Биохимические исследования ряда промысловых видов рыб прибрежной зоны Баренцева моря в условиях антропогенного загрязнения

Автор: Овчинникова С.И., Широкая Т.А.
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Статья в выпуске: 3 т.7, 2004 года.
Бесплатный доступ
В статье приводятся результаты биохимических исследований (период 1998-2003 гг.) ряда промысловых рыб, характерных для водных экосистем прибрежной зоны Баренцева моря, с позиций оценки состояния гидробионтов как компонентов водных экосистем Кольского Севера в условиях антропогенного загрязнения. Показано, что динамика содержания основных химических показателей (влаги, общего азота, белкового азота, небелкового азота, аминного азота, водорастворимого белка, липидов, минеральных веществ и др.) в исследованных гидробионтах носит сезонный характер. Собственные данные, полученные на базе лабораторий биохимии МГТУ, сравнивали с результатами лаборатории биохимии гидробионтов Полярного научно-исследовательского института морского рыбного хозяйства и океанографии им. Н.М. Книповича за предыдущий период (до 1998 г.).
Короткий адрес: https://sciup.org/14293643
IDR: 14293643
Текст научной статьи Биохимические исследования ряда промысловых видов рыб прибрежной зоны Баренцева моря в условиях антропогенного загрязнения
Хозяйственное освоение морей сопровождается ростом антропогенного воздействия на морские экосистемы. В этом отношении наиболее уязвимы экосистемы арктических морей.
Водные экосистемы Кольского Севера, отличаясь повышенной чувствительностью к воздействиям, наиболее остро испытывают антропогенную нагрузку. Баренцево море представляет собой важную часть арктической среды. На его долю приходится более 13 % от общего вылова рыбы и гидробионтов в России.
Вовлечение Баренцева моря в сферу хозяйственной деятельности человека определило высокую интенсивность антропогенного воздействия на природную среду водоема. С потоком атлантических вод в него транспортируются загрязняющие вещества, источниками которых являются нефтепромыслы Северного моря, промышленные и сельскохозяйственные предприятия стран Западной Европы. Кроме того, загрязняющие вещества поступают с атмосферными осадками, речными стоками и в результате хозяйственной деятельности на его акватории. Повышенное содержание токсикантов в воде, донных отложениях и гидробионтах приводит к снижению продуктивности водных экосистем и к потенциальной опасности для человека.
В первую очередь загрязнению подвержены заливы и губы Баренцева моря, в пределах которых расположены города и поселки, где ведется производственная и хозяйственная деятельность. В этом плане особое место принадлежит Кольскому заливу – самому крупному фиорду Кольского полуострова, который является рыбохозяйственным водоемом высшей категории, при этом состояние Кольского залива и его бассейна уже сравнительно давно характеризуется достаточно высоким уровнем загрязнения.
Поэтому актуальной проблемой является оценка воздействия загрязнения на состояние гидробионтов. Возможным путем к ее решению являются биохимические исследования гидробионтов как компонентов водных экосистем Кольского Севера, т.к. организмы способны очень быстро отвечать на изменения факторов окружающей среды перестройкой химического состава и биохимических механизмов, что позволяет использовать данные показатели функционального состояния организмов для оценки биологического эффекта загрязнения ( Агатова и др. , 2001). Биохимический мониторинг водных экосистем Северного бассейна является неотъемлемым и важным компонентом биологического мониторинга, позволяющим провести комплексную диагностику функционирования экосистем.
Физиологические показатели не всегда могут быть использованы для биоиндикации состояния организмов ( Черкесова и др ., 2002).
В современных программах мониторинга для быстрой биоиндикации предлагают различные биохимические маркеры, позволяющие на клеточном уровне охарактеризовать величину ответной реакции организма на действие загрязняющих веществ, еще задолго до популяционного ответа: характеристики энергетического метаболизма (энергетический показатель, характеризующий отношение содержания жира к влаге), каротиноиды, на которые оказывает влияние гипоксия как результат антропогенного загрязнения, глутатион (компонент антиоксидантной системы), малоновый альдегид (показатель перекисного окисления мембранных липидов), ферменты, например, фермент холинэстераза, свободные радикалы. Также ярким показателем, используемым в современной биохимии, является аденилатный энергетический заряд (АЭЗ), выявляющий нарушение метаболизма ( Лукьянова , 2002; Лукьянова и др ., 2002; Мелехова и др. , 2002; Христофорова , 1989).
Целью данной работы явилось выявление биохимических особенностей тканей гидробионтов (рыб), наиболее характерных для прибрежной зоны Баренцева моря, в условиях антропогенного загрязнения.
Нами были проведены исследования сезонной динамики химического состава и биохимических свойств ряда промысловых рыб, характерных для вод Кольского залива и прибрежной зоны Баренцева моря на разных стадиях жизненного цикла (1998-2003). С позиций выявления биологических маркеров как показателей, отражающих степень ответной реакции организма на действие загрязняющих веществ, оценивалась активность тканевых протеолитических ферментов (по нарастанию аминного азота и по методу Л.Л. Константиновой (Константинова, Пахомова, 1970)) и содержание макроэргических соединений в мышечной ткани рыб. Результаты данных исследований сравнивались с результатами, полученными в лаборатории биохимии гидробионтов Полярного научно-исследовательского института морского рыбного хозяйства и океанографии им. Н.М. Книповича (ПИНРО) и с результатами предыдущих экспериментов, ранее проводимых в лабораториях биохимии (1990-1997).
Комплексная оценка всех этих направлений позволит выявить наиболее полную картину воздействия на состояние тканей гидробионтов, оценить их адаптационные возможности в условиях Крайнего Севера.
-
2. Объекты и методы исследования
Объектами биохимического исследования явились следующие промысловые гидробионты: – мойва Mallotus villosus villosus , род Mallotus , семейство корюшковых – OSMERIDAE, – пикша Melanogrammus aeglefinus , род Melanogrammus , семейство тресковых – GADIDAE, – треска Gadus morhua morhua , рода Gadus , семейство тресковых – GADIDAE,
– сайка (полярная тресочка) Boreogadus saida , род Boreogadus , семейство тресковых – GADIDAE,
– морская камбала Platessa platessa , род Platessa , семейство камбаловых – PLEURONECTIDAE,
– песчанка Ammodytes marinus , семейство песчанковых – AMMODYTIDAE.
Для каждого периода жизненного цикла выбранных гидробионтов определяли: содержание влаги, общего азота, белкового азота, небелкового азота, фракции водорастворимого белка, липидов, минеральных веществ, аминного азота, макроэргических соединений, активность тканевых протеолитических ферментов.
Определение массовой доли воды проводили высушиванием при 100-105°С. Метод основан на выделении (испарении) воды из гидробионта при тепловой переработке и определении изменения массы его взвешиванием.
Количественное определение водорастворимого белка и полипептидов проводилось фотоколориметрическим биуретовым макрометодом, основанным на биуретовой реакции, которая является показателем наличия пептидных связей в соединении и методом Лоури (основан на биуретовой реакции и на цветной реакции тирозиновых остатков белковой молекулы с реактивом Фолина (окисление тирозина фосфорномолибденовольфрамовокислым натрием)). Содержание общего азота определяли методом Къельдаля, включающим минерализацию биологического материала и термическую перегонку аммиака с водяным паром ( Северин , 1989; Овчинникова , 1999; 2000).
Определение содержания жира в мышечной ткани проводили методом Блая – Даера ( Северин , 1989).
