Биологические особенности триплоидной и тетраплоидной форм Gladiolus tenuis Bieb. при интродукции в Пермском крае

Автор: Черткова М.А., Шумихин С.А.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
Описаны биологические особенности триплоидной и тетраплоидной форм G. tenuis. На коллекционном участке Учебного ботанического сада ПГНИУ обе формы G. tenuis ежегодно цветут, однако все стадии сезонного развития у триплоидной формы наступают на 7-10 дней раньше, чем у тетраплоидной. Триплоидная и тетраплоидная формы по морфологии отличаются незначительно. Трип-лоидная форма G. tenuis в условиях интродукции в Пермском крае, как и в природе, плодов и семян не образует, размножается только вегетативно. Тетраплоидная форма G. tenuis при интродукции в Пермском крае способна размножаться как семенным путем, так и вегетативным. При этом свободное опыление оказывается результативнее, чем самоопыление. Успешность интродукции обеих изученных форм G. tenuis в Пермском крае оценивается нами как высокая. Обе формы G. tenuis могут быть использованы в озеленении, а тетраплоидная форма, кроме того, - для включения в селекционные программы в качестве источника декоративных и адаптивных признаков.
Триплоид, тетраплоид, интродукция, антэкология, жизнеспособность пыльцы, семенная продуктивность
Короткий адрес: https://sciup.org/147227013
IDR: 147227013 | УДК: 581.14:
Biological features of Gladiolus tenuis Bieb. triploid and tetraploid forms at introduction in Perm krai
This article describes the biological features of Gladiolus tenuis Bieb. triploid and tetraploid forms. Both forms of G. tenuis bloom annually in the Botanical Garden of PSU. But all stages of seasonal development in the triploid form occur 7-10 days earlier than in tetraploid. The morphologically triploid and tetraploid forms differ insignificantly. The triploid G. tenuis does not form fruits and seeds, it propagates only vegetatively. The tetraploid form of G. tenuis is capable of multiplying both by seeds and vegeta-tively in Perm Krai. In this case, free pollination is more effective than self-pollination. The authors rate the introduction of the studied G. tenuis forms as highly successful in Perm Krai. The studied forms of G. tenuis can be used in gardening, tetraploid form can be included in breeding programs as a source of decorative and adaptive features.
Текст научной статьи Биологические особенности триплоидной и тетраплоидной форм Gladiolus tenuis Bieb. при интродукции в Пермском крае
Геофиты, размножаясь в природе семенным путем, как правило, отличаются при этом хорошим потенциалом вегетативного возобновления. В то же время среди них известны случаи полной утраты полового способа размножения и формирования клонов, занимающих значительные части ареала [Kutlunina, Permyakova, Belyaev, 2016]. Причинами этого явления могут служить нечетная полиплоидия, анеуплоидия, сложная аллоплоидная природа, приводящие к стерильности.
Примером такого растения является Gladiolus tenuis Bieb. – гладиолус тонкий, охраняемый ред- кий вид, включенный в Красные книги 13 субъектов Российской Федерации. Ареал этого немораль-но-лесостепного вида приурочен преимущественно к Средней и Восточной Европе, Западной Сибири, Средней и Малой Азии, встречается он и на Кавказе [Тамберг, 1977]. Южный Урал и Оренбургская область фактически являются краевой частью ареала гладиолуса тонкого. По последним литературным данным к востоку от Волги все исследованные популяции G. tenuis относятся к одному стерильному генотипу (клону) и имеют триплоид-ный набор хромосом (2n=3x=45), к западу от Волги популяции тетраплоидные [Кутлунина, Князев, Беляев, 2010; Кутлунина, Князев, 2012; Князев,
Кутлунина, Беляев, 2015; Кутлунина, 2017].
Целью нашей работы было сравнительное изучение биологических особенностей тетраплоидной и триплоидной форм Gladiolus tenuis Bieb. в условиях интродукции в г. Перми.
Материалы и методы исследований
Исследования проводились на базе Учебного ботанического сада Пермского государственного национального исследовательского университета (ПГНИУ) в 2014–2017 гг. Обе формы G. tenuis выращиваются в Учебном ботаническом саду на интродукционном участке редких и охраняемых видов растений. Клубнелуковицы тетраплоидной формы поступили в коллекцию в 2014 г. из Ботанического сада Петра Великого (г. Санкт-Петербург), куда они попали из окрестностей г. Пятигорска. Клубнелуковицы триплоидной формы поступили в коллекцию Учебного ботанического сада ПГНИУ в 2000 г. из ФГБУН «Ботанический сад УрО РАН» (г. Екатеринбург); место их сбора – Губерлинские горы Оренбургской области.
