Биогруппы в старых насаждениях сосны

Автор: Рогозин М.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
Исследована структура 180-летнего древостоя сосны обыкновенной (Pinus sylvestris L.) на площади 2.2 га. Измерены диаметры 736 деревьев на двух участках со средней высотой 32-33 м, запасами до 650 м3 /га и полнотами 0.92 и 1.07. В программе ArcGIS все деревья были нанесены на план в масштабе 1:100. На основе средней площади питания было рассчитано среднее расстояние между деревьями. Это расстояние использовали как единицу измерения силы взаимодействия между деревьями и выстраивали цепочки связей между ними. Приняли 7 уровней взаимодействия (конкуренции), для которых составили пять слоев карты. Оказалось, что в биогруппах, с расстоянием между соседями 84-270 см, растет в среднем 40% деревьев. Средние значения диаметров деревьев в них и в древостое отличаются недостоверно, не более чем на 1.0%. Даже в плотных биогруппах с расстоянием между соседями 84-150 см встречались наиболее крупные деревья, что противоречит известному и отрицательному влиянию высокой густоты древостоев на размер деревьев. Кроме того, пары диаметров деревьев в биогруппах коррелировали на уровне r = 0.47-0.58, что позволяет относить взаимодействие деревьев в них к партнерскому типу. Это явление меняет наше представление о древостоях, где наряду с конкуренцией есть и внутривидовое партнерство.
Сосна обыкновенная, древостой, конкуренция, биогруппы, партнерство
Короткий адрес: https://sciup.org/147227012
IDR: 147227012 | УДК: 581.9:
Biography in old stands of pine
To study the structure of 180-year-old stand of pine on the area of 2.2 hectares. Measure the diameters of 736 trees on two plots with an average height of 32-33 m, reserves of up to 650 cu. m/ha and fullness relative to the standard 0.92 and 1.07. In the ArcGIS computer program, all trees were plotted on a 1:100 scale. On the basis of the average area of food, the average distance between the trees was calculated. This distance was used as a unit of measurement of the force of interaction between trees and built a chain of links between them. Adopted 7 levels of interaction (competition), which amounted to five layers of the map. It turned out that in biogroups, with a distance between neighbors 84-270 cm, grows an average of 40% of trees. The average values of the diameters of the trees in them and in the forest differ untrue and not more than 1.0%. Even in dense biogroups with the distance between neighbors of 84-150 cm the largest trees were found, which contradicts the known and negative impact of high density of stands on the size of trees. In addition, the pairs of tree diameters in the biogroups were correlated at the level of r = 0.47-0.58, which allows us to relate the interaction of trees in the biogroups to the partner type. This new phenomenon changes our notion of stands, where, along with competition, there is also an intraspecific partnership.
Текст научной статьи Биогруппы в старых насаждениях сосны
Старые насаждения сосны обыкновенной (Pinus sylvestris L.) важны для представления о том, когда еще сохраняется их устойчивость. Вблизи г. Перми есть леса, где возраст деревьев превышает 200 лет [Рогозин, 2014; Рогозин, Андреев, 2014]. В них сохраняются и так называемые биогруппы, с предельно малым расстоянием (0.5–2.5 м) между деревьями. Среди российских ученых впервые их исследовал Н.Д. Лесков [Лесков, 1956]. Они представляют немалый интерес, так как деревья, оставшиеся в одиночестве после их разреживания при рубках ухода, существенно не увеличивали прирост в сравнении с контролем [Сеннов, 1999].
Однако этот факт, противоречащий обычным представлениям о действии конкуренции, которая наиболее сильна именно в биогруппах, не нашел приемлемого объяснения. Казалось вполне очевидным, что чем ближе деревья располагаются друг к другу, тем она сильнее и тем меньше по размерам будут деревья в местах усиления конкуренции. Но, несмотря на ее ослабление после рубок ухода с целью более равномерного размещения деревьев, в таких ухоженных лесах со временем все равно образовывались неравномерности в структу-
ре насаждения – близко расположенные деревья в группах, а также прогалины [Нестеров, 1961].
