicus (83%), по массе – голавль Squalius cephalus (50%), также большую роль по массе играла русская быстрянка (36%), доля остальных видов была значительно ниже. Таким образом, A. rossicus формирует основу данного ихтиоценоза, превосходя по численности все прочие компоненты.
Все виды рыб, обитающие на данном участке р. Валы, по классификации Никольского [Никольский, 1980] относятся к 2 пресноводным фаунистическим комплексам: понто-каспийский ком- плекс представлен голавлем, уклеей и быстрянкой, бореальный равнинный комплекс – плотвой, щукой, щиповкой.
Таблица 1
Относительные численность (экз./га) и масса (кг/га) рыб исследованного переката р. Валы в
2015 г.
|
Вид |
экз./га |
кг/га |
|
Squalius cephalus (Linnaeus, 1758) |
133 |
30.68 |
|
Rutilus rutilus (Linnaeus, 1758) |
400 |
5.34 |
|
Esox lucius Linnaeus, 1758 |
67 |
1.33 |
|
Cobitis taenia Linnaeus, 1758 |
67 |
0.67 |
|
Alburnoides rossicus Berg, 1924 |
4469 |
22.01 |
|
Alburnus alburnus (Linnaeus, 1758) |
267 |
1.33 |
|
Всего |
5403 |
61.36 |
По отношению к течению все рыбы относятся к реофильным и рео-лимнофильным видам, что характерно для верховий рек [Никольский, 1974].
Практически все виды рыб отличаются по характеру питания, формируя 5 групп: хищники (щука), бентофаги (щиповка), планктофаги (уклея), полизоофаги (голавль), поли(эври)фаги (плотва и быстрянка). При этом по составу пищи большинство видов преимущественно зоофаги, кроме плотвы и быстрянки, которые способны питаться не только животными, но также растительной пищей и детритом [Никольский, 1974; Kottelat, Freyhof, 2007].
Все отмеченные виды рыб нерестятся преимущественно поздней весной, за исключением щуки, которая начинает размножаться при температуре воды 5ºС. Для откладки икры все виды используют определённый субстрат. Фитофилами являются плотва, щука и уклея, литореофилы – голавль и быстрянка, щиповка предпочитает песчаный субстрат (псаммореофил). [Никольский, 1974; Kotte-lat, Freyhof, 2007].
Структура донных сообществ
В бентофауне реки в районе исследований зарегистрировано 46 видов и форм, принадлежащих 4 классам: малощетинковые черви (4 вида), двустворчатые моллюски (5), брюхоногие моллюски (1) и насекомые (36). Среди последних отмечены подёнки (5 видов), ручейники и жуки (по 2 вида), стрекозы, клопы, большекрылые, атерициды, ко-мары-болотницы, слепни (по 1 виду), мокрецы (не идентифицированы) и хирономиды (20 видов и форм).
Биомасса зообентоса реки на исследованном перекате в среднем составила 32.5±20.9 г/м² при численности 7.8±2.9 тыс.экз./м². Наибольшая доля в биомассе зообентоса в период исследований принадлежала двустворчатым моллюскам (48.2%), малощетинковым червям (21.7%) и ручейникам (8.4%), по численности преобладали хирономиды (54.4%) и ручейники (11.6%) (табл. 2).
К постоянным видам-зообентонтам, частота встречаемости которых превысила 50%, относятся 3, из них олигохета Lumbriculus variegatus (Muller, 1773) и хирономида Stictochironomus stic-ticus (Fabricius, 1781), отмечены в 60–80% проб. Максимальной частотой встречаемости (более 80%) характеризовались хирономида Cladotanytar-sus mancus (Walker, 1856).
