Биоиндикаторы стадий разрушения ветровально-буреломной древесины ели европейской в хвойно-широколиственных лесах Московской обл

Автор: Некляев С.Э.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесоведение и лесоводство
Статья в выпуске: 1, 2023 года.
Бесплатный доступ
Для выявления последовательности разрушения древесины и определения организмов, способных выступать в качестве биоиндикаторов, обследованы 65 модельных деревьев ели европейской в зоне хвойно-широколиственных лесов Московской обл. Разработан метод исследования модельного дерева, позволяющий получить данные, характеризующие степень разрушения древесины в результате влияния биологических и абиотических факторов. Предложена шкала стадий разрушения древесины ели под воздействием ксилотрофных базидиомицетов и сапроксильных насекомых. На основе показателей модельных деревьев выделены биоиндикаторы стадий биодеструкции и последовательность их смены в зависимости от состояния древесного субстрата. Сделан вывод о том, что ксилолиз является единым динамическим процессом, происходящим под воздействием комплексов мико- и зоодеструкторов. При этом определить биоиндикаторы можно только для 1-4-й стадий разрушения древесины. Для 5-й стадии характерна бурая мягкая гниль с уровнем разложения древесины более 80 %, что делает выявление биоиндикаторов ксилолиза в полевых условиях невозможным.
Ксилолиз, афиллофоровые грибы, сапроксильные насекомые
Короткий адрес: https://sciup.org/143179854
IDR: 143179854 | УДК: 632.915 | DOI: 10.24419/LHI.2304-3083.2023.1.02
Bioindicators of the stages of destruction of wind-blown windbreak wood of European spruce in coniferous-deciduous forests of the Moscow region
The process of natural destruction of wood combines two lines of decomposition of wood matter: mycogenic destruction of wood, which occurs under the influence of xylotrophs and mechanical destruction of the structure of wood matter during the vital activity of xylobionts. To identify the sequence of destruction and identify organisms capable of acting as bioindicators, 65 model trees of European spruce in the zone of coniferous-deciduous forests of the Moscow region were examined. To conduct the study, a method for studying a model tree has been developed that allows to obtain integrated data on the destruction of wood under the influence of biological and abiotic factors. A scale of stages of the destruction of spruce wood under the influence of xylotrophic basidiomycetes and saproxyl insects is proposed. Based on data from model trees, bionditactors of biodestruction stages and the sequence of their change depending on the state of the wood substrate are identified. It is concluded that xylolysis is a single dynamic process that takes place under the influence of complexes of mycodestructors and zoodestructors. Determination of bioindicators is possible for 1-4 stages of destruction of wood. Stage 5 is characterized by brown soft rot with a wood decomposition level of more than 80 %, which makes it almost impossible to identify biological indicators of xylolysis in the field.
Текст научной статьи Биоиндикаторы стадий разрушения ветровально-буреломной древесины ели европейской в хвойно-широколиственных лесах Московской обл
М етодики определения степени разрушения древесины не отражают многофакторный процесс биодеструкции, происходящий под воздействием сапроксильных организмов. Это связано в тем, что изучение процесса естественного разрушения древесины ствола ведется по двум почти не соприкасающимся направлениям: микогенной биодеструкции древесины, происходящей под воздействием ксилотрофов [1–8], и механического разрушения структуры древесного вещества в ходе жизнедеятельности ксилобионтов [5, 9, 10].
В конце ХХ в. было предложено несколько теорий о взаимовлиянии ксилотрофов и ксило-бионтов на процесс биодеструкции древесины ствола. В частности, для каждой стадии ослабления дерева и биодеструкции древесины могут быть определены индикаторные ксилотрофные базидомицеты и ксилофаги [11–17].
Более глубокое изучение биоиндикаторов стадий биодеструкции необходимо для формирования системного понимания биологического разрушения древесины как комплексного процесса возвращения вещества в природную среду под воздействием пула сапроксильных организмов, последовательно сменяющих друг друга при переходе от одной стадии разложения древесины к другой.
Цель исследования – выявление индикаторных видов сапроксильных организмов, активно участвующих в биодеструкции древесины ветровально-буреломных деревьев ели европейской. Для решения задачи проведен анализ модельных деревьев и определены признаки изменения структуры древесины на разных стадиях разложения под воздействием сапроксильного комплекса. В частности, необходимо было детализировать структуру и состав сапроксильного комплекса ели европейской на основе исследования полученных образцов повреждений, базидиом и сбора насекомых с модельных деревьев.