Наличие биогрупп, где формируется 28–57% деревьев в самых разных насаждениях, является безусловным атрибутом древостоя [Рогозин, Разин, 2015]. В связи с этим некоторые исследователи предлагают изменить теорию лесоводства и заложить в его основу принцип группового размещения деревьев; разработан даже учебный курс «Нетрадиционное лесоводство» [Марченко, Марченко, 1998]. Усиление конкуренции при более плотном размещении растений, согласно общим законам экологии и развития популяций [Реймерс, 1994], снижает размеры особей. Эту закономерность морфогенеза древостоев давно используют в моделировании их развития [Разин, 1979]. Из нее следует, что если в лесу будут густые места, то диаметры деревьев в них теоретически должны быть меньше.
В этой логике закономерность уровня фитоценоза перемещают на уровень микроценоза. Этот кажется справедливым, потому что упомянутой закономерности морфогенеза Г.С. Разина [Разин, 1979] был присвоен даже ранг закона [Рогозин, Разин, 2015] – наряду с еще двумя законами: законом естественного изреживания [Морозов, 1949] и ранговым законом роста деревьев Е.Л. Маслакова [Маслаков, 1984]. Однако в биогруппах все сложнее. С раннего возраста есть рекомендация оставлять их, если деревья в них крупные [Большакова, 2007].
Цель данной статьи – анализ влияния деревьев сосны друг на друга в биогруппах и в насаждении в целом в возрасте 180 лет.
Материал и методы
Материал
Наиболее старый лес найден в Кировском районе г. Перми, в кв. 1 бывших лесов завода им. С.М. Кирова, южнее остановки автобуса «ТЭЦ14», где обнаружены деревья с диаметрами до 68 см. Возраст определили по кернам древесины у 6 крупных деревьев, и число слоев на высоте ствола 1.3 м колебалось от 173 до 175 лет. Получается, что данная территория была пройдена рубками в 1830-е гг., и возраст деревьев, с учетом достижения высоты 1.3 м за 7 лет, составляет не менее 180 лет. Древостой оставляет хорошее впечатление; его структура не нарушена, так как при уборке отпавших деревьев пользовались прогалами между деревьями. Данный участок, как лес-эталон, используется для практики студентов по лесоведению. Здесь на участке 1 «Южный» и участке 2 «Северный» площадью 1.1102 и 1.1137 га соответственно были пронумерованы 736 деревьев и нанесены на план с точностью ±10–35 см, что опреде- лено при перекрестных линейных засечках, а также после распределения невязки на общем полигоне территории в масштабе 1:100. Взаимное расположение деревьев в биогруппах определялось более точно (±3–5 см). Таксационная характеристика яруса сосны следующая.
Участок 1 «Южный»: состав 10С, средняя высота по трем средним деревьям 32 м, средний диаметр по площади сечения 39.8 см, сумма площадей сечения 42.2 м2/га, полнота 0.92, запас 560 м3/га. Единично встречается ель 120 лет с диаметрами 20–45 см. Тип леса сосняк кисличниковый, в кустарничковом ярусе черника (70%) и брусника (30%). Местоположение ровное, с поднятием территории на 1.5 м в западной части; тип условий «свежий бор» (В 2 ), почва супесчаная, класс бонитета 1. Текущая густота 339 шт./га.
Участок 2 «Северный» близок по условиям, но отличается понижением территории на север на 1– 2° и несколько более влажными условиями, что определено косвенным путем по увеличению в кустарничковом ярусе черники до 95%. При высоте сосны в 33 м здесь показатели несколько выше: средний диаметр по площади сечения 42.06 см, сумма площадей сечения 45 м2/га, полнота 1.07, запас 650 м3/га. Здесь также единична ель с диаметрами 16–33 см. Густота сосны 324 шт./га, что на 4.6% меньше, чем на участке 1.
На обоих участках есть подрост ели густотой 0.3-1.5 тыс. шт./га, высотой 2–5 м в возрасте 35 лет, который появился после низового пожара 1982 г. Санитарное состояние сосны, с учетом её возраста, хорошее; имеются стволы с плодовыми телами гриба сосновая губка (9%). В 2003 г. (14 лет назад) здесь прошел ветровал, в результате которого было вывалено 6.8% деревьев. Частоты по ступеням диаметра идеально соответствуют нормальному распределению, и в целом древостой отвечает всем канонам «нормального» насаждения.
Методы исследования
Для изучения структуры древостоя наносили на план цепочки взаимодействий между деревьями, соединяя их линиями. При средней густоте 332 дерева на 1 га площадь питания одного дерева составила 30.1 м2, и квадратный корень из этой величины дает среднее расстояние между деревьями (Lср), равное 5.5 м, которое использовали как ориентир для градуировки взаимодействий между парами деревьев (табл. 1).
