Беспозвоночные песчаных массивов в Баргузинской котловине (Западное Забайкалье)
")
Автор: Хобракова Лариса Цыренжаповна, Убугунов Василий Леонидович, Рудых Сергей Геннадьевич
Журнал: Природа Внутренней Азии @nature-inner-asia
Рубрика: Байкальский регион, Россия
Статья в выпуске: 3 (4), 2017 года.
Бесплатный доступ
Установлен таксономический состав беспозвоночных животных, их численность и биомасса на разной стадии восстановления степных экосистем: язва дефляции, залежи и целина. Прослежены изменения мезофауны в различных ландшафтно-климатических условиях, которые выражаются в увеличении разнообразия, численности и биомассы травоядных беспозвоночных.
Баргузинская котловина, западное забайкалье
Короткий адрес: https://sciup.org/148317121
IDR: 148317121 | УДК: 595.762.12+574.472 | DOI: 10.18101/2542-0623-2017-3-90-101
Invertebrates of sanding massifs in Barguzin hollow (Western Transbaikale)
The taxonomic composition of invertebrate animals, their abundance and biomass at different stages of restoration of steppe ecosystems is established: deflation ulcers, deposits and virgin land. The changes in the mesofauna in various landscape and climatic conditions are traced, which are expressed in increasing diversity, abundance and biomass of herbivorous invertebrates.
Текст научной статьи Беспозвоночные песчаных массивов в Баргузинской котловине (Западное Забайкалье)
К обитанию на поверхности и в толще песков приспособлены представители особой экологической группы животных — псаммофилов. Рыхлость и подвижность песка, быстрое нагревание его днем и резкое охлаждение ночью, сухость верхних слоев и концентрация влаги на некоторой глубине создают своеобразные условия существования. В связи с этим у псаммофилов имеется ряд характерных особенностей. Некоторым свойственна быстрота передвижения (быстрый бег и полет, прыжки), как приспособление к добыванию пищи. Характерными представителями таких псаммофилов являются жуки-скакуны. Часто псаммофилы способны жить в толще песка. Например, личинки муравьиных львов форми- руют ловчую воронку, выставляя наружу лишь крючковидные челюсти, которыми ловят скатывающуюся вниз добычу (мелкие насекомые). Многие представители беспозвоночных среди жуков жужелицы, пластинчатоусые, чернотелки и долгоносики, среди перепончатокрылых дорожные и роющие осы, муравьи; часть саранчовых и пауков-волков являются характерными обитателями песчаных биотопов. Некоторые виды являются редкими и охраняются [Красная книга Бурятии, 2013]. Например, жужелица драгоценнокрылая Carabus glyptopterus, локально распространенная в степных котловинах Забайкалья, редкий вид на северной границе ареала [Хобракова, 2013].
Материал и методы
Исследования проводили в Баргузинской котловине, являющейся одной наиболее крупных тектонических депрессий Байкальского рифта. Впадина заполнена песками, большая часть из которых сосредоточена в обширные песчаные массивы, возвышаясь над озерно-аллювиальной до 200 и более метров. Зоологические исследования проведены в центральной части котловины на песчаном массиве «Верхний Куйтун», преимущественно занятом степной растительностью.
Материал по беспозвоночным животным собран методами почвенных ловушек. Почвенные ловушки выставлялись по 20 штук в линию через 5 м [Barber, 1931]. В качестве фиксатора использовали 4%-ный раствор формалина. Материал выбирался ежемесячно в течение всего вегетационного сезона в июне, июле, августе, сентябре в 6 биотопах. Собрано свыше 22 тысяч экземпляров имаго беспозвоночных общим весом 865,19 г (табл. 1).
Результаты изменения таксономической структуры, численности и биомассы на разной стадии восстановления степных экосистем
Стадия 1. Беспозвоночные на язвах дефляции
На первой стадии восстановления изучены два биотопа с различной степенью занесенности песками.
Песчаные выдувы, так называемые язвы дефляции, имеют максимальный характер занесенности песками в окружении тополевников (биотоп № 3). В этих условиях беспозвоночные представлены 1 типом, 2 классами, 10 отрядами из более 22 семейств. Здесь отмечено минимальное разнообразие беспозвоночных (31 таксон) с наименьшей численностью (2 135 экз., 9,5%). Среди них доминируют разные семейства жуков: жужелицы (32,5%), быстрянки (24,4%) и нарывники (11,7%). В числе субдоминантов в диапазоне от 5 до 10% представлены муравьи и двукрылые, реже встречаются, но обычны муравьиные львы и роющие осы. Биомасса беспозвоночных составила 66,56 г. По биомассе преобладают жужелицы (46,9%) и нарывники (26,3%).
