Биокристаллоскопические методы в изучении кристаллогенных свойств микроорганизмов

Автор: Мартусевич Андрей Кимович, Симонова Жанна Георгиевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Природопользование
Статья в выпуске: 5-3 т.13, 2011 года.
Бесплатный доступ
Приведены сведения о возможностях оценки кристаллогенной активности микроорганизмов с помощью специальных методов, относящихся к новой медико-биологической науке – биокристалломике. Примерами тестовых микроорганизмов выступили Escherichia сoli и Staphylococcus aureus, для которых проведен сравнительный анализ их кристаллогенных свойств.
Биокристалломика, метаболизм микроорганизмов, кристаллогенные свойства
Короткий адрес: https://sciup.org/148200477
IDR: 148200477 | УДК: 579.22
Biocrystalloscopic methods in investigation of microorganisms crystallogenic properties
In this paper data about possibilities of microorganisms crystallogenic activity estimation by special biocrystalloscopic methods are given. We tested crystallogenic properties of E. сoli and St. aureus comparatively.
Текст научной статьи Биокристаллоскопические методы в изучении кристаллогенных свойств микроорганизмов
(штаммы получены в Кировском НИИ гематологии и переливания крови под руководством зав. лабораторией консервации крови и тканей д.м.н. А.А. Костяева). Подобный выбор биообъектов неслучаен: E.сoli является типичным представителем ау-тохтонной микрофлоры организма человека и животных, тогда как S.aureus представляет собой классический патогенный бактериальный агент. В целях уточнения роли концентрации бактерии были использованы единые для обоих микроорганизмов тестовые количества биомикрообъектов: 106, 108, 1010, 1012, 1014, 1016, 1018 и 1020 КОЕ/мл раствора. Подобный дифференцированный подход позволил не только сопоставить микроорганизмы между собой по инициирующей активности, но и установить их пороги инициирующих свойств путем установления минимальной концентрации бактерий, вносящей существенные изменения в характер кристаллообразования базисного вещества. В качестве инициируемого (базисного) вещества применялся 10% раствор хлорида натрия, что позволило выполнить тезиграфический тест в варианте сравнительной тезиграфии (сопоставление опытного образца смеси микроорганизмов и базисного соединения с препаратом, полученным при высушивании на том же стекле только раствора используемой соли) [3-6]. Это дало возможность количественно оценить «инициаторный потенциал» бактерии, трактуемый нами как ее способность оказывать влияние на картину кристаллизации раствора соли.
Характер микроорганизм-ассоциированной модуляции кристаллообразования гипертонического раствора хлорида натрия оценивался дуалистично: а) с позиции поиска качественных особенностей о маркеров инфицированности раствора в целом (общие для обоих микроагентов черты) и видоспецифичных изменений кристаллогенеза базисного веществ (дифференциация микроорганизмов на основании различных градаций); б) с применением количественных критериев, позволяющих верифицировать наблюдаемые отличия тезиграфических картин, что осуществлялось в соответствии с предложенным ранее алгоритмом [5].
Статистическая обработка результатов производилась алгоритмами вариационной статистики с применением встроенных функций электронных таблиц Microsoft Excel 2003, а также в специализированных пакетах SPSS 11.0 и Primer of biostatistics 4.03.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Исследования особенностей инициаторного процесса 10% раствора хлорида натрия рассматри- ваемыми бактериями в зависимости от концентрации анализируемой культуры позволили установить, что имеет место закономерная динамика нарастания выраженности сдвигов кристаллообразования гипертонического раствора при увеличении содержания модулирующего его микроагента. Важно, что данная тенденция касается обоих микроорганизмов (рис. 1, 2).