Вместо известных полигонов питания [Наги-мов, 2000; Вайс, 2014] в данном исследовании мы применили цепочки взаимодействия между деревьями и отразили их на разных слоях карты. Их роль становится понятной, если считать, что при малом расстоянии взаимодействие сильнее, а его усиление пропорционально квадрату отношения расстояний (Lср / L)2, показанных в табл. 1.
Таблица 1
Градуировка конкуренции (взаимодействия) между соседствующими парами деревьев, при среднем расстоянии между деревьями в древостое Lср = 5.5 м
|
Предел расстояний (L), от – до, м |
Интервал |
Отношение Lср / L |
(Lср / L)2 |
Градация конкуренции |
Слой карты |
||
|
min |
max |
в среднем |
|||||
|
0.8 – 1.3 |
0.5 |
6.88 |
4.23 |
5.55 |
30.83 |
Очень высокая |
Слой 1 «био- |
|
1.31 – 2.70 |
1.4 |
4.20 |
2.04 |
3.12 |
9.72 |
Высокая |
группы» |
|
2.71 – 3.70 |
1.0 |
2.03 |
1.49 |
1.76 |
3.09 |
Повышенная |
Слой 2 |
|
3.71 – 6.03 |
2.3 |
1.48 |
0.91 |
1.20 |
1.43 |
Средняя |
Слой 3 |
|
6.04 – 8.75 |
2.7 |
0.91 |
0.63 |
0.77 |
0.59 |
Пониженная |
Слой 4 |
|
8.76 – 12 |
3.2 |
0.63 |
0.46 |
0.54 |
0.29 |
Низкая |
Слой 5 «Прога- |
|
12.1 – 16 |
3.9 |
0.45 |
0.34 |
0.40 |
0.16 |
Очень низкая |
лины» |
Для градуировки расстояний был выбран неравномерный интервал с целью получения примерно равного числа связей на слоях карты. Кроме того, за отправную точку отделения «биогрупп» от остальных связей послужило расстояние между деревьями в 50% от Lср = 5.5 м, принятое равным 2.7 м для обоих участков.
Первый слой карты оказался близок к параметрам выделения биогрупп, обсуждаемых в литературе (0.5–2.5 м) [Ипатов, Тархова, 1975; Марченко, 1995; Марченко, Марченко, 1998; Рогозин, 2014; Рогозин, Голиков, Разин, 2014].
Результаты
Биогруппы участка «Южный»
До ветровала 14 лет назад на участке было 83 биогруппы, где сформировалось 191 дерево при расстоянии между ними 84–270 см. После ветровала их осталось 78 и сейчас в них растет 171 дерево, что составляет 45% числа живых деревьев. В них измерено 111 расстояний между деревьями и среднее равно 204.3±4.6 см. Это в 2.64 раза меньше, чем среднее расстояние между деревьями, рассчитанное исходя из квадрата средней площади питания, и равное здесь 5.4 м. Используя это сокращение расстояния в 2.64 раза и возводя его в квадрат, можно оценить возрастание конкуренции в 7 раз (2.642 ≈ 7.0).
Так как конкуренция резко возросла, то мы ожидали снижения среднего диаметра деревьев в биогруппах. Однако этого не произошло.
Среднеарифметический диаметр у 171 дерева в биогруппах оказался равен 39.02±0.62 см, что практически совпало со среднеарифметическим диаметром у сосны на всем участке, равном 39.10±0.39 см.
Биогруппы участка «Северный»
Здесь среди 361 живых и 23 вываленных в 2003 г. деревьев обнаружено 61 биогруппа, где формировались 129, или 33.6% деревьев. После ветровала их осталось 59 и сейчас в них растет 35% деревьев. В биогруппах измерено 69 расстояний между деревьями и среднее расстояние между ними оказалось равно 1.976 м. Площадь питания 1-го дерева равна здесь 31.1 м2, и такую площадь имеет квадрат со стороной 5.58 м. Принимая это расстояние для оценки, можно определить сокращение расстояния в биогруппах в 5.58/1.976 = 2.82 раза. Возводя это сокращение в квадрат, получаем усиление конкуренции в биогруппах в 2.822 ≈ 8 раз.