Мозаичная язва дефляции среди степной растительности представлена биотопом №14. Сообщество беспозвоночных сформировано представителями 1 типа, 2 классов, 10 отрядов. В этих условиях возрастает разнообразие беспозвоночных (38 таксонов) и их численность (2 953 экз., 13%). Среди таксонов больше 1% в структуре доминирования по-прежнему преобладают жуки семейств жужелицы (33,9%), быстрянки (11,2%) и нарывники (9,3%). В составе субдоминантов пред- 91
ставлены муравьи, роющие осы, перепончатокрылые. Количество редких таксонов возрастает до 10, среди которых представлены прочие представители двукрылых, совок (гусеницы), дорожных ос, жуков долгоносиков, чернотелок, мертвоедов, карапузиков, саранчовых, клещей и пауков. Представители некрофиль-ного комплекса (мертвоеды, карапузики), очевидно, привлекаются попавшими в почвенные ловушки беспозвоночными. Из групп, играющих значительную роль в песчаных экосистемах, отсутствуют муравьиные львы. Биомасса данных животных в этом биотопе наибольшая и составляет 248,63 г. По биомассе преобладают жужелицы (70%).
Таким образом, на выдувах песчаных массивов формируется разнообразная фауна беспозвоночных, которая включает 45 таксонов на уровне семейств и отрядов с общей численностью 5 088 экземпляров (22,8% от общей численности всех изученных беспозвоночных), при этом только 13 таксонов больше 1%. Доминируют жужелицы (33,3%), быстрянки (16,8%), нарывники и муравьи (по 10,3%).
-
Стадия 2. Беспозвоночные на залежах
Изучен зоологический комплекс на залежах из двух биотопов.
Беспозвоночные на залежи из биотопа № 12 относятся к 1 типу, 10 отрядам и 30 семействам. Разнообразие беспозвоночных составляет 38 таксонов с численностью 4 522 экземпляров (20%). Среди таксонов больше 1% в структуре доминирования превалируют жужелицы (26,8%), быстрянки (16,8%), а также муравьи (12,5%) и нарывники (10,3%). В структуре субдоминантов отмечены только роющие осы и чернотелки, остальные встречаются реже — двукрылые, совки, дорожные осы, мертвоеды и саранчовые. Наряду с настоящими саранчовыми, нарывниками, чернотелками, муравьями значительную численность имеют дорожные осы, использующие пауков в качестве корма для своих личинок. Биомасса достигает 176 г. Вклад быстрянок (средний вес особи около 2 мг) в общие показатели биомассы невелик. По биомассе преобладают жужелицы (30,9%) и нарывники (18,5%).
На залежи (биотоп №11) население беспозвоночных, оцененное на основе материалов из почвенных ловушек, представлено 1 типом, 2 классами, 10 отрядами и 28 семействами. Разнообразие беспозвоночных этого биотопа не уступает предыдущему (37 таксонов), но здесь отмечена самая большая численность беспозвоночных в 7 347 экземпляров (33%). В структуре доминирования преобладают жужелицы (59,2%) и быстрянки (15,7%), в числе субдоминантов присутствуют чернотелки, остальные отмечены как редкие — двукрылые, совки, перепончатокрылые, муравьи, нарывники, полужесткокрылые и саранчовые. Общая биомасса составила 146,46 г. По биомассе преобладают жужелицы (30,69%) и чернотелки (21,52%).
Таким образом, на залежах разнообразие беспозвоночных составляет 41 таксон с максимальной численностью 11 869 экземпляров (53,3% от общей численности), из них только 10 таксонов больше 1%. Доминируют жужелицы (46,9%) и быстрянки (16,1%).