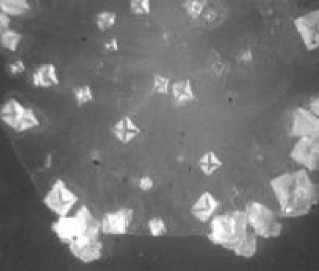
106 КОЕ/мл
1010 КОЕ/мл
1020 КОЕ/мл
Рис. 1. Кристаллогенез тезиосистем, включающих E.сoli и 10% раствор хлорида натрия (х7,5).
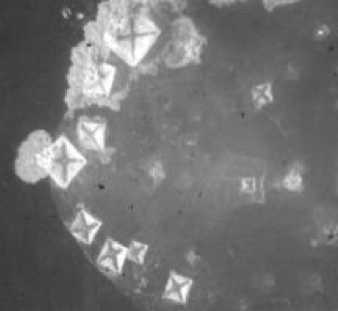
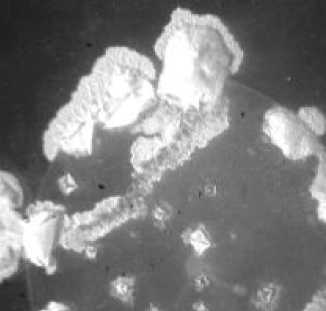
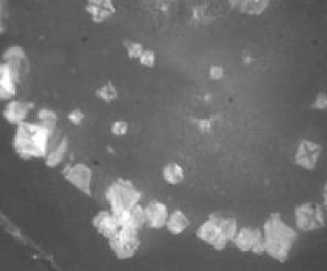
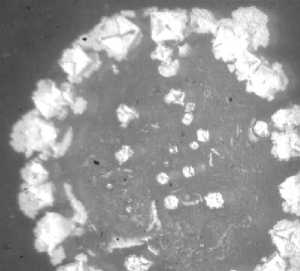
Рис. 2. Кристаллогенез тезиосистем, включающих S.aureus и 10% раствор хлорида натрия (х7,5).
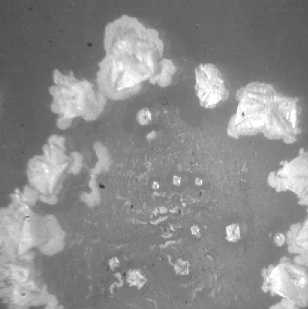
1020 КОЕ/мл
При более подробном изучении распределения тезиграмм по градации возрастающей инициирующей дозы микроагента с учетом всех контролируемых концентраций был обнаружен 2-ступенчатый характер реагирования хлорида натрия на метаболическую активность микроорганизма. Он включает первоначальное отсутствие значимых вариаций по сравнению с контрольным образцом базисного вещества. При нарастании количества микробных тел для каждого микроорганизма отчетливо регистрируется концентрация, начиная с которой появляется динамика трансформации кристаллогенеза в краевой зоне. Она выражается в формировании островков одиночных кристаллов, окружающих относительно крупные пирамидальные структуры, образованные хлоридом натрия (рис. 1, 2). Принципиально подчеркнуть, что концентрация, с которой начинают обнаруживаться подобные изменения, у эшерихии и стафилококка варьирует, причем она значительно меньше у патогенного микроагента. Так, для E.coli данная пороговая концентрация составляет 1012 КОЕ/мл (рис. 1), то относительно S.aureus даже при минимальном (из изученных)
количестве бактерий (106 КОЕ/мл) регистрируются значимые преобразования структуропостроения краевой зоны (рис. 2).
Третьим из выделенных нами вариантов инициации кристаллообразования хлорида натрия бактериями, ассоциированных с количеством микроагентов, является нарастающий неокристаллогенез (отсутствует в контрольном образце 10% раствора хлорида натрия) в центральной и промежуточной зонах высушенного препарата. На основании этого представляется возможным выделить вторую пороговую концентрацию, причем ее значение, как и для первой, видоспецифично (1014 КОЕ/мл – для E.coli и 1010 КОЕ/мл – для S.aureus ).