В биогруппах среднеарифметический диаметр оказался равен 40.85±0.72 см, а среднеарифметический диаметр деревьев на всем участке – 41.25±0.43 см, что всего лишь на 1.0% больше, и это различие недостоверно.
Оценка конкуренции
В биогруппах встречались очень разные деревья – как самые толстые (47–65 см), так и самые тонкие (24–30 см). Чтобы понять механизм их взаимодействия в биогруппах, было использовано отношение Дмакс / Дмин, где Дмакс – большее дерево и Дмин – меньшее. Затем сравнивали это отношение с таким же отношением каждого дерева из биогруппы с ближайшим соседом вне биогруппы, которое считалось контрольным.
Выдвинули гипотезу, что если это отношение в биогруппах больше, то в них будет более сильная конкуренция. Для ее проверки на участке «Южный» сформировали выборку из 56 биогрупп, состоящих из 2 деревьев, а также контроль из такого же числа соседних деревьев (рис. 1).
Сравнение выборок показывает, что их средние диаметры почти равны (соответственно, 39.0 и 39.2 см в опыте и контроле) несмотря на громадные различия в среднем расстоянии между деревьями – 195 и 525 см (табл. 2).
Сокращение расстояния в биогруппе в сравнении с контрольными соседями составило
525 / 195 = 2.7 раза. Считая, что конкуренция дей-
на соотношения диаметров входящих в нее деревь- ствует пропорционально квадрату расстояния, по- ев не влияет. Это весьма неожиданно, и получается лучаем ее усиление в 2.72 = 7.3 раза. Так как полу- явный парадокс – при усилении конкуренции в 7– ченные в таблице отношения Дмакс / Дмин оказа- 8 раз в биогруппах деревья почему-то не реагиру-лись практически равны (1.272 и 1.274), то из это- ют на это усиление.
го следует, что усиленная в биогруппе конкуренция
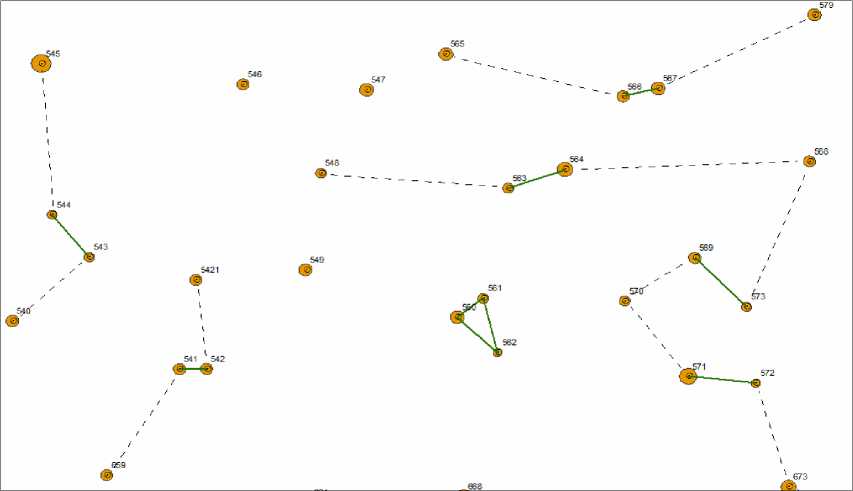
Рис. 1 . . Биогруппы и контрольные деревья: пунктирные линии – связь дерева из биогруппы с контрольным деревом
Таблица 2
Показатели деревьев в 56 биогруппах, состоящих из 2 деревьев, и в контроле в 180-летнем древостое на участке «Южный»
|
Показатель |
Статистики |
|||||
|
Число наблюдений, шт. |
Среднее |
Ошибка |
Вариация, % |
Минимум |
Максимум |
|
|
Биогруппы из 2 деревьев |
||||||
|
Диаметр, см |
112 |
39.0 |
0.76 |
20.6 |
24 |
65 |
|
Расстояние, см |
56 |
195 |
7.0 |
26.8 |
84 |
271 |
|
Дмакс/Д мин |
56 |
1.272 |
0.03 |
20.1 |
1.01 |
2.14 |
|
Контрольные деревья-соседи |
||||||
|
Диаметр, см |
112 |
39.2 |
0.70 |
18.9 |
25 |
63 |
|
Расстояние, см |
112 |
525 |
19.0 |
38.4 |
126 |
1427 |
|
Дмакс/Д мин |
112 |
1.274 |
0.022 |
18.2 |
1.00 |
2.29 |
Далее рассмотрим тренды: а) зависимость диаметра крупного дерева от расстояния между деревьями в биогруппе и б) ту же зависимость для минимального дерева. Вполне естественно, что первый тренд пойдет выше, а второй ниже (рис. 2).