Интродукционное изучение гладиолуса тонкого проводили по методическим рекомендациям, описанным И.Ф. Сацыперовой, А.М. Рабиновичем [1990] и Г.П. Семеновой [2007]. При этом описывали жизненную форму, морфологию растений, плодов, семян, фенологию, основной способ и особенности размножения. Наблюдения за ритмикой сезонного развития проводили по методике Р.А. Карписоновой [1972]. Антэкологические особенности изучали по методике, описанной А.Н. Пономаревым [1960]. Фертильность пыльцы определяли ацетокарминовым методом [Паушева, 1988]. Жизнеспособность пыльцы обеих форм G. tenuis определяли 2 способами: путем ее проращивания на искусственных средах ( in vitro ) и на рыльцах пестика ( in vivo ). Проращивание пыльцы in vitro проводили по методике И.Н. Голубинского [1974] на агаризованных средах, содержащих различные концентрации сахарозы (10, 15, 20, 25, 30, 35 и 40%). Жизнеспособность пыльцы in vivo и ре-цептивность рылец (их восприимчивость к пыльце) изучали на растениях триплоидной и тетраплоид-ной форм при их самоопылении и перекрестном опылении. Каждый вариант опыления проводили в цветках, находящихся в разных стадиях цветения, начиная с окрашенного бутона и до их увядания. Фиксацию и окрашивание пестиков проводили по методике Р.П. Барыкиной и др. [2004] . Срок наступления рецептивности рылец определяли по времени начала прорастания пыльцы после опыления.
Для изучения особенностей семенного размножения были поставлены 4 варианта опыта [Френкель, Галун, 1982; Шумихин, 2005]: 1) изоляция соцветий с кастрированными цветками без даль- нейшего вмешательства в процесс опыления, чтобы установить возможность апомиксиса; 2) изоляция соцветий без дальнейшего вмешательства в процесс опыления цветков, чтобы установить возможность естественного самоопыления; 3) изоляция соцветий с последующим искусственным самоопылением цветков, чтобы установить существование самонесовместимости и эффективность строгой автогамии; 4) учет завязывания семян при свободном опылении, чтобы определить его эффективность в конкретных условиях выращивания.
При изучении семенной продуктивности видов гладиолуса использовали стандартные методики [Вайнагий, 1974; Сацыперова, 1993]. При этом учитывали процент плодоцветения, потенциальную семенную продуктивность (ПСП), реальную семенную продуктивность (РСП), коэффициент семенной продуктивности (Кспр). При подведении итогов успешности интродукции использована 7балльная шкала оценки, разработанная в Донецком ботаническом саду [Баканова, 1984]. Статистическая обработка результатов проводилась по методике, описанной Г.Ф. Лакиным [1990].
Результаты и их обсуждение
Клубнелуковица у тетраплоидной формы G. tenuis шаровидная, 1.5–2 см в диаметре, крупнее, чем у триплоидной (1–1.5 см), покрытая волокнистыми чешуями. В условиях интродукции растения тетраплоидной формы в генеративной стадии развития в целом мельче триплоидной формы, достигают высоты 45–50 см по листьям и 53–70 см по генеративному побегу. Генеративные растения триплоидной формы достигают высоты в среднем 57–74 см по листьям и 70–80 см по генеративному побегу. Стебли у обеих форм прямые, тонкие. Нижние 2 листа чешуевидные, стеблевых листьев – 3. Они с развитой листовой пластинкой, мечевидные. Верхушечный лист 1, сильно уменьшенный. Цветки розово-фиолетовые, однако рисунок на нижних долях околоцветника тетраплоид-ной формы выражен четче и ярче (рис. 1, 2). В условиях Пермского края в соцветии тетраплоидной формы G. tenuis обычно формируется 5–9 цветков, триплоидной – 7–9. Цветки в соцветии триплоид-ной формы располагаются плотнее друг к другу. Околоцветник у обеих форм 3.0–3.5 см в диаметре. Цветок данного вида не имеет аромата. Плоды образуются только у тетраплоидной формы G. tenuis. Они представляют собой обратнояйцевидную коробочку до 10 мм длиной. Семена бескрылые, коричневые, овальные, 3–4 мм длиной.
По классификации жизненных форм декоративных геофитов, предложенной И.В. Борисовой [1991] с комментариями Л.Л. Седельниковой [2003], в условиях интродукции в Пермском крае G. tenuis отнесен нами к раннелетнецветущим гемиэфемероидом с ве- сенне-раннелетней вегетацией.