-
Стадия 3. Беспозвоночные на целине
На целине в биотопе №1 биоразнообразие беспозвоночных представлено 1 типом, 2 классами, 9 отрядами и 22 семействами. Всего отмечено 29 таксонов с 92
численностью 379 экземпляров (23,7%), среди них 15 таксонов больше 1% (рис. 5). В структуре доминирования представлены: доминанты — клопы (19%), равнокрылые, большей частью их личиночные формы (15,8%), гусеницы совок (12,1%); субдоминанты — жужелицы (9,5%), двукрылые (6,9%), клещи (6,6%), долгоносики (5,8%), саранчовые (5,5%). Среди жужелиц обычен степной вид Pseudotaphoxenus dauricus . Биомасса беспозвоночных — 17,56 г. По биомассе преобладают гусеницы совок (39,3%), жужелицы (11,3%), клопы (10,3%) и саранчовые (10,2%).
На целине в биотопе №18 таксономическое разнообразие беспозвоночных, принадлежащих к 1 типу, 3 классам, 11 отрядам, 31 семейству, является самым высоким из всех анализируемых биотопов. Зарегистрировано 42 таксона с численностью 5 292 экземпляра (23,7%), среди них 8 таксонов больше 1%. В структуре доминирования преобладают жужелицы (36,6%), быстрянки (19,7%) и саранчовые (15,9%). В структуре субдоминирования представлены муравьи, нарывники, реже встречаются двукрылые, перепончатокрылые, чернотелки (рис. 6). Традиционно многочисленны нарывники, чернотелки, муравьи. Заметную роль играют усачи-корнееды из рода Eodorcadion . Отсутствуют муравьиные львы. Только здесь обнаружены представители класса скрыточелюстных насекомых (ногохвостки). Биомасса составляет 209,98 г. Рекордных значений, как по численности, так и по биомассе достигают настоящие саранчовые (26,13%), также многочисленны нарывники (12,03%).
Таким образом, разнообразие беспозвоночных на целине представлено 1 типом, 3 классами, 11 отрядами и 31 семейством с общей численностью 5 671 экземпляров (25,5%). Доминируют жужелицы, быстрянки, саранчовые, нарывники.
Изменения мезофауны в различных ландшафтно-климатических условиях
На естественных, не затронутых вспашкой участках, с большим перепадом высот (150 м), отмечено увеличение мощности гумусового горизонта и, в целом, профиля в почвах, появление срединного палевого горизонта и меньшего содержание карбонатов в аккумулятивно-карбонатном горизонте.
Растительные сообщества реагируют на изменение почвенно-климатических условий с высотой — снижением общей продуктивности, ОПП, видового состава и резким увеличением абсолютной массы и доли надземной фитомассы по отношению к подземной. Возрастание надземной фитомассы ожидаемо привело к повышению числа видов и общей биомассы беспозвоночных.
По сравнению с целинными вариантами экосистем, антропогенное воздействие в виде пахоты и усиления дефляции, а затем залежное состояние в течение 20–30 (40) лет привело к неоднозначной реакции растительных сообществ на изменение почвенного плодородия. На залежах, не подверженных или слабо подверженных дефляции, число видов заметно возрастает, что характерно для соответствующих демутационных стадий, а на сильно и полностью дефлированных участках — резко сокращается. Общая биопродуктивность всех залежных фитоценозов сократилась независимо от степени дефляции почв, однако на недефли-рованных позициях надземная биомасса была сопоставима с теоретически возможным уровнем, а иногда его незначительно превышала.
Число видов и биомасса беспозвоночных на залежах, особенно на недефлиро-ванных, была выше, чем на целинных участках, сопоставимых по высотным отметкам (рис. 1, 2). Весьма интересным является, на наш взгляд, достаточно высокое обилие видов и их биомасса на язвах дефляции, практически полностью лишенных растительности, которые по значениям превышали или были равны теоретически возможным уровням.
Таким образом, увеличение гипсометрического уровня на 150 м приводит к локальному изменению климатических условий. В почвах с высотой увеличивается мощность гумусового и других генетических горизонтов, появляется палево-метаморфический гор. Горизонт палево-метаморфический (BPL), в растительном покрове снижается общая продуктивность, трансформируется долевое соотношение надземной и подземной фитомасс в пользу надземной, уменьшается число видов и общее проективное покрытие. Увеличение надземной фитомассы ведет к резкому увеличению видового состава и биомассы беспозвоночных.