На участке «Южный» для минимальных деревьев корреляция оказалась недостоверна (r = 0.15±0.13), но тренд для максимальных деревьев был достоверен (r = 0.33±0.12 при t = 2.8 > t 0.99 = 2.7). Аппроксимация линии тренда равна R2 = 0.1077, т.е. увеличение расстояния между соседями увеличивает диаметр крупного дерева с силой примерно в 11% (но диаметр мелкого дерева при этом достоверно не увеличивается).
Однако на участке «Северный» все эти тренды просто исчезают (см. рис. 2, б). Напомним, что этот участок по косвенным признакам (долей черники в кустарничковом ярусе напочвенного покрова 95%) более влажный. Это имело, вероятно, решающее значение для исчезновения слабых трендов, имевших место в более сухих условиях участка «Южный», где доля брусники возрастала до 30%, и об этих отличиях мы еще скажем далее.
Если по двум деревьям рассчитать средний диаметр дерева в биогруппе, то получим для участка «Южный» слабую корреляцию этого среднего диаметра с расстоянием между деревьями (r =
0.29), а на участке «Северный» эта корреляция нулевая и равна r = –0.003 (рис. 3).
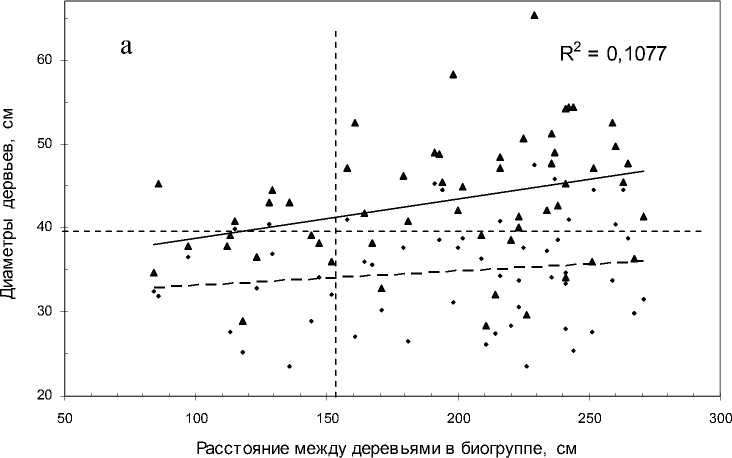
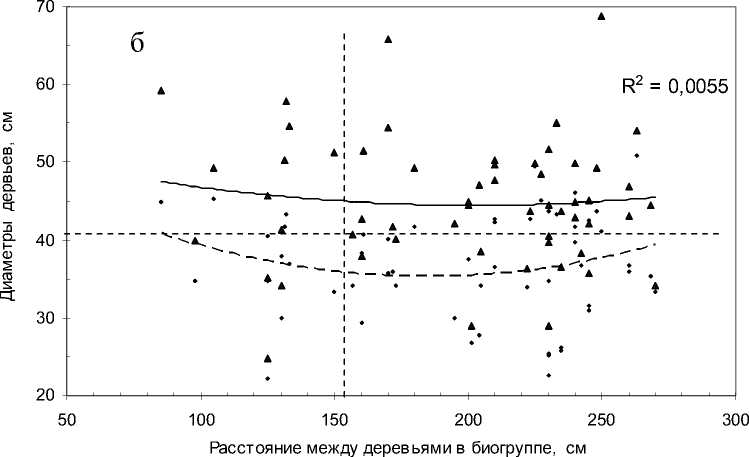
Рис. 2. Расстояние между деревьями в биогруппе и их диаметры на участках: а – «Южный»; б – «Северный»:
сплошная линия – тренд для деревьев с максимальным, прерывистая – с минимальным диаметром. Треугольные точки – деревья с максимальным диаметром. Горизонтальная линия – средний диаметр деревьев
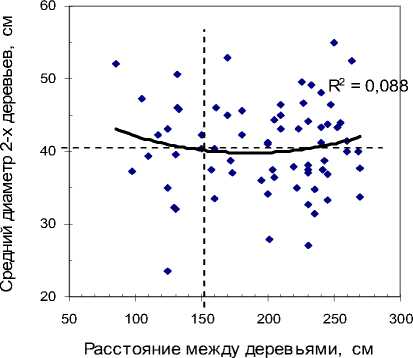
Принято биогруппы подразделять на «плотные», с расстоянием между деревьями до 150 см, и «менее плотные» с большим расстоянием [Ипатов, Тархова, 1975; Марченко, 1995]. Примерно по этому расстоянию (153 см) как раз и проведена вертикальная линия на рисунках, и вправо от нее на участке «Южный» (см. рис 3, а) начинают встречаться наиболее крупные деревья. Аппроксимация линии тренда низкая (R=0.088) и тем не менее, если разделить деревья на две выборки по этой вертикальной линии, то в левой средний диа- метр окажется 35.9±1.03 см, что меньше на 10.2% среднего диаметра в выборке справа (40.0±0.93 см). Различие достоверно при t = 2.9 > t0.99= 2.7.