Материалы и методы
Исследования процесса биодеструкции проведены в 2019–2021 гг. в Бородинском,
Виноградовском, Егорьевском, Клинском, Московском учебно-опытном, Орехово-Зуевском и Шатурском лесничествах. Объекты исследований – 65 модельных деревьев ели европейской ( Picea abies ).
С целью изучения скорости биодеструкции стволов нами была использована методика замены временных рядов пространственными [18]. Дату образования ветровалов и буреломов устанавливали на основе данных учета лесосечного фонда по лесничествам Московской обл. Точность определения времени, прошедшего с момента отпада дерева, составляла в среднем 3 месяца.
У каждого модельного дерева измеряли длину ствола, высоту остолопа или пня для буреломных экземпляров, диаметр ствола на высоте 1,3 м от шейки корня; осуществляли замеры влажности [19] и температуры ствола. На основании положений ГОСТ 18610–82 у каждого модельного дерева устанавливали индекс состояния древесины.
Для выявления сапроксильных зоодеструкторов на модельных деревьях закладывали палетки и отбирали образцы древесины на высоте 1, 3, 6, 12, 18, 24 м. На каждом уровне проводили учет всех повреждений, нанесенных сапроксиль-ными организмами. Шаг закладки палеток и от-рубков обусловлен дальнейшим отбором с этих участков контрольных отрубков для определения стадии [14, 16, 17, 19] и типа разложения древесины (коррозионного или деструктивного) [20]. У отобранных образцов определяли объем гнили. Сильно разложившиеся фракции собирали при помощи почвенного бура. На модельных деревьях было заложено 330 палеток для определения ксило- и сапротрофов, отобрано 978 образцов для установления стадии и типа разложения, а также пространственной геометрии ходов, взято 245 кернов для определения структуры гнили.
Объем образцов рассчитывали по формулам усеченного конуса и параллелепипеда. Объем сильно разложившихся образцов принимали равным объему почвенного бура.
Стадии разложения и фрагментации коры стволов оценивали по всей длине каждого модельного дерева. Долю сохранившейся коры
(% от видимой поверхности ствола) оценивали визуально. Этот же метод был использован для определения степени покрытия модельного дерева синузиями мхов и лишайников.
С модельных деревьев собрали 922 бази-диомы ксилотрофов и определили их видовую принадлежность по морфологическому строению [21–23]. Терминология грибов приведена в соответствии с Index Fungorum [24]. Кроме того, с модельных деревьев собрали 189 личинок, 147 имаго сапроксильных насекомых и провели идентификацию видовой принадлежности по морфологическому строению [25].
Результаты и обсуждение
Процесс разрушения древесины стволов ели происходит под воздействием таких основных биотических факторов, как комплекс дереворазрушающих грибов и комплекс ксилобионтов, а также абиотических факторов – температуры и влажности воздуха, которые влияют на жизнедеятельность сапроксильных организмов. Данные комплексы играют роль агентов деструкции древесины. В процессе ксилолиза последовательно участвуют 5 групп агентов: агенты, подготавливающие среду; первичные деструкторы живой древесины; вторичные деструкторы мертвой твердой древесины; третичные деструкторы мертвой мягкой древесины; потребители продуктов распада мягкой влажной древесины.
В процессе микогенной деструкции выделяют 5 стадий [1, 3, 8, 16, 17, 26, 27].