Активность тканевых протеолитических ферментов определяли по нарастанию аминного азота методом формольного (формолового) титрования по Серенсену и по модифицированному методу Л.Л. Константиновой ( Константинова, Пахомова , 1970).
Для количественного определения макроэргических соединений в мышечной ткани использовали фотоколориметрический метод, основанный на цветной реакции с молибдатом аммония в кислой среде.
-
3. Исследования химического состава и биохимических свойств мойвы
Динамика химического состава и биохимических свойств мойвы в разные периоды годового цикла (осень 1998 г. – весна 2003 г.) представлена в табл. 1.
Таблица 1. Динамика химического состава мойвы в различные периоды годового цикла
|
Период годового цикла |
Влага, % |
Белок, % |
Жир, % |
Зола, % |
СО/В, % |
|
Осень 1998 г. (период нагула) |
70,5 ± 3,5 |
14,95 ± 6,52 |
14,1 ± 2,4 |
1,65 ± 0,09 |
30,7 |
|
Зима 1998 г. (зимовка) |
72,5 ± 8,9 |
16,0 ± 0,5 |
12,8 ± 4,1 |
1,0 ± 0,1 |
27,5 |
|
Март 1999 г. (преднерестовый период) |
80,6 ± 12,4 |
15,5 ± 2,9 |
4,8 ± 0,1 |
1,0 ± 0,1 |
19,4 |
|
Май-июнь 1999 г. (посленерестовый период) |
82,5 ± 7,9 |
14,0 ± 0,5 |
1,7 ± 0,1 |
1,1 ± 0,1 |
17,5 |
|
Осень 1999 г. (период нагула) |
68,6 ± 0,2 |
14,85 ± 0,05 |
13,9 ± 0,5 |
1,2 ± 0,1 |
31,4 |
|
Декабрь 1999 г. (зимовка) |
73,1 ± 4,5 |
16,1 ± 0,5 |
12,9 ± 0,7 |
1,1 ± 0,1 |
26,9 |
|
Март 2000 г. (преднерестовый период) |
81,4 ± 9,2 |
15,2 ± 0,1 |
5,2 ± 0,1 |
1,5 ± 0,1 |
18,6 |
|
Май 2000 г. (посленерестовый период) |
83,0 ± 7,5 |
13,9 ± 0,6 |
1,6 ± 0,1 |
1,32 ± 0,05 |
17,3 |
|
Осень 2000 г. (период нагула) |
74,3 ± 4,5 |
13,9 ± 0,1 |
14,0 ± 1,5 |
1,25 ± 0,07 |
31,5 |
|
Зима 2000 г. (зимовка) |
71,8 ± 6,7 |
16,0 ± 0,9 |
12,7 ± 1,8 |
1,15 ± 0,08 |
28,2 |
|
Март 2001 г. (преднерестовый период) |
81,0 ± 8,5 |
15,4 ± 1,3 |
5,1 ± 0,4 |
1,3 ± 0,1 |
19,1 |
|
Май 2001 г. (посленерестовый период) |
82,4 ± 6,5 |
13,0 ± 1,5 |
1,6 ± 0,1 |
1,1 ± 0,1 |
17,6 |
|
Осень 2001 г. (период нагула) |
75,4 ± 3,8 |
14,9 ± 1,2 |
13,9 ± 1,7 |
1,12 ± 0,08 |
32,1 |
|
Зима 2001 г. (зимовка) |
72,0 ± 4,5 |
16,2 ± 1,9 |
12,8 ± 1,4 |
1,18 ± 0,04 |
28,1 |
|
Февраль-март 2002 г. (преднерестовый период) |
80,7 ± 5,8 |
15,3 ± 0,1 |
4,9 ± 0,5 |
1,17 ± 0,07 |
19,3 |
|
Май 2002 г. (посленерестовый период) |
84,0 ± 9,5 |
15,4 ± 0,3 |
1,58 ± 0,07 |
1,16 ± 0,04 |
16,0 |
|
Осень 2002 г. (период нагула) |
67,5 ± 4,3 |
14,8 ± 0,9 |
14,2 ± 0,5 |
1,2 ± 0,1 |
32,5 |
|
Зима 2002 г. (зимовка) |
72,6 ± 2,7 |
15,9 ± 1,4 |
12,8 ± 0,9 |
1,2 ± 0,1 |
28,3 |
|
Весна 2003 г. (преднерестовый период) |
81,5 ± 8,5 |
11,5 ± 0,7 |
5,3 ± 0,4 |
1,15 ± 0,06 |
17,1 |
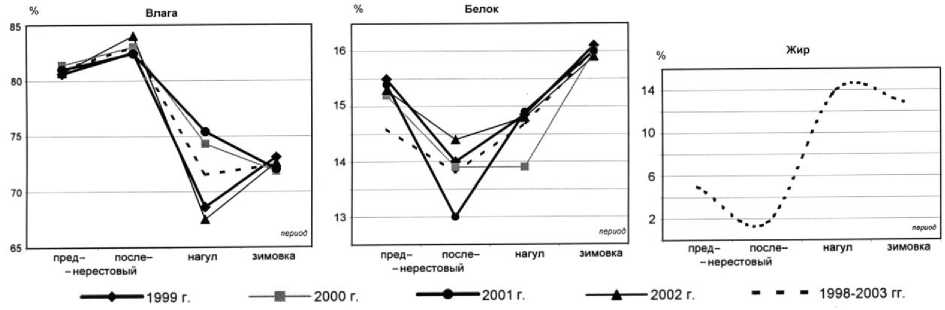
Рис. 1. Сезонная динамика содержания влаги, белка и жира в мышечной ткани мойвы
Изучена сезонная динамика влагосодержания (1998-2003). Установлено, что наименьшее содержание влаги в мышечной ткани мойвы наблюдается в конце периода нагула (осенний период – октябрь, с 1998 по 2003 гг.) – 67,5-75,4 %, составляя в среднем 71,5 ± 4,0 %. При этом минимальные величины в данный период отмечены в 2002 г., а максимальные – в 2001 г.
Количественное содержание влаги в мышечной ткани мойвы в преднерестовый, нерестовый и период зимовки на протяжении 1998-2003 гг. было сходным.
В период зимовки содержание влаги за период с 1998 по 2003 гг. составляет 72,4 ± 0,6 %. В преднерестовый период (февраль-март, с 1998 по 2003 гг.) содержание влаги возрастает до 80,6-81,4 %, составляя в среднем 81,0 ± 0,5 %. Наибольшее содержание влаги наблюдается в посленерестовый период (май-июнь) – 82,4-84 % со средним значением 83,0 ± 0,5 %. При этом минимальные величины в данный период отмечены в 1999 г., а максимальные – в 2002 г.
Среднегодовое содержание влаги за период 1998-2003 гг. составило 76,9 %. При этом наиболее низкие значения отмечены для 1999 и 2002 гг. (76,2 %), а максимальные для 2000 и 2001 гг. (до 77,7 %).
При сравнении полученных нами данных по влагосодержанию с данными ПИНРО до 1998 г. установлено, что в период с 1998 по 2003 гг. наблюдается незначительное увеличение содержания влаги по сравнению с предыдущим периодом.
Проведен анализ сезонной динамики содержания общего азота, белкового азота, небелкового азота в мышечной ткани мойвы (1998-2003).