В соответствии с выбранной методикой описания высушенных микропрепаратов нами производилась не только качественная, но и количественная их оценка путем применения системы критериев. В качестве наиболее информативных и несущих максимальный объем биологически значимых сведений параметров рассчитаны основной тезигра-фический коэффициент (Q) и степень деструкции фации (СДФ). Первый из показателей позволяет мерой последнего. Степень деструкции фации указывает на выраженность разрушения либо дефекты формирования кристаллов и других видимых при оптической микроскопии элементов образца.
охарактеризовать направленность (по градации «активация-ингибирование») и выраженность «инициаторного потенциала» микроорганизмов. Коэффициент Q является основной количественной
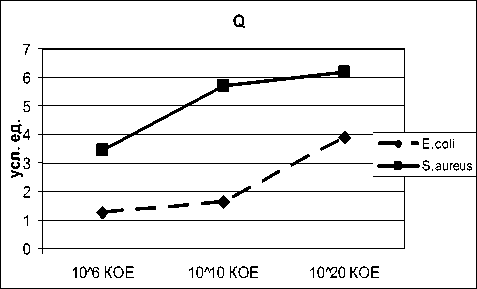
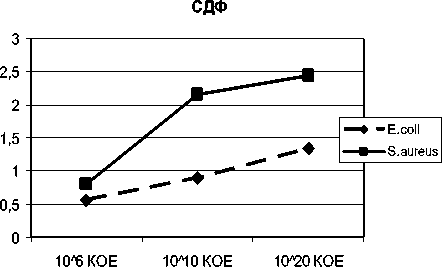
Рис. 3. Динамика нарастания основного тезиграфического коэффициента (Q) и степени деструкции (СДФ) образцов высушенных растворов, содержащих возрастающие концентрации микроорганизмов.
Проведенный с подобных позиций анализ показал, что качественные преобразования, связанные с метаболической активностью бактерий, находят полное подтверждение при оценке количественных критериев тезиграфического теста (рис. 3). В частности, монотонное нарастание уровня основного тезиграфического коэффициента является следствием сначала краевого, а затем и тотального (распространяющегося на все зоны микропрепарата) неокристаллогенеза, отражающего «инициаторный потенциал» микроорганизма. В то же время увеличение количества кристаллических образований не сопровождается повышением структурированности фации в целом, о чем свидетельствует прогрессивный рост показателя СДФ, визуализирующего качественные преобразования тезиграфической картины (рис. 3). При этом важно отметить, что выбранные микроорганизмы в отношении количественных оценочных критериев существенно различимы. Так, по основному тезиграфическому коэффициенту, а, следовательно, и по инициаторной способности (при использовании в качестве базисного вещества 10% раствора хлорида натрия) S.aureus достоверно превосходит E.coli при всех исследованных концентрациях микроагентов (p<0,05), и если на небольших значениях «инициаторный потенциал» эшерихии незначителен, то у стафилококков он исходно высок (усиление кристаллообразования хлорида натрия в 3,5 раза). При высоких концентрациях столь четкая дифференциация между бактериями несколько сглаживается за счет выраженного повышения инициаторного потенциала E.coli.
Аналогичные тенденции были обнаружены при оценке «правильности» модулируемого микроорганизмами кристаллогенеза раствора хлорида натрия (по степени деструкции фации), однако на низких концентрациях бактериальных агентов достоверных различий по рассматриваемому параметру не выявлено (p<0,05). Кроме того, представляется интересным, на наш взгляд, подчеркнуть, что патогенная бактерия (в нашем случае - золотистый стафилококк) на средних и высоких концентрациях обладает достоверно более четким деструктурирующим действием на кристаллизацию растворов хлорида натрия, чем микроорганизм-представитель аутохтонной микрофлоры многих биотопов человека и животных (кишечная палочка). Эти изменения не связаны с нарастанием «инициаторного потенциала» изучаемых агентов микромира, что верифицировано с помощью корреляционного анализа (r=-0,103±0,062), т. к. сопоставимые темпы прироста инициирующей способности бактерий с увеличением их концентрации не подтверждаются превалирующим ростом деструктивности микропрепарата, связанной с повышением количества стафилококка, аналогичной динамикой у эшери-хии.