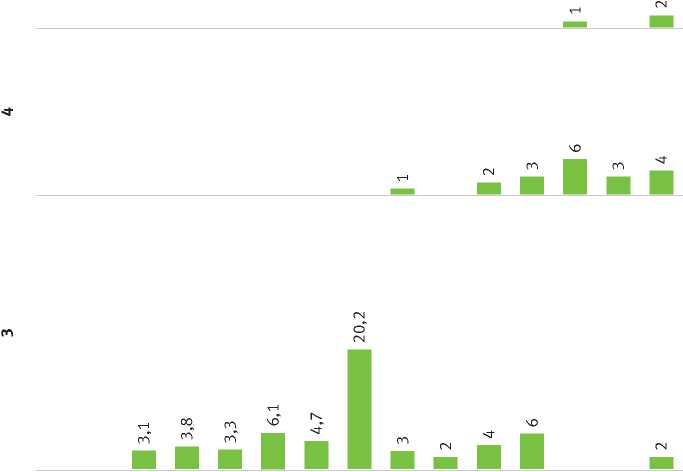
В результате исследования модельных деревьев было определено, что фитопатогенный тип разрушения древесины ограничивается 2-й стадией разложения древесного вещества. При этом на данной стадии облигатные паразиты или завершают свой жизненный цикл, или переходят к существованию в качестве факультативных сапротрофов. Сапротрофный тип разложения может быть определен не ранее перехода на 2-ю стадию разложения, так как в этот период происходит активное заселение сапротрофами: Fomitopsis pinicola, Rhodofomes roseus [=Fomitopsis rosea], Trichaptum abietinum. Данные виды сохраняют доминирование на 2–4-й стадиях разложения. Переход от стадии к стадии можно проследить по освоению ими субстрата. Так, для 2-й стадии характерно развитие красной гнили в древесине с сохранением ее физико-химических свойств, на 3-й стадии образуется бурая сухая твердая гниль, а на 4-й – мягкая бурая гниль. На 5-й стадии доминирование перейдет к подстилочным сапротрофам рода Postia [=Oligoporus] и Neoantrodia serialis [=Antrodia serialis], так как распад древесины составляет 80–100 % (табл. 1).
Анализ собранных образцов показал, что на 2–5-й стадиях основными биодеструкторами являются ксилотрофные базидиомицеты. При этом Heterobasidion parviporum, Phellinus chrysoloma, Armillaria mellea , выступающие на 1-й стадии как явные паразиты, переходят ко 2-й стадии как факультативные сапротрофы. Участие видов фитопатогенного типа разрушения в процессе ксилолиза обусловливает гибель дерева и частично подготавливает субстрат для заселения сапротрофными базидиомицетами (рис. 1).
В результате анализа можно выделить индикаторные виды афиллофоровых грибов для каждой стадии разрушения древесины (табл. 2).
По воздействию доминантных семейств на древесное вещество процесс зоогенной деструкции можно разделить на 5 стадий: 1-я – сколитид-ная и начало церамбицидной; 2-я – завершающая часть церамбицидной; 3-я – церамбицидная стадия разложения мертвой древесины; 4-я стадия – луканидно-скарабеидная; 5-я – лумбрицидная [9, 10, 11, 28] (табл. 3).
Анализ собранных образцов насекомых и повреждений древесины ксилофильными и са-проксильными насекомыми позволил выявить ряд закономерностей. В частности, на 1-й стадии зоодеструкции доминирует короедная группа, которую на 2-й стадии последовательно сменяют представители семейства Cerambycidae родов Monochamus , Tetropium , потребляющие живую древесину, а также Sirex gigas . На 3-й стадии зоодеструкции ведущая роль переходит к семейству Anobiidae и представителям семейства
Таблица 1. Распределение сапроксильных патогенов по стадиям микогенной деструкции








Рис. 1. Частота встречаемости микодеструкторов на модельных деревьях, %
Таблица 2. Индикаторные признаки стадий микогенной деструкции древесины ели
|
Стадия |
Сохранность коры, % |
Состояние древесины |
Тип (доля гнили, %) |
Индикаторные виды |
|
1 |
90–100 |
Древесина с плотной корой, отмершая в текущем году |
Деревоокрашивающие грибы (синева) и биотрофы (0–1) |
Heterobasidion parviporum, Phellinus chrysoloma, Armillaria mellea |