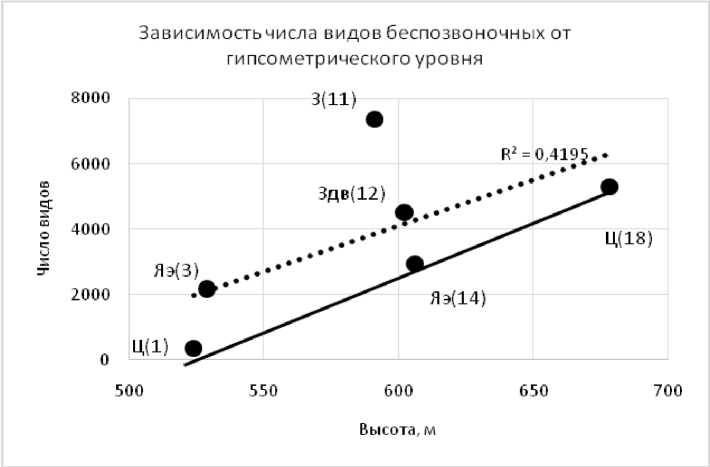
Рис. 1. Зависимость числа видов беспозвоночных от гипсометрического уровня поверхности
Условные обозначения (здесь и для рис. 2): сплошная линия — теоретическое увеличение числа видов при возрастании высотных отметок; точечная — линейный тренд, рассчитанный по всем изученным точкам. Степень антропогенного воздействия на экосистемы: Ц — целина; З — залежь; Здв — залежь с полностью дефлированным восстанавливающимся почвенным покровом; Яэ — язва эрозии.
Почвенные профили
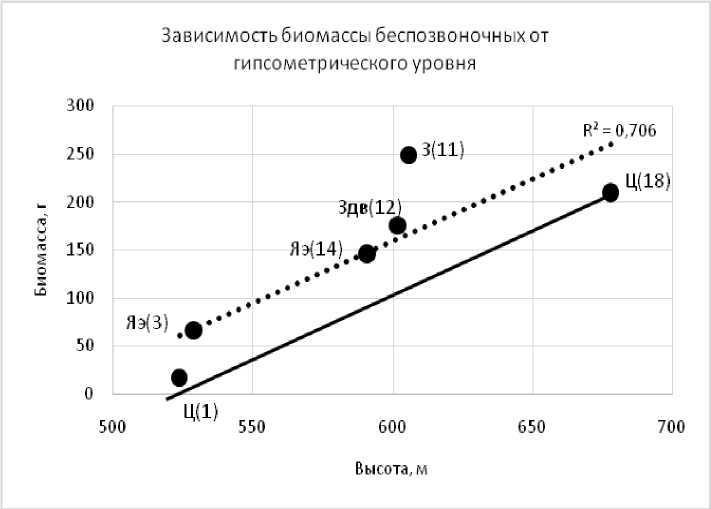
Рис. 2. Зависимость общей биомассы беспозвоночных от гипсометрического уровня поверхности. Условные обозначения (см. рис. 1)
Пахотное использование земель привело к дефляции почв и, как следствие, падению общей продуктивности фитоценозов, ОПП и сокращению их видового многообразия, особенно на сильно дефлированных позициях. Общая биомасса и число видов беспозвоночных на залежах намного выше, чем на целинных почвах с сопоставимыми высотами. Дефляционные нагрузки снижают эти показатели, но даже на язвах эрозии, лишенных растительности число видов и биомасса выше, чем на целинных территориях. Биомасса зависит, в основном, от численности жужелиц и тесно сопряжена с высотными отметками (r=0,84), независимо от степени антропогенной и дефляционной трансформации почвенно-растительного покрова, что также, но в несколько меньшей степени затрагивает и их видовое многообразие (r=0,65).
Таким образом, исследован таксономический состав беспозвоночных животных, их численность и биомасса на разной стадии восстановления степных экосистем: язва дефляции, залежи и целина. Фауна основных групп беспозвоночных состоит из представителей 1 типа, 3 классов, 11 отрядов, 46 семейств. Особенностью стадий восстановления является увеличение таксономического разнообразия, увеличение численности и биомассы травоядных беспозвоночных. По мере задернения и укрепления песчаных массивов в структуре доминирования по численности во всех биотопах преобладают степные виды беспозвоночных.