На основании прямых модельных экспериментов с колониями отдельных патогенных и непатогенных микроорганизмов впервые описано новое явление, ранее не выделяемое - микробная инициация кристаллообразования солей неорганического ряда (феномен микроорганизм-ассоциированного кристаллогенеза [МАК]). Принципиально важен тот факт, что данное явление универсально для агентов микромира, однако он носит и специфичные черты, накладываемые видовой принадлежностью микроорганизма и его текущим функциональным состоянием. Эти представления практически нивелируют предлагаемую ранее дифференциацию биокристаллогенеза на первичный (происходящий вне зависимости от факторов внешней среды) и вторичный (обусловленный действием факторов макро- и микроокружения формирующихся кристалов) в пользу абсолютного преобладания последнего варианта [3,4,6].
Кроме того, на основании собственных результатов, данных отечественной и зарубежной литературы можно заключить, что лимитирующим моментом при реализации МАК является строго определенная функциональная значимость кристал- лизации в каждом конкретном случае [1-2, 4, 9-14]. С наших позиций, она и выступает в качестве ключевого звена при выборе вида расположения кристаллических структур, связанных с деятельностью микроорганизмов. В настоящее время мы предлагаем выделять следующие основные виды:
-
1. Экстрацеллюлярный – кристаллизация обнаруживается на определенном расстоянии от микроорганизма. В этом случае феномен кристаллизации связан с метаболической активностью последнего, т.е. синтезом и экзоцитозом определенных веществ во внешнюю для бактерии, вируса или гриба среду. К данному виду, в частности, относятся выявленные нами особенности кристаллообразования сыворотки крови при вирусных гепатитах В и С [4].
-
2. Примембранный – кристаллизационный процесс происходит в непосредственной связи с мембраной микробной клетки [1,7,12,15]. В данном случае, по нашему мнению, на первый план по отношению к метаболической активности микроорганизма выступают свойства самой бактериальной стенки (этот феномен в большей степени характерен для бактерий и грибов, для вирусов подобная активность к настоящему моменту не установлена). Это центр-инициатор кристаллизации, молекулярным субстратом которого может явиться липополисахарид клеточной стенки, а фактором, модулирующим сродство конкретного микроагента к формированию кристаллов определенного вещества строго детерминированных размеров и конфигурации – различные нелипидные компоненты бактериальной стенки, в частности, присутствующие в ней изобилии рецепторы и ионные каналы, имеющие белковую природу. Большинству биологических агентов свойственен именно этот тип микробной инициации кристаллогенеза.
-
3. Интрацеллюлярный – осуществляется путем инициации внутриклеточной кристаллизации микроорганизмом отдельных веществ. Чаще всего микроагент использует данный феномен в целях создания дополнительной защиты некоторых собственных структурных элементов. Примером подобного эффекта могут являться широко обсуждаемые в зарубежной литературе факты протектив-ной (для различных видов изучений) кристаллизации белковых молекул (в частности, белка RecA Escherichia Coli ) вокруг носителей генетического материала бактериальной клетки – макромолекул ДНК [13, 14]. Данный эффект обеспечивается за счет построения достаточно герметичных белковых «капсул» вокруг ДНК–нитей. Необходимость в этом возникает вследствие того, что, в отличие от более высокоорганизованных существ, бактерии не имеют собственной кариолеммы, и генетический материал прокариотов диффузно распределен по объему цитоплазмы.
В данной работе нами проанализированы результаты достаточно слабо изученного вида – экстрацеллюлярного МАК, причем полученные сведения о видоспецифичности и дозозависимости вари- антов проявления рассматриваемого явления позволяют рассчитывать на потенциальные возможности управления этим феноменом, что может быть полезно как с технических, так и с биомедицинских позиций.