Наименьшее содержание белка отмечено в посленерестовый период (май-июнь), составляя в среднем 13,8 ± 0,6 %, наиболее высокое содержание белка характерно для зимнего периода (декабрь) – 16,2 ± 0,1%. Среднегодовое содержание белка в мышечной ткани мойвы за период 1998-2003 гг. составило 14,96 %.
■ Данные ПИНРО (до 1998 г.)
□ Собств. данные (1998-2003 гг.)
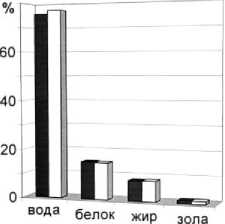
Рис. 2. Среднегодовое содержание основных химических показателей мышечной ткани мойвы
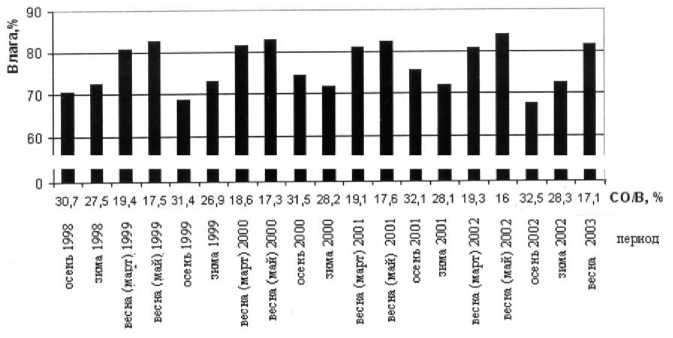
Рис. 3. Соотношение массовой доли воды и СО/В
При сравнении полученных нами данных по содержанию белка за период с 1998 по 2003 гг. с данными ПИНРО до 1998 г. отмечено увеличение содержания белка в период зимовки и уменьшение (в среднем на 2 %) в преднерестовый, посленерестовый периоды и в период нагула. Характер сезонной динамики содержания белка в мышечной ткани сходен.
Проанализирована сезонная динамика суммарного содержания липидов для периода 1998-2003 гг.
Интенсивность питания мойвы самая высокая в летние месяцы, откорм заканчивается в октябре, а в ноябре-январе рыба практически не питается. В период нагула массовый состав мойвы характеризуется наибольшим количеством полостного жира ( Лебская и др. , 1993; Лебская и др. , 1998).
В конце периода нагула (осень) наблюдается наиболее высокое содержание липидов - 13,914,2 %, составляя в среднем 14,0 ± 0,1 %; к периоду зимовки (декабрь) содержание липидов снижается и составляет в среднем 12,8 ± 0,1 %. В преднерестовый период (февраль) содержание жира в мышечной ткани резко снижается до 4,8-5,3 %, составляя в среднем 5,1 ± 0,2 %. Это обусловлено расходованием жировых запасов в процессе гаметогенеза ( Шульман , 1969). Самое низкое содержание липидов характерно для нерестового и посленерестового периода (май-июнь), составляя в среднем 1,62 ± 0,05 %.
Характер сезонной динамики содержания липидов и их количественное распределение за период 1998-2003 гг. сходны.
Среднегодовое содержание липидов за период 1998-2003 гг. составило 8,34 %. При сравнении полученных нами данных по содержанию липидов за 1998-2003 гг. с данными ПИНРО до 1998 г. установлено, что наблюдается незначительное увеличение среднегодового содержания жира, при этом отмечено повышение доли липидной фракции в преднерестовый период (в среднем на 16 %) и уменьшение в период нагула и посленерестовый период (на 12 %). Это может свидетельствовать о том, что в исследованный период мобилизация жировых запасов была выше по сравнению с предыдущим периодом.
В целом сезонная динамика содержания влаги и липидов соотносится с данными ПИНРО ( Лебская и др. , 1993; Лебская и др. , 1998).
По литературным данным, групповой состав липидов чрезвычайно лабилен и представлен в основном триглицеридами, на долю которых может приходиться от 51,0 до 87,5 %. Содержание фосфолипидов составляет 22,4-36,0 %, холестерина - 1,2-4,2 % от массы всех липидов. Жирнокислотный состав липидов мойвы представлен 24 жирными кислотами с числом углеродных атомов от 12 до 22. Наибольшее число двойных связей равно 6. Основную массу жирных кислот составляют мононенасыщенные (до 44,8 %), на втором месте по суммарному количеству находятся полиненасыщенные (до 41,5 %), на третьем - насыщенные (до 22,6 %) ( Костылев, Рябошапко , 1982; Лебская и др. , 1998).
Проанализирована динамика содержания минеральных веществ в мышечной ткани мойвы. Содержание золы по нашим данным колеблется в диапазоне 1,0 -1,65 %, составляя в среднем 1,19 %; зависимости от стадии жизненного цикла не наблюдается. По данным ПИНРО содержание минеральных веществ колеблется от 1,1 до 1,3 % (в среднем 1,2 %).
Показателем, удовлетворительно отражающим состояние организма рыбы, является относительное содержание влаги в теле рыбы. В то же время у многих рыб степень оводненности тканей тесно связана с относительным содержанием белка и жира - основными энергетическими характеристиками организма. У баренцевоморской мойвы между содержанием жира и влаги существует четкая обратная связь, что позволяет рассматривать влажность как интегрирующий (зависящий от содержания жира и белка) показатель, характеризующий энергетическое состояние мойвы. Двинин Ю.Ф. предлагает для оценки состояния организма рыбы такой характерный энергетический показатель как отношение сухого остатка к влаге (СО/В) (Двинин и др., 1992). Данный показатель реагирует на изменение в балансе между водой и запасными питательными веществами. Увеличение СО/В связано либо с накоплением жира и белка, либо с уменьшением оводненности тканей, что служит признаком благополучного состояния рыбы. Падение энергетического показателя, соответственно, свидетельствует об истощении рыбы (Двинин и др., 1992).
Нами проведен расчет СО/В для разных периодов жизненного цикла мойвы. Анализ СО/В показывает уменьшение СО/В от осеннего к весеннему периоду. Увеличение СО/В наступает в период нагула (рис. 3).
Проанализирована динамика содержания аминного азота в мышечной ткани мойвы (табл. 2).
Белок мяса мойвы полноценный, содержит все незаменимые аминокислоты, лимитирующих аминокислот нет. По количественному содержанию преобладают глутаминовая кислота, лизин и лейцин. Накопление свободных аминокислот в мышечной ткани подвержено значительным колебаниям, зависит от сезона, пола, возраста и, возможно, от экологических факторов. Диапазон их содержания велик – от 56,1 до 212,9 мг % ( Лебская и др. , 1998).
Содержание исходного аминного азота возрастает в осенний период, более низкое содержание характерно для преднерестового периода и нереста – 35-40 мг %. Наиболее высокое содержание аминного азота отмечено для периода нагула – 50-51 мг %. Данные по N ам для периода весна 1999 г. – весна 2003 г. представлены в табл. 2.
По содержанию аминного азота можно судить об интенсивности гидролиза белков и активности протеолитических ферментов.
Проанализирована относительная активность тканевых протеолитических ферментов в мышечной ткани мойвы (методом формольного титрования) для времени гидролиза 30 мин и 1 час. С увеличением времени гидролиза относительная активность тканевых протеолитических ферментов возрастает.
Наиболее высокая активность ферментов отмечена весной 2000 г. и осенью 2001 г., наиболее низкая – осенью 2002 г. (рис. 4).