В некоторых плотных биогруппах в лесу наш взгляд иногда сразу замечает это снижение диаметра у одного из деревьев. Ощущение, что одно из них «мешает» другому, вероятно, и появилось у лесоводов, тонко подметивших это уменьшение там, где расстояние между деревьями было менее 150 см, что хорошо видно на диаграмме слева. Ве- роятно, это наблюдение превратилось далее в убеждение, отвечающее ожиданиям выгоды от вырубки таких «мешающих» деревьев, а далее и в некий постулат рубок ухода, уже не нуждающийся в доказательствах (деление деревьев на три категории
– перспективные, вспомогательные и удаляемые). Напомним, однако, что вывод этот справедлив пока только для участка 1 (участка «Южный»), где тип леса несколько суше.
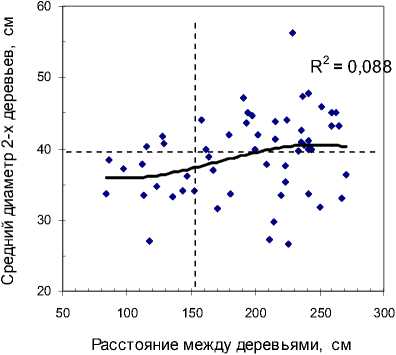
а
Рис. 3 . Влияние расстояния между деревьями в биогруппе из 2 особей на средний диаметр этой пары деревьев: а – участок «Южный»; б – участок «Северный»
б
Связь размеров деревьев в биогруппе
Продолжим анализ и выясним, не связаны ли диаметры деревьев в биогруппах между собой. Их коррелирование дало совершенно неожиданные результаты (рис. 4).
Так, на участке «Южный» пары деревьев в биогруппах оказались связаны между собой корре- ляцией диаметров r = 0.47. Заметим, что своеобразие тренда для полинома 3-й степени на рис. 4. а не стоит переоценивать – он сразу меняется при очистке выборки, если убрать пять точек (5% данных), образующих некий кластер справа внизу; после этого даже линейная связь поднимается до высокой при корреляции r = 0.73.
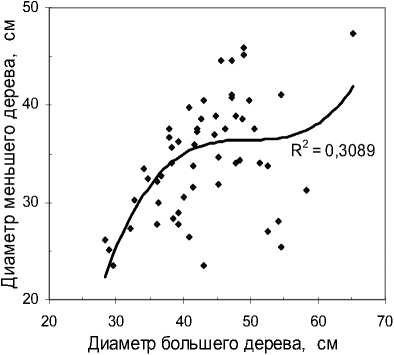
а
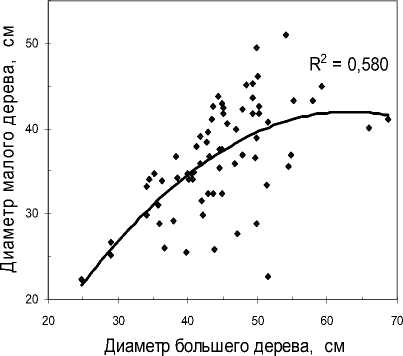
Рис. 4 . Корреляция диаметров сосны в биогруппах, состоящих из двух деревьев: а – участок «Южный»; б – участок «Северный»
б
На участке «Северный» корреляция пар оказалась несколько выше и равна r = 0.58 (рис. 4, б), а при очистке выборки в 5% также выросла до 0.70. При столь сильных связях в таксации уже моделируют объемы крон по диаметру дерева, определя- ют среднюю высоту насаждения по трем средним деревьям и определяют другие показатели для таблиц, нужных для практических целей.