|
2 |
60–90 |
Древесина плотная, но с видимыми признаками деструкции |
Красная гниль (1–10) |
Fomitopsis pinicola, Trichaptum abietinum |
|
3 |
30–60 |
Верхний слой древесины мягкий, кора местами отпала |
Бурая сухая твердая гниль (10–40) |
Rhodofomes roseus [= Fomitopsis rosea ] , Trichaptum abietinum, Stereum sanguinolentum, Gloeophyllum sepiarium |
|
4 |
10–30 |
Разложение, оцениваемое визуально, проникает на значительную глубину, гниль пластинчатая или призматическая |
Бурая сухая мягкая гниль (40–80) |
Rhodofomes roseus [= Fomitopsis rosea ] , Trichaptum abietinum, Stereum sanguinolentum, Gloeophyllum sepiarium |
|
5 |
- |
Остается лишь форма ствола, на поверхности обычно хорошо развиты синузии мхов и лишайников |
Бурая влажная мягкая гниль (80–100) |
- |
Таблица 3. Распределение сапроксильных беспозвоночных по стадиям зоогенной деструкции
|
Стадия |
Виды зоодеструкторов |
|
Сколитидная стадия разложения коры. Начало церамбицидной стадии разложения древесины |
Dendroctonus micans, Hylastes cunicularius, Ips typographus, Pityogenes chalcographus, Pityophtorus micrographus, Polygraphus polygraphus, Trypodendron lineatum, Monochamus sutor, Monochamus urussovii, Callidium (Callidostola) aeneum, Callidium violaceum, Rhagium inquisitor, Tetropium castaneum, Anthaxia quadripunctata |
|
Завершение церамбицидной стадии разложения древесины |
Monochamus sutor, Monochamus urussovii, Callidium (Callidostola) aeneum, Callidium coriaceum, Callidium violaceum, Rhagium inquisitor, Tetropium castaneum, Anthaxia quadripunctata, Molorchus minor, Sirex gigas |
|
Церамбицидная стадия разложения мертвой древесины (твердая гниль) |
Anobium punctatum, Hylotrupes bajulus, Oxymirus cursor, Stictoleptura rubra, Anoplodera sanguinolenta, Peltis grossa, Peltis ferruginea, Ostoma ferruginea, Serviformica fusca |
|
Луканидно-скарабеидная стадия разложения древесины (мягкая гниль) |
Ceruchus chrysomelinus, Denticollis linearis, Ampedus pomonae, Ampedus pomorum, Ampedus balteatus, Ampedus sanguineus, Oxymirus cursor, Stictoleptura rubra, Pyrrhidium sanguineum, Hylis procerulus, Peltis grossa, Peltis ferruginea, Ostoma ferruginea, Ceruchus chrysomelinus, Platyrhinus resinosus, Rhyncolus ater |
|
Лумбрицидная стадия разложения древесины (влажная гниль) |
Кивсяки, дождевые черви, мокрицы |
Cerambycidae триб Hylotrupini и Callidiini, которые предпочитают мертвую твердую древесину. На 4-й стадии в древесине преобладают представители семейства Elateridae и подсемейства Lepturinae. Под оставшимися участками коры как на 3-й, так и на 4-й стадиях часто обитают представители семейства Trogossitidae рода Peltis. На 5-й стадии единично встречаются представители семейства Elateridae, но только при сохранении незначительной части древесного вещества, а также в большом количестве дождевые черви и кивсяки (рис. 2).
Таким образом, выявленные виды сапрок-сильных насекомых можно распределить по стадиям механической зоодеструкции древесины (табл. 4).
Биоиндикаторы стадий разрушения ветровально-буреломной древесины ели европейской в хвойно-широколиственных лесах Московской обл.