Таблица 1 Таксономический состав беспозвоночных в степных фитоценозах песчаного массива «Верхний Куйтун» на разной стадии восстановления
|
Стадии восстановления |
Язва эрозии (№3) |
Язва эрозии (14) |
Залежь (12) |
Залежь (11) |
Целина (1) |
Целина (18) |
||||||
|
Ч |
Б |
Ч |
Б |
Ч |
Б |
Ч |
Б |
Ч |
Б |
Ч |
Б |
|
|
ОТРЯД ПАУКИ (ARANEI) |
||||||||||||
|
Сем. Теридиды (Theridiidae) |
2 |
0,06 |
1 |
0,03 |
4 |
0,12 |
2 |
0,06 |
2 |
0,06 |
||
|
Сем. Пауки-волки (Lycosidae) |
15 |
1,50 |
56 |
4,65 |
41 |
3,07 |
55 |
4,40 |
8 |
0,52 |
10 |
0,75 |
|
Сем. Бродячие пауки (Gnaphosidae) |
5 |
0,10 |
2 |
0,05 |
6 |
0,18 |
17 |
0,43 |
1 |
0,03 |
8 |
0,24 |
|
Сем. Крабы (Thomisidae) |
0,03 |
3 |
0,025 |
3 |
0,03 |
|||||||
|
ОТРЯД ПАУКИ, ПРОЧИЕ СЕМЕЙСТВА |
2 |
0,06 |
2 |
0,03 |
8 |
0,15 |
5 |
0,08 |
9 |
0,14 |
||
|
ОТРЯД КЛЕЩИ (ACARI) |
||||||||||||
|
Сем. иксодовые клещи (Ixodidae) |
1 |
0,03 |
||||||||||
|
ОТРЯД КЛЕЩИ, ПРОЧИЕ СЕМЕЙСТВА |
1 |
0,005 |
34 |
0,11 |
30 |
0,09 |
5 |
0,03 |
25 |
0,07 |
27 |
0,08 |
|
Надкласс Насекомые (Insecta) |
||||||||||||
|
Класс Скрыточелюстные насекомые (Entognatha) |
2 |
0,02 |
||||||||||
|
Класс Открыточелюстные насекомые (Ectognatha) |
||||||||||||
|
ОТРЯД СТРЕКОЗЫ (ODONATA) |
||||||||||||
|
Сем. Настоящие стрекозы (Libellullidae) |
2 |
0,38 |
||||||||||
|
ОТРЯД ПРЯМОКРЫЛЫЕ (ORTHOPTERA) |
||||||||||||
|
Сем. Настоящие кузнечики (Tettigoniidae) |
1 |
0,05 |
8 |
0,56 |
||||||||
|
Сем. Настоящие саранчовые (Acrididae) |
9 |
0,45 |
51 |
3,57 |
123 |
9,23 |
130 |
9,1 |
21 |
1,79 |
844 |
54,86 |
|
ОТРЯД РАВНОКРЫЛЫЕ (HOMOPTERA) |
12 |
0,06 |
11 |
0,20 |
13 |
0,30 |
17 |
0,34 |
60 |
0,90 |
11 |
0,25 |
|
ОТРЯД ПОЛУ-ЖЕСТКОКРЫЛЫЕ (HEMIPTERA) |
||||||||||||
|
Сем. Клопы– хищники (Reduviidae) |
2 |
0,07 |
30 |
0,90 |
||||||||
|
Сем. Кружевницы (Tingidae) |
8 |
0,18 |
||||||||||
|
Сем. Булавники (Rhopalidae) |
1 |
0,04 |
||||||||||
|
Сем. Настоящие щитники (Pentatomidae) |
1 |
0,04 |
||||||||||
|
ОТРЯД ПОЛУ-ЖЕСТКОКРЫЛЫЕ, ПРОЧИЕ СЕМЕЙСТВА |
16 |
0,24 |
15 |
0,24 |
27 |
0,67 |
76 |
1,52 |
72 |
1,80 |
25 |
0,72 |
|
ОТРЯД ЖЕСТКОКРЫЛЫЕ (COLEOPTERA) |
||||||||||||
|
Сем. Жужелицы (Carabidae) |
693 |
31,19 |
1000 |
174,04 |
1210 |
54,45 |
4351 |
44,95 |
36 |
1,98 |
1935 |
87,080,24 |
|
Сем. Карапузики (Histeridae) |
56 |
1,30 |
24 |
0,48 |
8 |
0,12 |
1 |
0,02 |
18 |
0,36 |
||
|
Сем. Мертвоеды (Silphidae) |
34 |
3,74 |
76 |
8,36 |
35 |
4,20 |
8 |
0,88 |
||||
|
Сем. Стафилины (Staphylinidae) |
6 |
0,06 |
2 |
0,04 |
2 |
0,03 |
||||||
|
Сем. Пластинчатоусые (Scarabaeidae) |