Обнаруженная корреляция пар диаметров деревьев, растущих в биогруппах, и показанная на рис. 4, за пределами этих биогрупп сразу же исчезает. Так, при увеличении расстояний между парами деревьев до 271–375 см диаметры коррелируют очень слабо и отрицательно, причем на пределе достоверности связи (r = - 0.19±0.09), а далее, при расстояниях 376 см и более, связь между парами деревьев по их диаметрам уже недостоверна (r = 0.04).
Обсуждение результатов
Обнаруженная корреляция пар диаметров деревьев в биогруппах – самая большая загадка. Что она означает? Например, она может говорить о том, что условия для развития обоих деревьев близки, так как малое расстояние увеличивает вероятность попадания в сходные условия. Также и возраст деревьев может быть одинаковым. Но какие еще могут быть факторы – почвенные, световые, отличия в глубинных почвенных горизонтах или, может быть, локальное излучение энергии Земли в месте, где растет биогруппа?
Если условия плохие, то оба дерева синхронно снижают, а если хорошие, то увеличивают размеры. Но явных признаков богатой почвы по растениям-индикаторам в местах с крупными деревьями или бедной почвы вблизи мелких пар деревьев мы не обнаружили, по крайней мере, при визуальном осмотре напочвенного покрова. Не находили таких отличий и другие лесоводы и селекционеры, в частности, при выделении плюсовых деревьев в окружении конкурирующих соседей.
Вторым неожиданным явлением в биогруппах является разное влияние расстояний между деревьями на их средний диаметр при изменении увлажнения почвы. На более влажном участке «Северный» увеличение расстояний между деревьями не влияло на средний диаметр этой пары деревьев, тогда как на участке «Южный» с более сухой почвой это влияние имело место при корреляции r = 0.29. Отличие может быть случайным и если так полагать, то искать другие объяснения этих различий уже не нужно. Но есть и иное объяснение, интересное в генетическом плане и открывающее новые направления работ по изучению взаимодействий между деревьями.
Дело в том, что каждая популяция состоит из двух популяций-изомеров [Голиков, 2014], или левых и правых форм деревьев. Они имеют доказанные генетические отличия и противоположную адаптацию: левые предпочитают прямой свет и слабую конкуренцию, а правые – рассеянный и то-лерантны к конкуренции. Кроме того, левые лучше растут в сухих условиях, а правые – во влажных. В оптимальных условиях их частота одинакова, но если древостой становится густым, то начинают доминировать правые, а если он более редкий – левые формы. И они доминируют почти всегда, когда густота соответствует их предпочтениям, причем даже в несвойственных условиях по влажности. Частота этих форм выступает как основной механизм гомеостаза популяции при изменении ее плотности (густоты стояния деревьев) в процессе развития древостоя [Голиков, 2014; Рогозин, Голиков, Разин, 2014].
Вероятно, именно этот механизм, скорее всего, и приводит к тому, что в чуть более сухих условиях на участке «Южный», где правые формы расти «не любят» и они снижают там свои размеры; но одновременно им приходится преобладать в биогруппах из-за более высокой конкуренции, к которой они более приспособлены (см. рис. 3, а). В оптимальных условиях увлажнения сосняка кислич-никового на участке «Северный» правые формы чувствуют себя уже лучше, и снижения диаметров в плотных биогруппах у них уже не происходит (см. рис. 3, б).
Все эти новые направления работ нуждаются, безусловно, в тщательной проверке именно в биогруппах, где взаимодействия между деревьями наиболее сильны, и это при том, что в биогруппах растет чуть ли не половина деревьев.
Выводы
-
1. В биогруппах с расстоянием между соседями 84–270 см растет в среднем 40% сосны.
-
2. Среднеарифметические диаметры деревьев в биогруппах и во всем насаждении отличаются недостоверно и не более чем на 1.0%.
-
3. На участке с более сухими условиями при расположении соседей в плотных биогруппах с расстоянием 150 см и менее средний диаметр деревьев снижается на 8.2% в сравнении со средним на участке. Однако на участке с оптимальными условиями увлажнения такого снижения нет.