Стадия
Среднее количество:
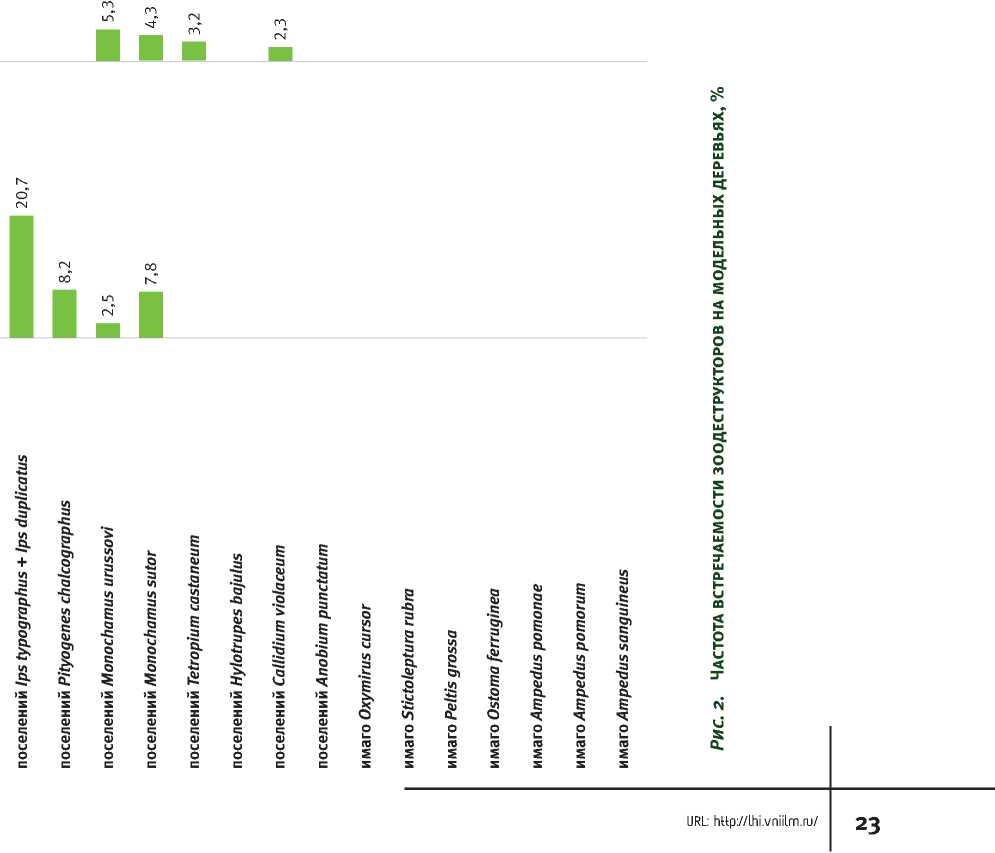
Таблица 4. Индикаторные виды стадий зоодеструкции древесины ели
|
Стадия |
Сохранность коры, % |
Состояние древесины |
Индикаторные виды |
|
1 |
90–100 |
Древесина с плотной корой, отмершая в текущем году |
Ips typographus, Ips duplicatus, Pityogenes chalcographus |
|
2 |
60–90 |
Древесина плотная, но с видимыми признаками деструкции |
Monochamus urussovi, Monochamus sutor, Tetropium castaneum |
|
3 |
30–60 |
Верхний слой древесины мягкий, кора местами отпала |
Hylotrupes bajulus, Anobium punctatum, Peltis grossa, Ostoma ferruginea |
|
4 |
10–30 |
Разложение, оцениваемое визуально, проникает на значительную глубину, гниль пластинчатая или призматическая |
Oxymirus cursor, Stictoleptura rubra, Ampedus pomonae, Ampedus pomorum, Ampedus sanguineus, Peltis grossa, Ostoma ferruginea |
|
5 |
- |
Остается лишь форма ствола, на поверхности обычно хорошо развиты синузии мхов и лишайников |
- |
Сапроксильные насекомые играют важную роль в развитии микодеструкторов, хотя и не могут выступать в качестве ведущей силы деструкции древесины ели. Причем эта роль наиболее значительна в качестве ускоряющего фактора разложения древесины дереворазрушающими грибами как за счет распространения спор внутри ходов, так и за счет повышения аэрации и влажности древесины, что улучшает условия дыхания и развития ксилотрофов [10, 11, 29–34].
Основываясь на предыдущих работах, а также данных, полученных на модельных деревьях, можно предложить следующую последовательность стадий биодеструкции под воздействием зоо- и микодеструкторов (табл. 5).
Для выявления степени воздействия мико-и зоодеструкторов необходимо ввести расчет коэффициента биодеструкции (∆Ξ). Этот коэффициент позволяет установить стадию разрушения на основе показателей измерений модельных деревьев.
Для определения коэффициента биодеструкции нужно рассчитать коэффициенты микодеструкции и зоодеструкции. Коэффициент микодеструкции (∆Mct) определяют по фактическому объему гнили в модельном дереве в соответствии с предлагаемой шкалой, т. е.: ∆Mct = стадия ми-когенной деструкции.
Коэффициент зоодеструкции (∆Int) рассчитывают как средневзвешенную категорию среднего количества ходов на модельном образце на каждой стадии:
AInt = (Scx1+Cb1x2+Cb2x3+LSx4+Lmx5)/100, где:
Sc – средняя площадь повреждения камбия ствола поверхностными деструкторами в сколитидной стадии разложения коры и начале церамбицидной стадии разложения древесины на модельном образце;
Cb1 – средний объем повреждения заболонной древесины первичными деструкторами в завершающей части церамбицидной стадии разложения древесины на модельном образце;
Cb2 – средний объем повреждения древесины вторичными деструкторами в церамбицидной стадии разложения древесины на модельном образце;
LS – среднее количество третичных деструкторов в луканидно-скарабеидной стадии разложения древесины на модельном образце;
Lm – среднее количество потребителей продуктов распада в лумбрицидной стадии разложения древесины на модельном образце.