Проанализирована динамика содержания макроэргических соединений (аденозинтрифосфорной кислоты) в мышечной ткани мойвы. В табл. 3 представлены средние значения содержания макроэргов в зависимости от периода жизненного годового цикла (1998-2003).
Как видно из представленных данных, наиболее высокое содержание макроэргических соединений характерно для преднерестового периода и нереста. Наименьшее содержание макроэргов наблюдается в посленерестовый период (истощенное состояние рыбы), в период нагула имеет место увеличение количества аденозинтрифосфорной кислоты по сравнению с посленерестовым промежутком времени.
|
Весна 2003 г. [ Таблица 2. Динамика содержания аминного азота _ Осень 2002 г. 1 в мышечной ткани мойвы ____________________________________________________________________________________ Весна 2002 г. 1 |
__________________I |
||||||
|
Период годового цикла |
N ам. исх , мг % |
N ам , мг % , t гидр = 30 мин |
N ам , мг % , t гидр = 1 час |
Осень 2001 г. |_____ Весна 2000 г. [ _________________________ |
1 |
||
|
Весна 1999 г. |
45,0 ± 3,0 |
180 |
890 |
Осень 1999 г. |___________________ | |
|||
|
Осень 1999 г. |
50,0 ± 2,5 |
175 |
910 |
Весна 1999 г. |____________________________| |
|||
|
Весна 2000 г. |
46,0 ± 6,3 |
140 |
1700 |
0 10 20 30 Мам, t гидр 60 мин / Мам исх Рис. 4. Соотношение аминного азота в мышечной ткани мойвы до и после гидролиза |
|||
|
Осень 2001 г. |
51,0 ± 3,5 |
125 |
1760 |
||||
|
Весна 2002 г. |
35,0 ± 2,5 |
120 |
607,6 |
||||
|
Осень 2002 г. |
50,7 ± 2,7 |
130 |
650 |
||||
|
Весна 2003 г. |
37,0 ± 3,5 |
120 |
607,6 |
||||
Таблица 3. Динамика изменения содержания макроэргических соединений в мышечной ткани мойвы (1998-2002)
|
Период |
Содержание макроэргических соединений, моль/г мышечной ткани |
|
Осень (период нагула) |
3 X 10-5 |
|
Весна (преднерестовый период) |
10- 4 |
|
Май-июнь (посленерестовый период) |
10- 6 |
-
4. Исследования химического состава и биохимических свойств пикши
Изучена динамика химического состава и биохимических свойств пикши в разные периоды годового цикла (осень 1998 г. – весна 2003 г.). Полученные данные по содержанию влаги, белка, жира, золы в мышечной ткани пикши представлены в табл. 4.
Проанализирована сезонная динамика содержания воды в мышечной ткани пикши для разных периодов годового жизненного цикла.
Таблица 4. Динамика химического состава пикши в разные периоды годового цикла
|
Период годового цикла |
Влага, % |
Белок, % |
Жир, % |
Зола, % |
|
Осень 1998 г. (посленерестовый период) |
80,1 ± 2,6 |
16,4 ± 1,7 |
0,11 ± 0,06 |
1,1 ± 0,2 |
|
Зима 1998 г. (перед нагулом) |
71,0 ± 1,5 |
18,0 ± 2,0 |
0,12 ± 0,02 |
1,2 ± 0,1 |
|
Весна, март 1999 г. (преднерестовый период) |
76,0 ± 4,5 |
18,5 ± 2,0 |
0,13 ± 0,02 |
1,3 ± 0,1 |
|
Май-июнь 1999 г. (нерест) |
73,0 ± 2,5 |
18,3 ± 2,2 |
0,14 ± 0,02 |
1,2 ± 0,3 |
|
Осень 1999 г. (посленерестовый период) |
81,0 ± 4,0 |
16,3 ± 0,3 |
0,10 ± 0,05 |
1,15 ± 0,08 |
|
Зима 1999 г. (перед нагулом) |
73,0 ± 5,0 |
18,1 ± 0,5 |
0,10 ± 0,02 |
1,25 ± 0,15 |
|
Весна, март 2000 г. (преднерестовый период) |
76,1 ± 4,9 |
18,2 ± 0,3 |
0,14 ± 0,04 |
1,30 ± 0,15 |
|
Весна, апрель-май 2000 г. (нерест) |
77,5 ± 5,5 |
18,6 ± 0,2 |
0,145 ± 0,023 |
1,1 ± 0,1 |
|
Осень 2000 г. (посленерестовый период) |
80,3 ± 6,9 |
16,2 ± 0,2 |
0,10 ± 0,01 |
1,1 ± 0,1 |
|
Зима 2000 г. (перед нагулом) |
73,0 ± 3,5 |
18,2 ± 0,2 |
0,13 ± 0,04 |
1,2 ± 0,1 |
|
Весна, март 2001 г. (преднерестовый период) |
76,7 ± 3,7 |
18,7 ± 0,2 |
0,133 ± 0,042 |
1,3 ± 0,2 |
|
Весна, май 2001 г. (нерест) |
76,9 ± 4,9 |
18,0 ± 0,5 |
0,138 ± 0,036 |
1,20 ± 0,15 |
|
Осень 2001 г. (посленерестовый период) |
80,5 ± 2,1 |
16,6 ± 0,3 |
0,11 ± 0,01 |
1,15 ± 0,08 |
|
Зима 2001 г. (перед нагулом) |
72,8 ± 1,4 |
18,0 ± 0,5 |
0,14 ± 0,04 |
1,25 ± 0,15 |
|
Весна, март 2002 г. (преднерестовый период) |
76,1 ± 4,9 |
18,5 ± 0,4 |
0,142 ± 0,058 |
1,3 ± 0,2 |
|
Весна, май 2002 г. (нерест) |
75,0 ± 4,5 |
18,2 ± 0,6 |
0,14 ± 0,04 |
1,20 ± 0,15 |
|
Осень 2002 г. (посленерестовый период) |
81,2 ± 3,9 |
16,3 ± 0,7 |
0,09 ± 0,01 |
1,1 ± 0,1 |
|
Зима 2002 г. (перед нагулом) |
72,0 ± 3,0 |
18,1 ± 0,4 |
0,15 ± 0,03 |
1,20 ± 0,15 |
|
Весна, март 2003 г. (преднерестовый период) |
76,2 ± 2,9 |
18,8 ± 0,7 |
0,14 ± 0,04 |
1,25 ± 0,22 |
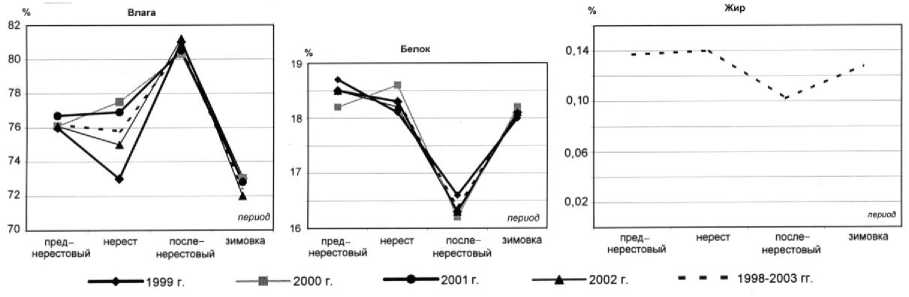
Рис. 5. Сезонная динамика содержания влаги, белка и жира в мышечной ткани пикши
Наиболее высокое содержание влаги характерно для посленерестового периода (осень) – 8081,2 %, в среднем составляя 80,6 ± 0,5 %. Наименьшее содержание отмечено в зимний период – 71-73 %, составляя в среднем 72,4 ± 0,9 %. В преднерестовый период содержание влаги составило в среднем 76,2 ± 0,3 %, в нерестовый – 75,6 ± 2,0 %.