Таблица 2
Численность (экз./м²) и биомасса (г/м²) основных групп зообентоса р. Валы в 2015 г.
|
Группа |
экз./м² |
г/м² |
|
Oligochaeta |
801 |
7.03 |
|
Bivalvia |
583 |
15.61 |
|
Gastropoda |
17 |
0.08 |
|
Ephemeroptera |
567 |
0.88 |
|
Odonata |
33 |
0.05 |
|
Trichoptera |
900 |
2.73 |
|
Coleoptera |
33 |
0.01 |
|
Heteroptera |
133 |
1.05 |
|
Megaloptera |
17 |
0.25 |
|
Athericidae |
67 |
1.67 |
|
Ceratopogonidae |
217 |
0.03 |
|
Limoniidae |
133 |
0.52 |
|
Tabanidae |
50 |
0.50 |
|
Chironomidae |
4236 |
1.96 |
|
Всего |
7787 |
32.37 |
Монодоминантом в донных сообществах выступала олигохета L. variegatus , субдоминантом – двустворчатый моллюск Pisidium amnicum (O.F. Muller, 1774). Величина индекса Шеннона, рассчитанного по численности донных сообществ, составила 4.34±0.02 бит/экз., по биомассе – 3.12±0.04 бит/г.
Общая характеристика питания русской быстрянки
При средних длине и массе рыб – 6.9±0.2 см и 6.0±0.4 г, соответственно, восстановленная масса пищевого комка составила 56.9±14.3 мг, ОИН – 76.3±20.50/ 000 , ОИП равнялся 99.8±24.90/ 000 .
Содержимое пищевого тракта русской быст-рянки состояло из 33 компонентов, из которых 3 объекта приходилось на растительные формы, относящиеся к 2 отделам: Chlorophyta (зеленые водоросли), Magnoliophyta (покрытосеменные). Зеленые водоросли представлены 2 видами, из них наибольшие встречаемость (в 8% проб) и долю в восстановленной массе пищевого комка имела зеленая нитчатая водоросль Ulothrix tenuissima Kütz.
Среди животных компонентов пищеварительного тракта русской быстрянки было отмечено 29 объектов, относящихся к 2 классам беспозвоночных: Arachnida (паукообразные) и Insecta (насекомые). Наибольшим разнообразием отличались насекомые, среди которых отмечено 28 таксонов из 5 отрядов. Из них к разным стадиям развития амфи-биотических насекомых относилось 27 форм, к на- земно-воздушным – 1 форма. Кроме того, в содержимом пищеварительных трактов отмечен песок. Зоопланктон в пищевых комках рыб не зарегистрирован.
Видовое богатство поденок (Ephemeroptera) было относительно велико, в бентофауне отмечено 5 видов, из которых в пищеварительных трактах русской быстрянки зарегистрирован только 1 вид – Baetis vernus Curtis, 1834, который играл существенную роль в питании, встречаясь в 54.9% проб.
Среди ручейников (Trichoptera) в зообентосе изученной реки зарегистрировано 2 вида, тогда как в пищевых комках отмечено 3 таксона видового ранга, из которых 1 вид, Hydropsyche contubernalis McLachlan, 1865, входил в доминантные комплексы донных сообществ, но не имел большого значения в питании русской быстрянки.
Жуки (Coleoptera), представленные семейством Dytiscidae, не были зарегистрированы в зообентосе данного водотока, но выступали второстепенным компонентом питания, встречаясь в 13.7% проб.
Клопы (Heteroptera) не играли существенной роли в численности и биомассе зообентоса. Их наличие в питании быстрянки носило случайный характер (2% встречаемости).
Двукрылые (Diptera) были наиболее богато представлены как в бентофауне, так и в питании русской быстрянки. В пищеварительных трактах обнаружено 22 таксона видового ранга из 5 семейств двукрылых, что составляет около 67% от общего таксономического списка беспозвоночных животных. Наибольшую частоту встречаемости в питании проявил вид мошек (Simuliidae) – Simu-lium ornatum Мeigen, 1818 (в 70.6% проб), не отмеченный в бентосе исследованного переката. Представители комаров-болотниц (Limoniidae), атерицид (Athericidae), комаров-долгоножек (Tipulidae) встречались в питании быстрянки редко (2.0–3.9% проб). Около 52% от всего видового богатства двукрылых насекомых, отмеченных в питании, составили комары-звонцы – 17 таксонов. Наибольшую частоту встречаемости в питании бы-стрянки обнаружили Cricotopus bicinctus (Meigen, 1818), Orthocladius rhyacobius Kieffer, 1911 и Thienemanniella vittata (Edwards, 1924) (13.7%), также отсутствующие в донных сообществах переката.