11 |
0,33 |
3 |
0,12 |
1 |
0,04 |
2 |
0,06 |
1 |
0,05 |
||
|
Сем. Щелкуны (Elateridae) личинки |
1 |
0,06 |
2 |
0,07 |
||||||||
|
Сем. Кожееды (Dermestidae) |
1 |
0,025 |
1 |
0,02 |
||||||||
|
Сем. Нарывники (Meloidae) |
250 |
17,53 |
275 |
19,25 |
466 |
32,62 |
134 |
8,71 |
7 |
0,35 |
361 |
25,27 |
|
Сем. Быстрянки (Anthicidae) |
522 |
1,04 |
332 |
0,66 |
760 |
1,52 |
1151 |
2,30 |
2 |
0,005 |
1040 |
2,08 |
|
Сем. Чернотелки (Tenebrionidae) |
44 |
2, 21 |
49 |
2,70 |
221 |
12,16 |
573 |
31,52 |
15 |
0,45 |
182 |
10,01 |
|
Сем. Горбатки (Mordellidae) |
2 |
0,01 |
1 |
0,01 |
||||||||
|
Сем. Божьи коровки (Coccinellidae) |
1 |
0,03 |
||||||||||
|
Сем. Усачи (Cerambycidae) |
1 |
0,03 |
52 |
2,86 |
||||||||
|
Сем. Листоеды (Chrysomelidae) |
1 |
0,03 |
3 |
0,09 |
1 |
0,02 |
8 |
0,24 |
||||
|
Сем. Долгоносики (Curculionidae) |
44 |
1,98 |
3 |
0,12 |
33 |
1,32 |
22 |
0,88 |
22 |
0,88 |
||
|
Отряд Перепончатокрылые (Hymenoptera) |
||||||||||||
|
Сем. Осы-блестянки (Chrysididae) |
1 |
0,02 |
2 |
0,04 |
2 |
0,04 |
1 |
0,025 |
||||
|
Сем. Осы-немки (Mutillidae) |
1 |
0,03 |
2 |
0,06 |
3 |
0,09 |
||||||
|
Сем. Дорожные осы (Роmpilidae) |
2 |
0,10 |
60 |
2,99 |
47 |
2,35 |
40 |
2,05 |
1 |
0,05 |
6 |
0,30 |
|
Сем. Роющие осы (Sphecidae) |
38 |
2,85 |
183 |
10,98 |
249 |
14,94 |
56 |
3,92 |
4 |
0,26 |
41 |
2,67 |
|
Сем. Муравьи (Formicidae) |
208 |
1,05 |
318 |
1,59 |
563 |
2,25 |
196 |
0,98 |
14 |
0,07 |
272 |
1,23 |
|
Сем. Настоящие пчелы (Apidae) |
4,08 |
25 |
2,25 |
0,03 |
1 |
0,08 |
||||||
|
ОТРЯД ПЕРЕПОНЧАТО-КРЫЛЫЕ, ПРОЧИЕ СЕМЕЙСТВА |
102 |
0,10 |
148 |
5,75 |
325 |
11,38 |
152 |
6,08 |
6 |
0,025 |
93 |
3,72 |
|
ОТРЯД СЕТЧАТОКРЫЛЫЕ (NEUROPTERA) |
||||||||||||
|
Сем. Муравьиные львы (Myrmeleontidae) |
49 |
4,90 |
5 |
0,55 |
1 |
0,11 |
||||||
|
Сем. Myrmeleontidae, личинки |
1 |
0,03 |
||||||||||
|
ОТРЯД ЧЕШУЕКРЫЛЫЕ (LEPIDOPTERA) |
||||||||||||
|
Microlepidoptera |
2 |
0,005 |
12 |
0,06 |
7 |
0,03 |
2 |
0,01 |
3 |
0,015 |
4 |
0,02 |
|
Надсемейство Огневкообразные (Pyraloidea) |
8 |
0,16 |
2 |
0,05 |
2 |
0,04 |
|
Сем. Пяденицы (Geometridae), гусеницы |
1 |
0,08 |
6 |
0,42 |
1 |
0,08 |
||||||
|
Сем. Совки (Noctuidae) |
17 |
1,70 |
8 |
0,80 |
1 |
0,10 |
13 |
1,30 |
7 |
0,70 |
15 |
1,45 |
|
Сем. Совки (Noctuidae), гусеницы |
1 |
0,15 |
31 |
4,65 |
90 |
13,95 |
109 |
16,4 |
46 |
6,90 |
41 |
6,15 |
|
Сем. Голубянки (Lycaenidae) |
12 |
1,20 |
23 |
1,84 |
24 |
1,80 |
1 |
0,08 |
22 |
1,54 |
||
|
Сем. Нимфалиды (Nymphalidae) |
5 |
0,55 |
17 |
2,04 |
23 |
2,76 |