Характер сезонной динамики содержания влаги за период 1998-2003 гг. сходен.
Наибольшее содержание влаги отмечено в 2000-2001 гг. (до 76,7 %), наименьшее в 1999 г. (75,8 %).
Среднегодовое содержание влаги в мышечной ткани пикши за 1998-2003 гг. составило 76,3 %.
При сравнении полученных результатов с данными ПИНРО отмечено снижение среднегодового содержания влаги на 5 %, а в зимний период – на 11 %.
Исследована сезонная динамика содержания общего азота, белкового азота, небелкового азота в мышечной ткани пикши. Наибольшее содержание белковой фракции в мышечной ткани пикши наблюдается перед нерестом – 18,2-18,8 % (в среднем 18,5 ± 0,2 %) и в период нереста – 18,3 ± 0,2 %, наименьшее – после нереста (осень) – 16,2-16,6 %, в среднем составляя 16,4 ± 0,2 %. Зимой содержание белка увеличивается до 18,0-18,2 %, в среднем 18,1 ± 0,1 %.
Белок мышечной ткани содержит все незаменимые аминокислоты. Лимитирующих аминокислот нет. В наибольшем количестве содержатся глутаминовая и аспарагиновая кислоты, лизин, лейцин ( Лебская и др. , 1993; Лебская и др. , 1998).
Среднегодовое содержание белка в мышечной ткани пикши за 1998-2003 гг. составило 17,8 %.
Характер сезонной динамики содержания белка и его количественное распределение за период 1998-2003 гг. сходен.
При сравнении полученных результатов с данными ПИНРО отмечено снижение содержания белка в посленерестовый период на 10 % и увеличение в зимний период на 7 %.
Изучена сезонная динамика содержания жира в мышечной ткани пикши также в зависимости от периода годового цикла. Жирность мышечной ткани пикши редко бывает выше 0,1-0,5 % ( Лебская и др ., 1993; Лебская и др. , 1998). Наибольшая массовая доля липидов отмечена в период нереста (весна, апрель-май) и в преднерестовый период (весна, март) - 0,13-0,15 %, составляя в среднем 0,14 ± 0,04 %. Наименьшая массовая доля липидной фракции характерна для посленерестового периода – 0,09-0,11 %, составляя в среднем 0,10 ± 0,01 %.
В зимний период перед нагулом среднее содержание липидов составило 0,13 ± 0,02 %.
Среднегодовое содержание липидов в мышечной ткани пикши за 1998-2003 гг. составило 0,13 %.
Характер сезонной динамики содержания жира и его количественное распределение за период 1998-2003 гг. сходен.
При сравнении полученных результатов за период 1998-2003 гг. с данными ПИНРО до 1998 г. отмечено снижение содержания липидов в среднем до 40 %.
Основную долю липидов мышечной ткани составляют фосфолипиды, холестерин и свободные жирные кислоты. В групповом составе липидов печени преобладают триглицериды, у рыбы, выловленной осенью, их доля составляет 65-66 % ( Лебская и др. , 1993; Лебская и др. , 1998).
Проанализирована динамика содержания минеральных веществ в мышечной ткани пикши. Содержание золы, по нашим данным, составляет 1,1-1,3 % (в среднем 1,20 ± 0,05 %). Зависимости от стадии жизненного цикла не наблюдается. По данным ПИНРО, содержание минеральных веществ колеблется от 1,14 до 1,47 %, составляя в среднем 1,3 %.
Проанализирована динамика активности тканевых протеолитических ферментов мышечной ткани пикши методом формольного титрования и по методу Л.Л. Константиновой (метод Ансона в модификации И.С. Петровой). Данные, полученные методом формольного титрования, представлены в табл. 5, результаты определения методом Л.Л. Константиновой – в табл. 6.
Как видно из представленных данных, наиболее высокая активность тканевых протеолитических ферментов мышечной ткани пикши характерна для преднерестового периода и нереста, наименьшая – для посленерестового периода. Активность тканевых протеолитических ферментов мышечной ткани пикши в 2003 г. несколько выше по сравнению с предыдущими годами, что можно объяснить понижением уровня антропогенного загрязнения вод Кольского залива и прибрежной зоны Баренцева моря.
Проанализировано содержание аминного азота в мышечной ткани исходных образцов пикши. Полученные данные представлены в табл. 7.
Таблица 5. Динамика относительной активности тканевых протеолитических ферментов мышечной ткани пикши (метод формольного титрования)
|
Период жизненного цикла |
N ам . , мг % (1997) |
N ам . , мг % (2003) |
|
Осень (после нереста) |
271,0 ± 28,5 |
400,0 ± 20,5 |
|
Зима (нагул) |
555,3 ± 32,8 |
650,5 ± 18,0 |
|
Весна, март-апрель (нерест) |
686,2 ± 34,2 |
710,3 ± 17,2 |
Таблица 6. Динамика активности тканевых протеолитических ферментов мышечной ткани пикши (метод Л.Л. Константиновой)
|
Период жизненного цикла |
Активность, мкмоль тирозина/г (1998-1999) |
Активность, мкмоль тирозина/г (2002-2003) |
|
Осень |
0,016 ± 0,002 |
0,025 ± 0,003 |
|
Зима |
0,030 ± 0,003 |
0,040 ± 0,003 |
|
Весна |
0,040 ± 0,003 |
0,050 ± 0,005 |
Таблица 7. Содержание аминного азота в пикше (2002)
|
Время |
N ам. исх . , мг % |
N ам . , мг % t гидр = 0,5 ч |
N ам . , мг % t гидр = 1ч |
|
Апрель |
39,4 ± 5,1 |
140,4 ± 9,6 |
275,5 ± 11,8 |
|
Сентябрь |
47,0 ± 4,4 |
130,2 ± 6,7 |
210 ± 12 |
|
Ноябрь |
1680 ± 50 |
1700 ± 14 |
1750 ± 44 |
Таблица 8. Содержание макроэргических соединений в мышечной ткани пикши в зависимости от периода жизненного цикла
|
Период годового цикла |
моль/г |
|
Нерест |
3 х 10-5 |
|
Преднерестовый |
10-4 |
|
Посленерестовый |
10-6 |
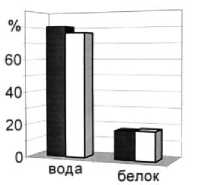
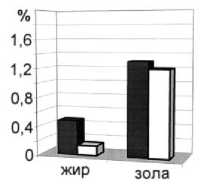
Рис. 6. Среднегодовое содержание основных химических показателей мышечной ткани пикши
-
■ Данные ПИНРО (до 1998 г.)
-
□ Собственные данные (1998-2003 гг.)
-
5. Исследования химического состава и биохимических свойств трески и сайки
Была исследована динамика изменения химического состава и биохимических свойств трески в зависимости от стадии жизненного цикла (1998-2003).
Проведен анализ динамики содержания макроэргических соединений (аденозинтрифосфорной кислоты) в мышечной ткани пикши. Определение макроэргов проводилось фотоколориметрическим методом. В табл. 8 представлены средние значения результатов по содержанию макроэргов в зависимости от периода жизненного годового цикла (1998-2003).