Наземно-воздушные формы в питании быст-рянки были представлены пауками-кругопрядами (Araneidae), которые не играли существенной роли по встречаемости и, по всей видимости, являлись случайным компонентом в пище.
Ведущую роль по массе в пищевом комке бы-стрянки играли личинки и имаго мошек S. ornatum , составляя около 84% восстановленной массы пищевого комка. Отсутствие этого вида в бентосе свидетельствует о том, что данный объект потребляется русской быстрянкой из толщи воды. Это объясняется тем, что развитие преимагиналь-ных фаз мошек связано с регулярными (судя по всему обязательными) массовыми миграциями личинок, плывущих вниз по течению; в это время они служат одним из основных компонентов пищи рыб [Определитель …, 1999].
Максимальные величины индекса избирательности Ивлева ( Е ) среди бентосных животных имели поденка B. vernus (0.7) и представители комаров-звонцов Polypedilum spp. (0.4).
Особенности питания русской быстрянки в зависимости от возраста
В целом, с возрастом спектр питания русской быстрянки значительно расширяется, от 7 до 19 компонентов. Наибольшим разнообразием спектра питания отличались пятилетки (19 компонентов) и семилетки (18 компонентов) за счёт видового богатства хирономид, при этом у них отмечены наименьшие величины восстановленной массы пищевого комка, ОИН и ОИП. У особей возраста 2+ обнаружено наименьшее количество компонентов питания (табл. 3).
В среднем, восстановленная масса пищевого комка, интенсивность питания и накормленность рыб снижались от трёхлеток до пятилеток, а затем возрастали у шести- и семилеток. При этом, величины ОИН и ОИП варьировали в широких пределах: от 50.7 до 130.80/ 000 , и от 55.4 до 164.20/ 000 , соответственно. Величины ОИН и ОИП трёхлеток были значимо больше, чем у пятилеток и семилеток. Восстановленная масса пищевого комка только у пятилеток была значимо ниже, чем у шестилеток. Остальные возрастные группы не отличались по этому показателю (табл. 3).
Таблица 3
Основные характеристики питания разных возрастов русской быстрянки ( M±tm ) в р. Вале в 2015 г.
|
Показатель |
Возраст |
||||
|
2+ |
3+ |
4+ |
5+ |
6+ |
|
|
Количество исследованных рыб, шт. |
7 |
9 |
9 |
13 |
13 |
|
Длина рыб, см |
5.9±0.2 |
6.4±0.2 |
6.8±0.2 |
7.1±0.1 |
7.5±0.1 |
|
Масса рыб, г |
3.6±0.4 |
4.7±0.2 |
5.6±0.4 |
6.5±0.3 |
7.8±0.3 |
|
Общий индекс потребления, 0/ 000 |
164.2±76.6 |
108.2±81.5 |
55.4±40.1 |
121.4±57.2 |
69.3±41.5 |
|
Общий индекс наполнения, 0/ 000 |
130.8±77.1 |
96.2±72.4 |
41.6±36.7 |
88.4±40.6 |
50.7±29.7 |
|
Восстановленная масса пищевого комка, мг |
61.4±27.4 |
49.5±37.3 |
29.9±19.9 |
80.7±37.4 |
54.5±33.5 |
|
Количество компонентов |
7 |
11 |
19 |
11 |
18 |
Окончание табл. 3
|
Показатель |
Возраст |
||||
|
2+ |
3+ |
4+ |
5+ |
6+ |
|
|
Основные компоненты |
S. ornatum (larvae) B. vernus (larvae) S. ornatum (imagoes) |
S rnatum (larvae) B. vernus (larvae) S. ornatum (imagoes) |
S. ornatum (larvae) B. vernus (larvae) Araneidae (imagoes) |
S. ornatum (larvae) A. ibis (larvae) B. vernus (larvae) |
S. ornatum (larvae) B. vernus (larvae) P. latipennis (pupae) |
Примечание. ± – предельная ошибка среднего значения.