Список литературы Биокристаллоскопические методы в изучении кристаллогенных свойств микроорганизмов
- Гусев М.В., Лебедева А.Ф., Саванина Я.В. и др. Устойчивость культур цианобактерии Anacystis nidulans и микроводоросли Dunaliella maritima к токсическому действию ванадия: влияние фосфата, железа и цистеина//Вестник МГУ. Сер. 16. 1997. № 2. С. 17-21.
- Дятлова К.Д. Микробные препараты в растениеводстве//Соросовский образовательный журн. 2001. Т. 7. № 5. С. 17-22.
- Камакин Н.Ф., Мартусевич А.К., Колеватых Е.П. О первичной и вторичной биокристаллизации//Естествознание и гуманизм (Томск). 2005. Т. 2. № 1. С. 18-19.
- Мартусевич А.К. Идеология изучения метаболической активности микроорганизмов кристаллографическими методами//Естествознание и гуманизм (Томск). 2007. Т. 4. № 2. С. 144.
- Мартусевич А.К., Камакин Н.Ф. Унифицированный алгоритм исследования свободного и инициированного кристаллогенеза биологических жидкостей//Клин. лаб. диагностика. 2007. № 6. С. 21-24.
- Мартусевич А.К., Зимин Ю.В. Роль физико-химических процессов в системе «микроорганизм-человек»//Вестник Санкт-Петербургской гос. мед. акад. им. И.И. Мечникова. 2008. № 3. С. 112-115.
- Сорокин Д.Ю. Совмещенные микробиохимические процессы трансформации неорганических веществ: роль в природных системах и возможности использования в биотехнологии//Микробиология. 1997. Т. 66. № 3. С. 293-301.
- Юшкин Н.П., Гаврилюк М.В., Голубев Е.А. Сингенез, взаимодействие и коэволюция живого и минерального миров: абиогенные и углеводородные кристаллы как модели протобиологических систем. Концепция кристаллизации жизни//Инф. бюлл. РФФИ. 1996. № 4. С. 393.
- Bozzi M., Mignogna G., Stefanini S. et al. A novel non-heme iron-binding ferritin related to the DNA-binding proteins of the Dps family in Listeria innocua//J. Biol. Chem. 1997. V. 272. P. 3259-3265.
- Ceci P., Ilari A., Falvo E. et al. The Dps protein of Agrobacterium tumefaciens does not bind to DNA but protects it toward oxidative cleavage: x-ray crystal structure, iron binding, and hydroxyl-radical scavenging properties//J. Biol. Chem. 2003. V. 278. P. 20319-20326.
- Ceci P., Mangiarotti L., Rivetti C. et al. The neutrophilactivating Dps protein of Helicobacter pylori, HP-NAP, adopts a mechanism different from Escherichia coli Dps to bind and condense DNA//Mol. Biol. 2007. № 3. P. 76-84.
- Minsky A., Shimoni E., Frenkiel-Krispin D. Biocrystallization: stress, order and survival//Nat. Rev. Mol. Cell Biol. 2002. № 3. P. 50-60.
- Namsaraev E., Baitin D., Bakhlanova I. et al. Biochemical basis of hyper-recombination activity of Pseudomonas aeruginosa RecA protein in Escherichia coli cells//Mol. Microbiol. 1998. V. 27. № 4. P. 727-738.
- Wolf S.G., Frenkiel D., Arad T. et al. DNA protection by stress-induced biocrystallization//Nature. 1999. V. 400. P. 83-85.
- Yong P., Rowson Neil A., Farr G.P.J. et al. Bioreduction and biocrystallization of palladium by Desulfovibrio desulfuricans NCIMB 8307//Biotechnol. and Bioengin. 2002. V. 80. № 4. P. 369-379.