Коэффициент биодеструкции (∆Ξ) рассчитывают по следующей формуле:
∆Ξ = (∆Int+∆Mct)/1,5.
Деление суммы коэффициентов микогенно-го и зоогенного разрушения древесины ствола на
Таблица 5. Шкала стадий микогенной и зоогенной деструкции
|
Стадия |
Сохранность коры, % |
Состояние древесины |
Период протекания стадии (среднее значение) |
Стадия зоогенной деструкции |
Стадия микогенной деструкции (доля гнили, %) |
|
1 |
90–100 |
Древесина с плотной корой, отмершая в текущем году |
1–2 года (1,5) |
Сколитидная стадия разложения коры, начало церамбицидной стадии разложения древесины первичных деструкторов древесины (поверхностные деструкторы) |
Деревоокрашивающие грибы (синева) и биотрофы (0–1) |
|
2 |
60–90 |
Древесина плотная, но с видимыми признаками деструкции |
2–3 года (2,5) |
Завершение церамбицидной стадии разложения древесины первичными деструкторами (первичные деструкторы древесины) |
Красная гниль (1–10) |
|
3 |
30–60 |
Верхний слой древесины мягкий, кора местами отпала |
4–6 лет (5) |
Церамбицидная стадия разложения древесины вторичными деструкторами (вторичные деструкторы мертвой древесины) |
Бурая сухая твердая гниль (10–40) |
|
4 |
10–30 |
Разложение, оцениваемое визуально, проникает на значительную глубину, гниль пластинчатая или призматическая |
4–8 лет (6) |
Луканидно-скарабеидная стадия разложения древесины третичными деструкторами (третичные деструкторы – сапроксилофаги) |
Бурая сухая мягкая гниль (40–80) |
|
5 |
Остается лишь форма ствола, на поверхности обычно хорошо развиты синузии мхов и лишайников |
8–10 лет (9) |
Лумбрицидная стадия разложения древесины, гумификаторы (потребители продуктов ксилолиза) |
Бурая влажная мягкая гниль (80–100) |
1,5 объясняется тем, что зоодеструкция является фактором, ускоряющим процесс развития мико-генного ксилолиза. Механическое разрушение беспозвоночными способствует проникновению спор и мицелия дереворазрушающих грибов в за-болоневую и ядровую древесину.
Расчет соотношения коэффициента биодеструкции (∆Ξ) и коэффициентов микогенного (∆Mct) и зоогенного (∆Int) разрушения древесины ствола показал связь между микогенной и зоогенной деструкцией, отражающую ход естественного разрушения древесины ствола. При этом коэффициент микодеструкции демонстрирует значимость дереворазрушающих грибов в процессе деструкции древесины, а коэффициент зоодеструкции иллюстрирует процесс внутреннего истачивания древесины ходами сапроксильных беспозвоночных, что влияет на скорость распространения гнили внутри ствола (рис. 3.)
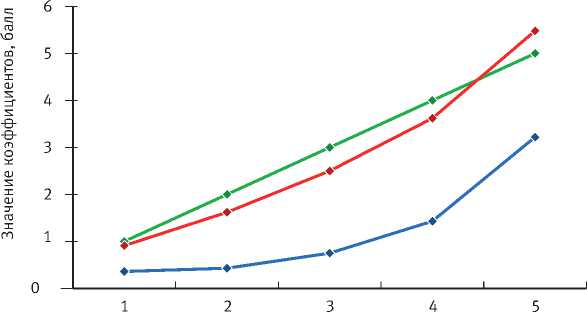
Стадия
^—»^— Коэффициент биодеструкции (∆ Ξ )
---■♦■--- Коэффициент микодеструкции (∆Mct)
----♦’--- Коэффициент зоодеструкции (∆int)
Рис. 3. Соотношение коэффициента деструкции (∆ Ξ ) и коэффициентов микогенного (∆Mct) и зоогенного (∆Int) разрушения древесины ствола
Выводы
По результатам исследования установлено, что ксилолиз является единым динамическим процессом, происходящим под воздействием комплексов микодеструкторов и зоодеструкторов.