Степень перекрывания пищевых ниш всех возрастных групп оказалась очень высока: величины индекса cλ варьировали от 0.92 до 0.99.
Основу содержимого пищеварительных трактов всех отмеченных возрастных групп (2+-6+) по массе составляли личинки мошек S. ornatum , доля которых была значимо выше только у шестилеток по сравнению с пятилетками. Кроме того, в питании рыб всех возрастов велико значение животных, отмеченных в бентосе данного переката, а именно – подёнок B. vernus , атерицид Atherix ibis (Fabricius, 1798), куколки ручейников Potamophy-lax latipennis (Curtis, 1834), доля которых в питании рыб разного возраста значимо не различалась.
Особенности питания русской быстрянки в зависимости от пола ветственно. Пищевой спектр самок расширялся благодаря большей представленности ручейников, растительных компонентов и нехирономидных двукрылых. При этом по величине восстановленной массы пищевого комка, индексам ОИП и ОИН значимых различий не зарегистрировано (табл. 4). Степень перекрывания пищевых ниш (cλ) самцов и самок весьма высока и составляла 0.99±0.23.
Основу пищи по массе, как самцов, так и самок, играли личинки мошек S. ornatum и подёнки B. vernus , доля которых в пищевом комке не зависела от половой принадлежности. Различия заключались лишь в представленности дополнительных объектов питания. Так, у самок это представители ручейников: P. latipennis и H. сontubernalis , у самцов – представители жуков семейства Dytiscidae и зелёная нитчатая водоросль U. tenuissima (табл. 4).
В спектрах питания самцов и самок обнаруже но разное количество компонентов: 18 и 25, соот-
Таблица 4
Основные характеристики питания самок и самцов русской быстрянки ( M±tm ) в р. Вале в 2015 г.
|
Показатель |
Пол |
|
|
Самцы |
Самки |
|
|
Количество исследованных рыб, шт. |
23 |
28 |
|
Длина рыб, см |
6.6±0.2 |
7.1±0.2 |
|
Масса рыб, г |
5.4±0.6 |
6.5±0.6 |
|
Общий индекс потребления, 0/ 000 |
93.1±37.8 |
105.4±35.2 |
|
Общий индекс наполнения, 0/ 000 |
69.9±29.4 |
81.6±30.0 |
|
Восстановленная масса пищевого комка, мг |
47.8±19.4 |
64.3±21.4 |
|
Количество компонентов |
18 |
25 |
|
Основные компоненты |
S. ornatum (larvae) B. vernus (larvae) Dytiscidae (imagoes) |
S. ornatum (larvae) B. vernus (larvae) P. latipennis (pupae) |
Примечание. ± – предельная ошибка среднего значения.
Обсуждение
Основу рыбного сообщества исследованного участка р. Валы слагают реофильные виды понто-каспийского пресноводного комплекса (русская быстрянка и голавль), которые приспособлены к жизни в реках с каменистым дном, прозрачной водой, насыщенной кислородом. Здесь к представителям понтического комплекса присоединяются реолимнофильные рыбы бореального равнинного комплекса (плотва).
Фауна донных сообществ исследованного уча- стка р. Валы включает в себя эвритермные и тепловодные организмы, среди которых доминируют формы, достигающие своего максимального развития в стоячих водах, например моллюски и олигохеты. По классификации Хайнса, данный участок реки может быть отнесён к потамали [Hynes, 1970].