У G. tenuis цветки в соцветиях раскрываются в

Рис. 1. Тетраплоидная форма G. tenuis

Рис. 2. Триплоидная форма G. tenuis
В условиях Пермского края первой начинает отрастать триплоидная форма (24 апреля – 5 мая), затем в разные годы 30 апреля – 17 мая – тетрап-лоидная форма (рис. 3). Цветение триплоидной формы наступает также раньше, 5–19 июня. Тет-раплоидная форма G. tenuis зацветает в период 13 июня – 5 июля. Диссеминация у тетраплоидной формы отмечалась в разные годы с 10 июля по 3 августа. Усыхание листвы обеих форм отмечалось в разные годы в период 22 июля - 10 августа. Таким образом, за период исследования стадии сезонного развития у триплоидной формы наступали на 7–10 дней раньше, чем у тетраплоидной.
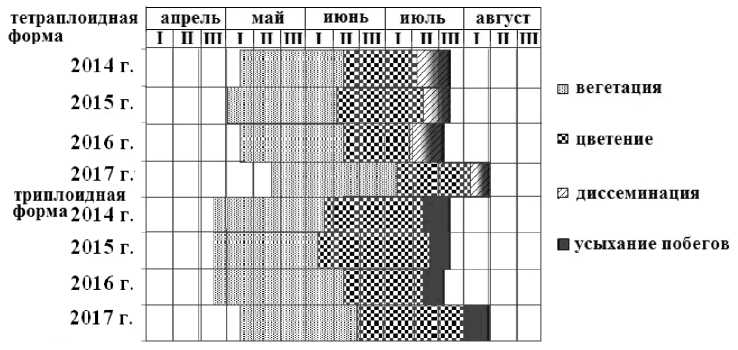
Рис. 3 . Феноспектр сезонного развития тетраплоидной и триплоидной форм G. tenuis в условиях интродукции в Пермском крае (2014–2017 гг.)
Изученные формы G. tenuis имеют пыльцу с высокими показателями фертильности. Однако фертильность пыльцы тетраплоидной формы (83.14±0.81%) достоверно превышает таковую триплоидной (76.23±1.42%; χ2= 8.39, р= 0.004).
При исследовании жизнеспособности пыльцы in vitro выяснено, что на искусственных средах прорастать способна только свежесобранная пыль- акропетальном направлении, от основания к вершине, с промежутком в один день. У обеих форм в соцветии не больше трех одновременно раскрытых цветков. От стадии окрашенного бутона до увядания цветка проходит 4–5 дней. Стадии раскрывания цветка описаны нами ранее [Черткова, Шумихин, 2015]. В целом они совпадают у обеих форм G. tenuis. Однако морфологическая готовность рыльца к опылению (рыльцевая стадия) фиксируется у тетраплоидной формы несколько раньше, через 3–7 ч после начала вскрывания пыльников, чем у триплоидной формы (через 3–9 ч). Наблюдения за ходом распускания цветка показали присутствие у обеих форм геркогамии и морфологически выраженной дихогамии в форме несовершенной протерандрии, которая не исключает самоопыление в форме автогамии или гейтоногамии. Автогамия возможна при неудавшемся перекрестном опылении на 4-й день функционирования цветка. Этому способствует, во-первых, отсутствие четкого разделения тычиночной и рыльцевой стадий из-за их наложения в течение 24–26 ч; во-вторых, изменение взаимного расположения пыльников и рылец в конце цветения, когда создаются условия для контактной автогамии. Гейтоногамия также вполне вероятна, поскольку существует вероятность попадания пыльцы с верхних цветков соцветия на готовые к опылению рыльца нижних.
ца тетраплоидной формы G. tenuis. Пыльца трип-лоидной формы на изученных искусственных средах не прорастала. Оптимальными для прорастания пыльцы тетраплоидной формы G. tenuis можно считать искусственные среды с содержанием сахарозы 10–15%. У тетраплоидной формы G. tenuis на оптимальных искусственных средах жизнеспособность пыльцы в целом невысока и составля- ет 23–29%. цы гладиолуса тонкого на рыльцах пестиков (in
Результаты изучения жизнеспособности пыль- vivo ) представлены в табл. 1.
Таблица 1
Жизнеспособность пыльцы тетраплоидной формы G. tenuis in vivo (%)
|
Вариант опыления |
День цветения цветка |
||||
|
1 |
2 |
3 |
4 |
5 |
|
|
Тетраплоидная форма × тетраплоидной форма |
0 |
18.88± 0.10 |
10.56± 0.45 |
3.79± 0.17 |
0 |
|
Триплоидная форма × тетраплоидная форма |
0 |
0 |
6.77± 1.63 |
6.17± 1.77 |
0 |
При самоопылении триплоидной формы пыльца не прорастает, являясь стерильной, что связано с нарушениями в процессах мейоза у форм с непарным числом хромосом в соматических клетках. При опылении триплоидной формы G. tenuis пыльцой тетраплоидной формы наблюдалось ее единичное прорастание (6.77±1.63% проросшей пыльцы). При самоопылении тетраплоидной формы наблюдалось в среднем 18.88±0.10% проросшей пыльцы .