Индикаторами стадий ксилолиза древесины со стороны микодеструкторов выступают: 1-я стадия – Heterobasidion parviporum, Phellinus chrysoloma, Armillaria mellea; 2-я стадия – Fomitopsis pinicola, Trichaptum abietinum; 3–4-я стадии – Rhodofomes roseus [= Fomitopsis rosea ] , Trichaptum abietinum, Stereum sanguinolentum, Gloeophyllum sepiarium .
Индикаторами стадий биодеструкции древесины со стороны зоодеструкторов выступают: 1-я стадия – Ips typographus, Ips duplicatus, Pityogenes chalcographus; 2-я стадия – Monochamus urussovi, Monochamus sutor, Tetropium castaneum; 3-я стадия – Hylotrupes bajulus, Anobium punctatum, Callidium violaceum, Peltis grossa, Ostoma ferruginea; 4 стадия – Oxymirus cursor, Stictoleptura rubra, Ampedus pomonae, Ampedus pomorum, Ampedus sanguineus, Peltis grossa, Ostoma ferruginea.
Для 5-й стадии характерна бурая мягкая гниль с разложением древесины более 80 %, что делает выявление в полевых условиях биологических индикаторов ксилолиза почти невозможным. Относить образцы к данной стадии следует в зависимости от доли гнили ствола, наличия мхов и лишайников, сохранности формы ствола.
Список литературы Биоиндикаторы стадий разрушения ветровально-буреломной древесины ели европейской в хвойно-широколиственных лесах Московской обл
- Биодеградация древесины ферментными комплексами дереворазрушающих грибов / А.Н. Веревкин, Г.Н. Кононов, Ю.В. Сердюкова [и др.] // Лесной вестник. - 2019. - Т. 23. - № 5. - С. 95-100.
- Boddy, L. Ecology of Saprotrophic Basidiomycetes / L. Boddy, J. Frankland, P. Van West. - Amsterdam: Elsevier, 2007. - 386 p.
- Gavrilov, T.A. The influence of natural and climatic factors on the wood biodestruction process / T.A. Gavrilov, T.B. Stankevich. - Text: electronic // Journal of Agriculture and Environment. - 2022. - № 5 (25). - Available at: https://jae.cifra.science/index.php/jae/article/view/284
- Kisternaya, M. Biodeterioration of historic timber structures: A comparative analysis / M. Kisternaya, V. Kozlov // Wood Material Science and Engineering. - 2014. - Vol. 9. - Issue 3. - P. 156-161.
- Maeda, K. Relationship between the mass profile and the strength property profile of decayed wood / K. Maeda, M. Ohta, I. Momohara // Wood Science and Technology. - 2015. - Vol. 49. - P. 331-344.
- Types and rates of decomposition of Larix sibirica trees and logs in a mixed European boreal old-growth forest / E. Shorohova, E. Kapitsa, A. Ruokolainen, I. Romashkin, I. Kazartsev // Forest Ecology and Management. - 2019. - Vol. 439. - P. 173-180.
- Sippola, A.-L. Wood-decomposing fungi and seed-tree cutting: a 40-year perspective / A.-L. Sippola, P. Renvall // Forest Ecology and Management. - 1999. - Vol. 115. - P. 183-201.
- The roles of fungi and animals in the immobilisation and release of nutrient elements from decomposing branch-wood / M.J. Swift, U. Lohm, T. Persson [and etc.] // Soil Organisms as Components of Ecosystems. Ecological Monographs. - 1977. - Vol. 25. - P. 193-202.
- Неволина, Н.Б. Видовой состав и экологическая структура жесткокрылых-ксилобионтов в хвойно-широколиственных лесах / Н.Б. Неволина // Экология, мониторинг и рациональное природопользование: науч. тр. - Вып. 307. - Москва: МГУЛ, 2001. - С. 30-37.
- Stokland, J.N. Biodiversity in dead wood / J.N. Stokland, J. Siitonen, B.G. Jonsson. - Cambridge: Cambridge University Press, 2012. - 509 p.
- Красуцкий, Б.В. Ксилофильные и мицетофильные жесткокрылые Висимского заповедника / Б.В. Красуц-кий // Экология процессов биологического разложения древесины. - Екатеринбург, 2006. - С. 110-133.
- Грибы и насекомые - консорты лесообразующих древесных пород Карелии / В.И. Крутов, В.И. Шубин, О.О. Предтеченская [и др.]. - Петрозаводск: КарНЦ РАН, 2014. - 216 с.