Также большое значение в зообентосе имели пассивные фильтраторы (Hydropsychidae), альго-фаги-соскребатели обрастаний (Heptageniidae, Baetidae), щелевые детритофаги (Chironomidae), жизнь которых в основном связана с более или менее быстрым течением, и все жизненные формы отличаются наличием приспособлений к течению, из которых основное – способность надёжно прикрепляться к стабильному субстрату, что характеризует население ритрали [Illies, 1961; Чертопруд, 2014].
Таким образом, по гидробиологическим характеристикам исследованный участок реки можно отнести к переходной зоне от ритрали к потамали, а с учётом гидрологических параметров – к эпипо-тамали.
В неоднородной по своему происхождению фауне наиболее напряжённые пищевые отношения складываются между видами, принадлежащими к разным фаунистическим комплексам, но потребляющими сходные экологические группы кормов. Особенностью ихтиоценоза исследованного участка р. Валы является расхождение видов рыб по типу питания. Таким образом, ослабляется межвидовая конкуренция внутри сообщества.
Разнообразие и видовое богатство содержимого пищеварительных трактов русской быстрянки р. Валы свидетельствует о высокой степени полифагии, но по составу пищи A. rossicus данного водотока преимущественно зоофаг.
В целом тип питания русской быстрянки р. Валы можно определить как «бенто-пелагический». Большая часть животных объектов питания – фитофильные формы (мошки, хироно-миды и т.д.), отсутствующие в бентосе данного переката могут быть съедены рыбой только из толщи воды, что свидетельствует о преобладании у быст-рянки «пелагического» типа питания. Меньшее значение в питании имеют организмы, потребляемые рыбами непосредственно со дна – прикреплённые куколки ручейников, хирономиды, многочисленные на перекате.
Питание русской быстрянки р. Валы не зависит от пола и возраста. Различия наблюдаются в представленности дополнительных и случайных объектов питания.
Сравнивая питание русской быстрянки р. Валы и р. Чепцы (по нашим данным), участки которых отличаются по объёму вод, можно отметить, что по типу питания представители обеих популяций являются поли(эври)фагами. Однако есть существенная разница в представленности компонентов, формирующих основу пищевого комка. Так, в питании половозрелых особей русской быстрянки р. Чепцы как самцов, так и самок преобладает комплекс растительных компонентов, а именно нитчатой зеленой водоросли Ulothrix subtilissima Rabenhorst и детрита. В питании быстрянки р. Валы преобладает комплекс донных беспозвоночных.
Заключение
Изученный перекат р. Валы характеризуется невысокими видовым богатством, относительными численностью и биомассой рыб. Качественные и количественные показатели зообентоса, напротив, были достаточно велики. Основной компонент ихтиоценоза переката реки – русская быстрянка проявляет высокую степень полифагии. Ключевым компонентом питания, независимо от пола и возраста (2+ – 6+), выступали разные стадии развития мошки S. ornatum .
Автор выражает искреннюю признательность своему научному руководителю – чл.-корр., д.б.н. С.М. Голубкову (ЗИН РАН), директору Пермского отделения ФГБНУ «ГосНИОРХ» к.б.н. А.Г. Мельниковой, своим коллегам – к.б.н. И.В. Поздееву, С.П. Огородову, В.В. Безматерных, Н.А. Мартыненко.
Список литературы Бентофауна и ихтиофауна переката средней равнинной реки, населяемого русской быстрянкой Alburnoides rossicus Berg
- Алимов А.Ф. Элементы теории функционирования водных экосистем. СПб.: Наука, 2001. 147 с.
- Арнольди Л.В. Материалы по количественному изучению зообентоса Черного моря. Каркинитский залив//Тр. Севаст. биол. станции. 1949. Т. 7. С. 127-192.
- Балушкина Е.В. Функциональное значение личинок хирономид в континентальных водоемах. Л.: Наука, 1987. 180 с.