Как видно из представленных данных, наиболее высокое содержание макроэргических соединений характерно для преднерестового периода и нереста. Наименьшее содержание макроэргов наблюдается для посленерестового периода (истощенное состояние рыбы). В период нагула имеет место увеличение количества аденозинтрифосфорной кислоты по сравнению с посленерестовым промежутком времени.
Ряд объектов был выловлен в ноябре 2002 г. возле пос. Чан-Ручьи (южная сторона залива). Биохимия данных образцов может охарактеризовать антропогенное воздействие на Кольский залив.
Полученные данные по содержанию влаги, белка, жира, золы в мышечной ткани трески представлены в табл. 9.
Проанализирована сезонная динамика содержания воды в мышечной ткани трески для разных периодов годового жизненного цикла.
Наиболее высокое содержание влаги характерно для посленерестового периода (осень) – 80,482 % (среднее 81,3 ± 0,6 %), что обусловлено нерестовым истощением и менее благоприятными условиями питания в конце зимы; наименьшее содержание отмечено в преднерестовый период – в среднем 73,2 ± 0,8 %, с увеличением в нерестовый период до 76,9 ± 1,8 %. Для зимнего периода содержание влаги составило – 73-77 % (среднее 75 %).
Характер сезонной динамики содержания влаги за период 1998-2003 гг. не менялся.
Среднегодовое содержание влаги в мышечной ткани трески за 1998-2003 гг. составило 76,5 %.
При сравнении полученных результатов с данными ПИНРО (до 1998 г.) отмечено снижение содержания влаги в исследованный нами период на 5 %.
Таблица 9. Динамика химического состава мышечной ткани трески
|
Период годового цикла |
Влага, % |
Белок, % |
Жир, % |
Зола, % |
|
Осень 1998 (посленерестовый период) |
82,0 ± 3,5 |
16,5 ± 0,5 |
0,61 ± 0,08 |
0,85 ± 0,01 |
|
Зима 1998 (зимовка) |
74,0 ± 4,0 |
16,8 ± 0,3 |
0,85 ± 0,08 |
1,0 ± 0,2 |
|
Весна 1999 (преднерестовый период) |
74,2 ± 6,3 |
16,9 ± 0,5 |
0,92 ± 0,09 |
1,0 ± 0,1 |
|
Весна 1999 (нерест) |
79,6 ± 2,8 |
15,8 ± 0,7 |
0,65 ± 0,07 |
1,15 ± 0,08 |
|
Осень 1999 (посленерестовый период) |
80,4 ± 7,4 |
16,3 ± 0,3 |
0,71 ± 0,03 |
1,16 ± 0,05 |
|
Зима 1999 (зимовка) |
73,1 ± 5,4 |
16,45 ± 0,18 |
0,87 ± 0,07 |
1,13 ± 0,01 |
|
Весна 2000 (преднерестовый период) |
72,3 ± 4,5 |
16,85 ± 0,35 |
0,91 ± 0,06 |
1,14 ± 0,03 |
|
Весна 2000 (нерест) |
75,6 ± 4,6 |
15,75 ± 0,50 |
0,68 ± 0,07 |
1,15 ± 0,08 |
|
Осень 2000 (посленерестовый период) |
81,5 ± 6,3 |
16,0 ± 0,5 |
0,65 ± 0,06 |
1,10 ± 0,11 |
|
Зима 2000 (зимовка) |
76,0 ± 7,5 |
16,45 ± 0,52 |
0,8 ± 0,1 |
1,12 ± 0,06 |
|
Весна 2001 (преднерестовый период) |
73,2 ± 8,3 |
17,2 ± 0,5 |
0,89 ± 0,09 |
1,0 ± 0,1 |
|
Весна 2001 (нерест) |
76,5 ± 6,3 |
16,1 ± 0,4 |
0,65 ± 0,05 |
1,12 ± 0,06 |
|
Осень 2001 (посленерестовый период) |
81,5 ± 6,0 |
16,4 ± 0,5 |
0,75 ± 0,08 |
1,15 ± 0,05 |
|
Зима 2001 (зимовка) |
77,0 ± 7,5 |
16,5 ± 0,4 |
0,88 ± 0,09 |
1,11 ± 0,14 |
|
Весна 2002 (преднерестовый период) |
73,0 ± 7,0 |
16,8 ± 0,7 |
0,93 ± 0,05 |
1,12 ± 0,07 |
|
Весна 2002 (нерест) |
76,0 ± 7,0 |
15,7 ± 0,8 |
0,69 ± 0,05 |
1,10 ± 0,08 |
|
Осень 2002 (посленерестовый период) |
81,3 ± 5,7 |
16,3 ± 0,4 |
0,65 ± 0,07 |
0,90 ± 0,12 |
|
Зима 2002 (зимовка) |
73,0 ± 6,5 |
16,4 ± 0,5 |
0,86 ± 0,07 |
1,12 ± 0,11 |
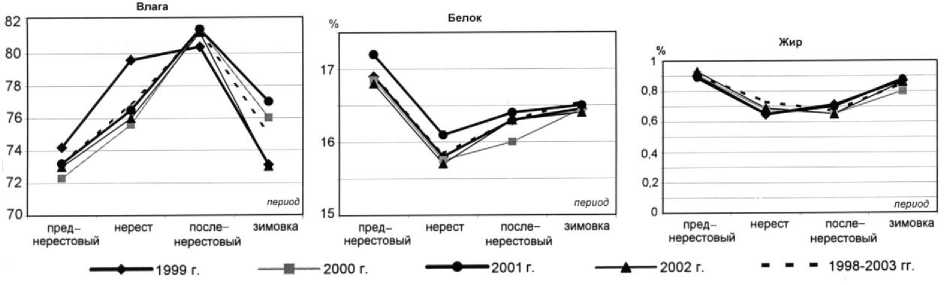
Рис. 7. Сезонная динамика содержания влаги, белка и жира в мышечной ткани трески
Исследована сезонная динамика содержания общего азота, белкового азота, небелкового азота в мышечной ткани трески. Белок мышечной ткани трески содержит все незаменимые аминокислоты, их сумма составляет 43,6 % от общего количества аминокислот. В наибольших количествах присутствуют аспарагиновая и глутаминовая кислоты, лизин. Лимитирующих аминокислот нет. Характер сезонной динамики содержания белка и его количественное распределение за период 1998-2003 гг. не менялись. Наибольшее содержание белка в мышечной ткани трески наблюдается перед нерестом – 16,9 ± 0,2 %, наименьшее – в период нереста (март) – 15,8 ± 0,2 %. В посленерестовый период (осень) содержание белка увеличилось до 16,3 ± 0,2 %. В зимний период содержание белка составило в среднем 16,5 ± 0,1 %.
Белок мышечной ткани полноценный, лимитирующих аминокислот нет. В аминокислотном составе белка мышечной ткани количественно преобладают глутаминовая кислота, лизин, аспарагиновая кислота и лейцин ( Лебская и др ., 1993; Лебская и др. , 1998).
Среднегодовое содержание белка в мышечной ткани трески за период 1998-2003 гг. составило 16,4 %. При сравнении полученных результатов с данными ПИНРО (до 1998 г.) отмечено снижение содержания белка в исследованный нами период на 7 %.
Исследована сезонная динамика содержания жира в мышечной ткани трески также в зависимости от периода годового цикла.