13 |
1,43 |
||||
|
Сем. Сатиры (Satyridae) |
3 |
0,45 |
2 |
0,30 |
1 |
0,15 |
||||||
|
ОТРЯД ДВУКРЫЛЫЕ (DIPTERA) |
||||||||||||
|
Сем. Журчалки (Syrphidae) |
3 |
0,06 |
5 |
0,13 |
2 |
0,035 |
2 |
0,04 |
2 |
0,05 |
||
|
Сем. Ктыри (Asilidae) |
7 |
0,28 |
3 |
0,12 |
12 |
0,48 |
26 |
1,04 |
||||
|
Сем. Жужжала (Bombyliidae) |
1 |
0,025 |
12 |
0.36 |
15 |
0,50 |
15 |
0,45 |
18 |
0,63 |
||
|
Сем. Тахины или Ежемухи (Tachinidae) |
2 |
0,08 |
14 |
0,80 |
14 |
0,75 |
2 |
0,11 |
4 |
0.22 |
||
|
Отряд Двукрылые, прочие семейства |
141 |
2,82 |
114 |
2,85 |
136 |
2,99 |
119 |
2,37 |
26 |
0,52 |
123 |
3,07 |
|
Итого |
2179 |
66,56 |
2953 |
248,63 |
4522 |
176 |
7347 |
146,46 |
379 |
17,56 |
5292 |
209,98 |
Примечание: Ч — численность (в экземплярах), Б — биомасса (в граммах).
Работа выполнена в рамках проектов «Эволюция, функционирование и экологобиогеохимическая роль почв Байкальского региона в условиях аридизации и опустынивания, разработка методов управления их продукционными процессами» (регистрационный номер: АААА-А17-117011810038-7; ФАНО 0337-2016-0005) и «Реакции животного мира Байкальского региона на глобальные изменения климата» (Регистрационный номер: АААА-А17-117011810035-6; ФАНО 0337-2016-0002).
Список литературы Беспозвоночные песчаных массивов в Баргузинской котловине (Западное Забайкалье)
- Barber H. Traps for cave-inhabiting insects // J. Elisha Mitchell Sci Soc. 1931. V.46. P. 259-266.
- Имехенова Т. К., Хобракова Л. Ц. К фауне насекомых заповедника Джергинский (Северное Прибайкалье) // Состояние и проблемы охраны природных комплексов Северо-Восточного Прибайкалья. Труды гос. заповедника «Джергинский». Вып. 2. Улан-Удэ: Изд-во Бурят. гос. ун-та, 1997. С. 71-79.
- Красная книга Республики Бурятия: Редкие и находящиеся под угрозой исчезновения виды животных, растений и грибов. 3-е изд., перераб. и доп. / отв. ред. Н. М. Пронин. Улан-Удэ: Изд-во БНЦ СО РАН, 2013. 688 с.
- Рудых С. Г. Булавоусые чешуекрылые Джергинского заповедника // Биоразнообразие экосистем Прибайкалья. Труды заповедника "Джергинский". Вып. 1. Улан-Удэ: Бурят. кн. изд-во, 1995. С. 65-69.
- Рудых С. Г., Власова Т. В. Животный мир. Раздел 4.1. Насекомые-Insecta // Природа заповедника «Джергинский». Улан-Удэ: Изд-во Бурят. гос. ун-та, 1998. С. 31-40.
- Хобракова Л. Ц., Шиленков В. Г., Дудко Р. Ю. Жуки-жужелицы (Coleoptera, Carabidae) Бурятии. Улан-Удэ: БНЦ СО РАН, 2014. 380 с.
- Шодотова А. А., Гордеев С. Ю., Рудых С. Г. Чешуекрылые Бурятии // Новосибирск: Издательство СО РАН, 2007. 250 с.