- Боруцкий Е.В. Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. М.: Наука, 1974. 254 с.
- Ивлев B.C. Экспериментальная экология питания рыб. М.: Пищепромиздат, 1955. 253 с.
- Комлев A.M., Черных Е.А. Реки Пермской области. Пермь: Кн. изд-во, 1984. 214 с.
- Котельникова B.C. Особенности питания русской быстрянки Alburnoides rossicus Berg, 1924.//Труды Зоологического института РАН, 2016. Т. 320. С. 326-335.
- Методика изучения биогеоценозов внутренних водоёмов. М.: Наука, 1975. 240 с.
- Нежиховский Р.А. Вопросы гидрологии реки Невы и Невской губы. Л.: Гидрометеоиздат, 1988. 224 с.
- Никольский Г.В. Теория динамики стада рыб. М.: Пищевая пром-сть, 1974. 447 с.
- Никольский Г.В. Структура вида и закономерности изменчивости рыб. М.: Пищевая пром-сть, 1980. 184 с.
- Определитель пресноводных беспозвоночных России и сопредельных территорий. Двукрылые/под ред. С.Я. Цалолихина. СПб.: Наука, 1999. Т. 4. 998 с.
- Правдин И.Ф. Руководство по изучению рыб. М.: Пищевая пром-сть, 1966. 376 с.
- Ресурсы поверхностных вод СССР. Основные гидрологические характеристики. Средний Урал и Приуралье. Вып. 1. Кама. Л., 1967. Т. 11. 536 с.
- Ручин А.Б. Пластические признаки быстрянок (Teleostei: Cyprinidae: Alburnoides) из различных частей ареала//Вестник Оренбургского университета. 2013. № 10 (159). С. 85-89.
- Чертопруд М.В. Разнообразие реофильных сообществ макробентоса//Материалы лекций П-й Всерос. шк.-конф./Институт биологии внутренних вод им. И. Д. Папанина. Ярославль: Филигрань, 2014. Том I. С. 112-124.
- Шитиков В.К., Розенберг Г.С. Рандомизация и бутстреп: статистический анализ данных по биологии и экологии с использованием R. Тольятти: Кассандра, 2013. 314 с.
- Щербина Г.Х. Годовая динамика макрозообентоса открытого мелководья Волжского плеса Рыбинского водохранилища//Зооценозы водоемов бассейна Верхней Волги в условиях антропогенного воздействия. СПб., 1993. С. 108-144.
- Horn HS. Measurement of «overlap» in comparative ecological studies//Amer. Natur., 1966. Vol. 100. P. 419-424.
- Hubbs C.L., Lagler K.F. Fishes of the Great Lakes Region. Ann Arbor: University of Michigan Press., 1958. 213 p.
- Hynes H.B.N. The ecology of running waters. Toronto, 1970. 555 p.
- lilies J. Versuch einer allgemeinen biozonotischen Gliederang der Fliessgewasser//Int. Revue Ges. Hydrobiol. 1961. Bd. 46, № 2. P. 205-213.
- Kottelat M., Freyhof J. Handbook of European Freshwater fishes. Berlin, 2007. 646 p.
- Lelek A. Threatened fishes of Europe. The freshwater fishes of Europe. Wiesbaden: Aula-Verlag, 1987. Vol. 9. 343 p.
- Raikova-Petrova G.N. et al. Growth Rate and Condition of Riffle Minnow (Alburnoides bipunctatus Bloch) from the Middle Stream of Iskar River (Bulgaria)//Acta Zoologica. 2011. Vol. 63, № 3. P. 295-300.
- Treer T. et al. Diet and growth of spirlin, Alburnoides bipunctatus in the barbell zone of the Sava River//Folia Zool., 2006. Vol. 55(1). P. 97-106.
- Wallace R.K. An assessment of diet-overlap indexses//Trans. Amer. Fish. Soc, 1981. Vol. 110. P. 2-76.