-
4. Диаметры деревьев в биогруппах коррелируют на уровне r = 0.47–0.58, что позволяет отнести их взаимодействие к партнерскому типу.
Заключение
Изучение структуры расположения 736 деревьев сосны на двух участках 180-летнего насаждения 1-го класса бонитета, отличающихся увлажнением (сосняк чернично-брусничниковый и сосняк кис-личниковый), с высокой относительной полнотой и с анализом диаметров деревьев в 137 микросообществах (биогруппах) показало, что встречаемость в них крупных деревьев входит в противоречие с известным и закономерным снижением размеров деревьев при повышении густоты в целом по древостою. Обнаружено новое явление во взаимоотношениях между деревьями – внутривидовое партнерство в биогруппах. Это меняет наше представление о древостое как сообществе, где дейст- вует одна лишь конкуренция. Подобное партнерство, по-видимому, будет обнаружено и у других древесных пород.
Автор благодарит сотрудников лаборатории геологического моделирования и прогноза ЕНИ ПГНИУ и лично к.г.н. П.А. Красильникова за консультации по составлению электронных карт.
Список литературы Биогруппы в старых насаждениях сосны
- Большакова Н.В. Влияние густоты и размещения посадочных мест на рост ели при выращивании культур по интенсивным технологиям: автореф. дис.... канд. с.-х. наук. СПб., 2007. 24 с
- Вайс А.А. Научные основы оценки горизонтальной структуры древостоев для повышения их устойчивости и продуктивности (на примере насаждений Западной и Восточной Сибири): автореф. дис.....д-ра с.-х. наук. Красноярск, 2014. 33 с
- Голиков А.М. Эколого-диссимметрический подход в генетике и селекции видов хвойных. Саар-брюкен: LAP LAMBERT Academic Publishing, 2014. 162 с
- Ипатов В.С., Тархова Т.Н. Количественный анализ ценотических эффектов в размещении деревьев по территории // Ботанический журнал. 1975. Т. 60, № 9. С. 1237-1250
- Лесков Н.Д. Особенности таксационной характеристики и структуры биогрупп в типе леса бор-брусничник // Труды по лесному хозяйству Уральского лесотехнического института. Свердловск, 1956. С. 56-60
- Марченко И.С. Биополе лесных экосистем. Брянск,1995. 188 с
- Марченко И.С., Марченко С.И. Нетрадиционное лесоводство: авторский курс / ред. Е.С. Мурах-танов. Брянск, 1998. 419 с
- Маслаков Е.Л. Формирование сосновых молодня-ков. М.: Лесн. пром-сть, 1984. 168 с
- Морозов Г.Ф. Учение о лесе / под ред. В.Г. Нестерова. М.: Гослесбумиздат, 1949. 453 с
- Нагимов З.Я. Закономерности роста и формирования надземной фитомассы сосновых древосто-ев: дис.... д-ра с.-х. наук. Екатеринбург, 2000. 409 с
- Нестеров В.Г. Вопросы современного лесоводства. М.: Госсельхозиздат, 1961. 384 с
- Разин Г.С. Динамика сомкнутости одноярусных древостоев // Лесоведение. 1979. № 1. С. 23-25
- Реймерс Н.Ф. Экология (теории, законы, правила, принципы и гипотезы). М.: Журнал «Россия молодая», 1994. 367 с
- Рогозин М.В. Высочайшие деревья Пермского края // Вестник Пермского университета. Сер. Биология. 2014. Вып. 3. С. 21-26
- Рогозин М.В., Андреев Д.Н. Туризм в лесах Пермского края // Географiя та туризм. Науковий збiрник. Кшв, 2014. Вип. 28. С. 82-90
- Рогозин М.В., Голиков А.М., Разин Г.С. О выращивании леса на сухих почвах: теоретические подходы // Вестник Поволжского гос. технологического университета. Сер. Лес. Экология. Природопользование. 2014. № 3 (23). С. 5-17
- Рогозин М.В., Разин Г.С. Развитие древостоев. Модели, законы, гипотезы [Электронный ресурс]: монография / под ред. М.В. Рогозина. Пермь, 2015. 277 с. URL: http://elibrary.ru/item.asp?id=24420793 (дата обращения: 25.11.15)
- Сеннов С.Н. Итоги 60-летних наблюдений за естественной динамикой леса. СПб.: СПбНИИЛХ, 1999. 98 с