Согласно литературным данным ( Лебская и др ., 1993; Лебская и др. , 1998), в липидах мышечной ткани преобладают фосфолипиды (81 %), на долю триглицеридов приходится 3,7 %, холестерина – 5,5 %, эфиров стеринов – 3,9 %, свободных жирных кислот – 4,6 %; в печени преобладают триглицериды (8692 %).
Характер сезонной динамики содержания жира и его количественное распределение за период 1998-2003 гг. не менялись. Наибольшая массовая доля липидов отмечена в преднерестовый период (весна, март) – 0,9-0,93 %, в среднем составляя 0,91 ± 0,02 %, снижаясь в период нереста (весна, апрель-май) до 0,67 ± 0,02 %. Наименьшая массовая доля липидов характерна для посленерестового периода – 0,66 ± 0,04 %. В зимний период содержание липидов составило 0,8-0,88 % (среднее 0,85 ± 0,03 %).
Среднегодовое содержание жира в мышечной ткани трески за 1998-2003 гг. составило 0,78 %.
Определено содержание аминного азота в мышечной ткани трески (табл. 10).
Оценена относительная активность тканевых протеолитических ферментов в мышечной ткани трески методом формольного титрования для времени гидролиза 30 мин и 1 час. Установлено, что с увеличением времени гидролиза относительная активность тканевых протеолитических ферментов возрастает.
Наиболее высокая активность ферментов отмечена в мае 1999 г., наиболее низкая – осенью 1999 г.
Проанализирована динамика содержания минеральных веществ в мышечной ткани трески. Содержание золы, по нашим данным за исследованный период (1998-2003), составляет 0,85-1,16 % (в среднем 1,1 ± 0,1 %). Зависимости от стадии жизненного цикла не наблюдается. По данным ПИНРО (до 1998 г.), содержание минеральных веществ колеблется от 0,79 до 1,55 %, составляя в среднем 1,2 %.
Сайка является мелкой пелагической рыбой. Химический состав мяса сайки отличается от состава мяса тресковых меньшим содержанием белка (составляет 15,6 % против 17,5-18,0 %).
Таблица 10. Динамика содержания аминного азота в мышечной ткани трески (1999)
|
Период годового цикла |
N ам исх . , мг% |
N ам . , мг% t гидр = 0,5 ч |
N ам . , мг% t гидр = 1ч |
|
Апрель (нерест) |
40,0 ± 4,5 |
180,0 ± 13,0 |
350,0 ± 20,0 |
|
Май (посленерестовый) |
25,3 ± 4,6 |
140,5 ± 10,0 |
295,2 ± 26,0 |
|
Осень (посленерестовый) |
60,0 ± 3,5 |
180,4 ± 13,8 |
320 ± 24 |
Образцы сайки были выловлены в ноябре 2002 г. в п. Чан-Ручьи. Данный период является преднерестовым периодом (для которого характерна минимальная интенсивность питания). Мышечная ткань сайки в преднерестовый период характеризуется повышением содержания белка и жира. Получены следующие результаты: содержание влаги – 86,0 ± 2,5 %, белка – 15,6 ± 0,2 %, жира – 1,64 ± 0,09 %, золы – 1,2 ± 0,2 %, аминного азота – 60 мг %, водорастворимого белка – 1,3 г/л вытяжки. Результаты близки к данным, полученным в ПИНРО. Белок сайки полноценный, лимитирующих аминокислот нет. Жир сайки обладает сравнительно высокой непредельностью (йодное число в среднем составляет 132 % йода), что обусловливает его биологическую ценность и неустойчивость к окислению ( Лебская и др ., 1993; Лебская и др. , 1998).
Полученные результаты сравнивали с данными ПИНРО (до 1998 г.). В наших данных наблюдается увеличение содержания влаги в среднем на 6 %, золы – на 14 % и незначительное снижение содержания белка.
-
6. Исследования химического состава и биохимических свойств камбалы
Проводились исследования химического состава камбалы. Результаты представлены в табл. 11.
Проанализирована сезонная динамика содержания воды в мышечной ткани камбалы для разных периодов годового жизненного цикла.
Наиболее высокое содержание влаги характерно для посленерестового периода (осень) – в среднем 81,3 ± 3,7 %, наименьшее содержание отмечено в нерестовый период – в среднем 75,0 ± 7,5 %.
При сравнении полученных нами результатов (1998-2002) с данными ПИНРО (до 1998 г.) отмечено снижение содержания влаги в исследованный нами период на 2 %.
Исследована сезонная динамика содержания жира в мышечной ткани камбалы также в зависимости от периода годового цикла. Наибольшая массовая доля липидов отмечена в преднерестовый период (весна, март) – в среднем 7,6 ± 0,5 %, снижаясь в период нереста (весна, апрель-май) до 5,4 ± 0,5 %. Наименьшая массовая доля липидов характерна для посленерестового периода – 3,2 ± 0,3 %.
При сравнении полученных нами результатов (1998-2002) с данными ПИНРО (до 1998 г.) отмечено увеличение содержания жира в исследованный нами период в среднем на 40 %.
Исследована сезонная динамика общего белка в мышечной ткани камбалы. Мышечная ткань морской камбалы по содержанию белка беднее трески. Обводненность белков мяса камбалы высокая, несколько выше, чем у трески ( Лебская и др ., 1993; Лебская и др. , 1998).
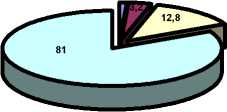
Наибольшее содержание белка в мышечной ткани камбалы наблюдается перед нерестом – 14,5 ± 0,6 %, снижаясь в период нереста до 14,0 ± 0,9 % и достигая наименьших значений в посленерестовый период – 12,8 ± 0,7 %.

Рис. 10. Среднегодовое содержание основных химических показателей мышечной ткани камбалы
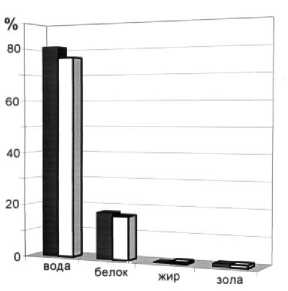
Рис. 8. Среднегодовое содержание основных химических показателей мышечной ткани трески
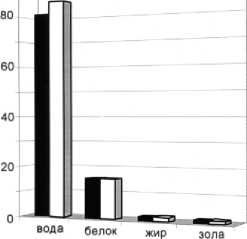
Рис. 9. Среднегодовое содержание основных химических показателей мышечной ткани сайки в преднерестовый период
Таблица 11. Динамика химического состава мышечной ткани морской камбалы
|
Период годового цикла |
Влага, % |
Белок, % |
Жир, % |
Зола, % |
|
Осень 1998 г. (посленерестовый период) |
81,1 ± 3,7 |
12,8 ± 0,7 |
3,2 ± 0,3 |
1,21 ± 0,04 |
|
Зима 2000 г. (преднерестовый период) |
78,2 ± 6,0 |
14,5 ± 0,6 |
7,6 ± 0,5 |
1,25 ± 0,13 |
|
Весна 2002 г. (нерест) |
75,2 ± 7,3 |
14,0 ± 0,5 |
5,4 ± 0,6 |
1,23 ± 0,09 |
При сравнении полученных нами результатов с данными ПИНРО (до 1998 г.) отмечено снижение содержания белка в исследованный нами период (1998-2002) на 8 %.