- Christensen, M. Study on Dead Wood in European Beech Forest Reserves / M. Christensen. - Denmark: University of Copenhagen, 2013. - 30 p.
- Ehnstrom, B. Life after death - an insect drama / B. Ehnstrom. - Stockholm: Media Express Frlag och Information, 1998. - 116 p.
- Fungus-Insect Relationship. Perspectives in Ecology and Evolution / ed. by Q. Wheeler, M. Blackwell. - New York: Columbia University Press, 1984. - 516 p.
- Schmidt, O. Wood and Tree Fungi. Biology, Damage, Protection, and Use / О. Schmidt // Springer. - 2006. - P. 334.
- Zabel, R.A. Wood Microbiology. Decay and Its Prevention / R.A. Zabel, J.J. Morrell, S. Robinson. - London: Elsevier Academical Press, 2020. - 556 p.
- Тарасов, М.Е. Методические подходы к определению скорости разложения древесного детрита / М.Е. Тарасов // Лесоведение. - 2002. - № 5. - С. 32-38.
- Линдеман, Г.В. Взаимоотношения насекомых-ксилофагов и лиственных деревьев в засушливых условиях / Г.В. Линдеман. - Москва: Наука, 1993. - 205 с.
- Стороженко, В.Г. Древесный отпад в коренных лесах Русской равнины / В.Г. Стороженко. - Москва: КМК, 2011. - 122 с.
- Бондарцева, М.А. Определитель грибов России: Порядок афиллофоровые/ М.А. Бондарцева. - Санкт-Петербург: Наука, 1998. - 391 с.
- Ниемеля, Т. Трутовые грибы Финляндии и прилегающих территорий России / Т. Ниемеля. - Хельсинки: Helsinki University Printing House, 2001. - 120 c.
- Атлас-определитель дереворазрушающих грибов Русской равнины / В.Г. Стороженко, В.И. Кругов, А.В. Руоколайнен, В.М. Коткова, М.А. Бондарцева. - Москва: КМК, 2014. - 198 с.
- Index Fungorum - http://www.indexfungorum.org.
- Иллюстрированный справочник жуков-ксилофагов - вредителей леса и лесоматериалов Российской Федерации / С.С. Ижевский, Н.Б. Никитский, О.Г. Волков, М.М. Долгин. - Тула: Гриф и К, 2005. - 220 с.
- Hatakka, A. Fungal biodegradation of lignocelluloses / A. Hatakka, K.E. Hammel, M. Hofrichter (ed.) // Industrial Applications (The Mycota). - 2011. - Vol. 10. - P. 319-340.
- Renvall, P. Community structure and dynamics of wood-rotting Basidiomycetes on decomposing conifer trunks in northern Finland / P. Renvall // Karstenia. - 1995. - Vol. 35. - P. 1-51.
- Yaxley, B. Biology and Conservation Ecology of Selected Saproxylic Beetle Species in Tasmania's Southern Forests / B. Yaxley. - Australia: School of Zoology, University of Tasmania, 2013. - 262 p.
- Dajoz, R. Insects and Forests: The Role and Diversity of Insects in the Forest Environment / R. Dajoz. - Paris: Intercept Limited, 2000. - 620 р.
- Carbon concentration of standing and downed woody detritus: effects of tree taxa, decay class, position, and tissue type / M.E. Harmon, B. Fasth, C.W. Woodall, J. Sexton // Forest Ecology and Management. - 2013. -Vol. 291.-P. 259-267.
- Hunter, M.L. Wildlife, Forests, and Forestry. Principles of Managing Forests for Biological Diversity / M.L. Hunter. - Englewood Cliffs, New Jersey: Prentice-Hall ; Simon & Schuster, 1990. - 370 p.
- Ódor, P. Preferences of Dead Wood Inhabiting Bryophytes to Decay Phase, Log Size and Habitat Types in Hungarian Beech Forests/ P. Ódor, A.F.M. van Hees // Journal of Bry. - 2013. - P. 79-95.
- Siitonen, J. Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example / J. Siitonen // Ecological bulletins. - 2001. - Vol. 49. - P. 11-41.
- Yatskov, M.A. Chronosequence of wood decomposition in the boreal forests of Russia / M. Yatskov, M.E. Harmon, O.N. Krankina // Canadian Journal of Forest Research. - 2003. - Vol. 33. - P. 1211-1226.