Мясо морской камбалы по содержанию белка (в среднем 14,9 %) беднее мяса трески. Жирность его также невысока (в среднем 3,3 %) и колеблется в зависимости от годового цикла ( Лебская и др. , 1993, 1998).
Обводненность белков мяса довольно высокая, несколько выше, чем у трески. Белок мышечной ткани полноценный, лимитирующих аминокислот нет.
Проанализирована динамика содержания минеральных веществ в мышечной ткани камбалы. Содержание золы, по нашим данным за исследованный период (1998-2003), составило 1,23 ± 0,02 %. Зависимости от стадии жизненного цикла не наблюдается. По данным ПИНРО содержание минеральных веществ в период до 1998 г. колеблется от 1,1 до 1,3 %, составляя в среднем 1,2 %.
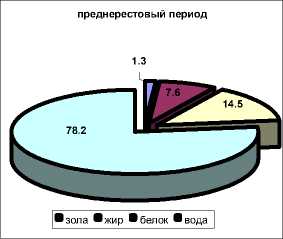
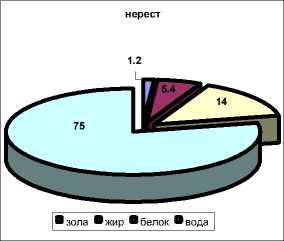
посленерестовый период
1,2
я зола ■ жир ■ белок ■ вода
Рис. 11. Химический состав мышечной ткани камбалы в разные периоды годового цикла (1998-2003)
-
7. Исследования химического состава и биохимических свойств песчанки
Песчанка – рыба, также характерная для Кольского залива; относится к мелким морским стайным рыбам, преимущественно к арктическо-бореальным. Обитает в основном у берегов на песчаном или мелкокаменистом грунте. У берегов Мурмана нерест морской песчанки происходит зимой – с ноября по февраль ( Промысловые рыбы …, 1952).
-
8. Заключение
Проведен анализ химического состава мышечной ткани песчанки, характерной для вод Кольского залива. Изучен химический состав песчанки, выловленной в период нереста (массовая доля влаги – 72,0 ± 2,5 %, жир – 5,7 ± 0,2 %, белок – 16,8 ± 0,3 %, зола – 2,3 ± 0,2 %). По химическому составу мяса песчанка – белковая среднежирная рыба ( Лебская и др. , 1993). Сравнивали полученные результаты с литературными данными. Для периода 1998-2003 гг. по сравнению с периодом до 1998 г. наблюдается снижение содержания влаги в среднем на 8 %, содержания белка на 9 %, золы – на 14 %, увеличение содержания жира на 57 %, золы – на 31 %.
Проведено изучение сезонной динамики химического состава и биохимических свойств ряда промысловых видов рыб Баренцева моря.
Исследована сезонная динамика влагосодержания (1998-2003) в мышечной ткани гидробионтов. Установлено, что наибольшее содержание влаги для мышечной ткани мойвы, пикши, трески, камбалы наблюдается в посленерестовый период. Наименьшее содержание влаги отмечено в конце периода нагула (с 1998 по 2003 гг.) и зимовки. Для трески минимальное содержание влаги отмечено в преднерестовый период, для камбалы – в нерестовый. Колебания между минимальными и максимальными значениями влагосодержания в течение годового цикла составляют 10-14 %.
Исследована сезонная динамика содержания белка в мышечной ткани гидробионтов в зависимости от периода годового цикла. Наименьшее содержание белка для мойвы, камбалы, пикши отмечено в посленерестовый период (май-июнь), для мышечной ткани трески – в период нереста. Наибольшее содержание белковой фракции в мышечной ткани пикши, трески и камбалы наблюдается перед нерестом, для мойвы – в зимний период.
Исследована сезонная динамика содержания жира в мышечной ткани гидробионтов в зависимости от периода годового цикла. Для мойвы в период нагула и зимовки (осень) наблюдается наиболее высокое содержание липидов. Самое низкое содержание липидной фракции для всех исследованных видов рыб характерно для нерестового и посленерестового периода.
Для пикши, трески и камбалы наибольшая массовая доля липидов отмечена в зимний и преднерестовый период. Колебания между минимальными и максимальными значениями жиросодержания в течение годового цикла составляют для трески и пикши 27 %, для камбалы до 60 %, для мойвы до 90 %.
Анализ проведенного нами расчета СО/В для разных периодов жизненного цикла мойвы показывает уменьшение данного показателя от осеннего к весеннему периоду. Увеличение СО/В наступает в период нагула.
На основе анализа данных по динамике содержания макроэргических соединений в мышечной ткани гидробионтов установлено, что наиболее высокое содержание аденозинтрифосфорной кислоты характерно для преднерестового периода и нереста; наименьшее содержание макроэргов наблюдается в посленерестовый период (истощенное состояние рыбы), в период нагула имеет место увеличение количества АТФ по сравнению с посленерестовым промежутком времени.
Проанализирована динамика содержания минеральных веществ в мышечной ткани промысловых рыб. Содержание золы, по нашим данным, колеблется от 0,85-1,0 до 1,1-1,65, в среднем составляя 1,2 % (для песчанки – 2,3 %).
Собственные данные, полученные в 1998-2003 гг. сравнивали с литературными данными (до 1998 г.).
При сравнении полученных нами данных по влагосодержанию влаги за период с 1998 по 2003 гг. с данными ПИНРО до 1998 г. установлено, что для мойвы наблюдается увеличение содержания влаги в среднем на 1,3 %. Содержание влаги в посленерестовый период совпадает с данными ПИНРО. Для пикши и трески отмечено общее снижение содержания влаги в исследованный период нами период в среднем на 5 %, для камбалы – на 2 %.
В отношении содержания белка отмечено снижение его концентрации в исследованный период для мойвы, трески и камбалы на 3, 7 и 8 % соответственно. Для пикши отмечено снижение содержания белка в посленерестовый период на 9,4 % и увеличение в зимний период на 7 %.
При сравнении полученных нами данных по содержанию липидов за период с 1998 по 2003 гг. с данными ПИНРО до 1998 г. установлено, что для мойвы наблюдается увеличение содержания жира в среднем на 8 %. Для пикши отмечено снижение содержания липидов до 40 %, а для трески – увеличение.
Полученные нами результаты по содержанию минеральных веществ соответствуют данным ПИНРО. Зависимости от стадии жизненного цикла не наблюдается. По данным ПИНРО, содержание минеральных веществ колеблется от 1,14 до 1,47 %, составляя в среднем 1,3 %.
Результаты исследований вышеперечисленных гидробионтов подтверждают сезонный характер динамики содержания основных химических показателей (общего азота, белкового азота, небелкового азота, аминного азота, липидов и других).
Нами установлено некоторое отличие химического состава данных промысловых рыб в период с 1998 по 2003 гг. по сравнению с периодом до 1998 г.
Впервые проанализирована динамика содержания макроэргических соединений и активности тканевых протеолитических ферментов в мышечной ткани гидробионтов Севера. Данные характеристики предложены в качестве биохимических маркеров, характеризующих ответные реакции организмов гидробионтов на антропогенный стресс.
Проведенные исследования промысловых гидробионтов, направленные на выявление особенностей их химического состава и биохимических свойств с целью установления характерных биохимических маркеров, вносят вклад в решение оценки состояния гидробионтов Северного бассейна и в разработку единой комплексной биохимической системы тестирования и биоиндикации водных экосистем Кольского Севера.