При изоляции соцветий с кастрированными цветками без дальнейшего вмешательства в процесс опыления ни на одном растении за все годы исследования семена не завязывались. Таким образом, апомиксис у исследованных форм G. tenuis отсутствует. Апомиктичное завязывание семян в роде гладиолус не отмечается и в обзорных работах исследователей этого явления [Fryxell, 1957; Хохлов и др., 1978].
Семена у триплоидной формы при разных вариантах опыления и при свободном цветении не
Таблица 2
Показатели семенной продуктивности тетраплоидной формы G. tenuis при разных вариантах опыления
|
Вариант опыления |
Количество цветков |
Количество плодов |
Процент плодоцве-тения, % |
Показатели семенной продуктивности в расчете на плод |
К спр , % |
|||
|
ПСП (М±m) |
cv, % |
РСП (М±m) |
cv, % |
|||||
|
Естественное самоопыление |
26 |
9 |
34.62 |
33.95±0.59 |
2.48 |
4.94±1.87 |
53.54 |
14.56 |
|
Искусственное самоопыление |
42 |
20 |
47.62 |
34.28±1.55 |
10.12 |
8.18±2.20 |
60.17 |
23.86 |
|
Свободное опыление |
39 |
25 |
64.10 |
39.94±2.86 |
10.13 |
18.82±1.69 |
12.67 |
47.11 |
На коллекционном участке Учебного ботанического сада ПГНИУ обе формы G. tenuis ежегодно цветут, а тетраплоидная форма формирует плоды и семена. Отмечается хорошее вегетативное размножение обеих форм клубнелуковицами и клуб-непочками. Поражаемость изученных форм болезнями и вредителями низкая, а их зимостойкость оценивается нами как высокая, в течение всех лет исследования выпад растений не отмечался.
Заключение завязываются, что согласуется с литературными данными [Kutlunina, Permyakova, Belyaev, 2016]. Растения тетраплоидной формы способны формировать плоды и семена как при самоопылении, так и при свободном опылении, однако последнее оказывается наиболее результативным (табл. 2). Так, при самоопылении процент плодоцветения (34.62%) достоверно меньше (χ2= 5.44; р=0.02), чем при свободном опылении (64.10%). Коэффициент семенной продуктивности в обоих вариантах самоопыления (самоопылении при изоляции – 14.56% и при искусственном самоопылении – 23.86%) достоверно ниже, чем при свободном цветении – 47.11% (χ2= 82.00; р=0.000 и χ2=91.37; р=0.000 соответственно). Реальная семенная продуктивность в расчете на плод при свободном цветении (17.13±5.22) достоверно выше (t=[3.83; 5.51]> t05=1.96), чем при самоопылении при изоляции (4.94±1.87) и при искусственном самоопылении (8.18±2.20).
нелетнецветущими гемиэфемероидами с весеннераннелетней вегетацией. По результатам проведенных исследований успешность интродукции изученных форм G. tenuis в Пермском крае оценивается нами в 6 баллов, которые характеризуют высокую устойчивость к местным климатическим условиям. Изученные формы гладиолуса могут быть использованы в озеленении, а тетраплоидная форма – для включения в селекционные программы в качестве источника декоративных и ценных адаптивных признаков.
Таким образом, в условиях интродукции в Пермском крае обе формы G. tenuis являются ран-
Список литературы Биологические особенности триплоидной и тетраплоидной форм Gladiolus tenuis Bieb. при интродукции в Пермском крае
- Баканова В.В. Цветочно-декоративные многолетники открытого грунта. К.: Наук. думка, 1984. 156 с
- Барыкина Р.П. и др. Справочник по ботанической микротехнике. Основы и методы. М.: Изд-во МГУ, 2004. 312 с
- Борисова И.В. О понятиях «биоморфа», «экобио-морфа» и «архитектурная модель» // Ботанический журнал. 1991. Т. 76, № 10. С. 1360-1367
- Вайнагий И.В. О методике изучения семенной продуктивности растений // Ботанический журнал. 1974. Т. 59, № 6. С. 826-831.
- Голубинский И.Н. Биология прорастания пыльцы. Киев: Наук. думка, 1974. 368 с
- Карписонова Р.А. Методика фенологических наблюдений за травянистыми многолетниками в отделе флоры СССР // Методика фенологических наблюдений в ботанических садах СССР. М., 1972. С. 47-52
- Князев М.С., Кутлунина Н.А., Беляев А.Ю. Феномен особой устойчивости природных популяций клональных растений // Теоретические проблемы экологии и эволюции: шестые Лю-бищевские чтения. Тольятти, 2015. С. 167-170
- Кутлунина Н.А. Оценка генетического разнообразия триплоидных и тетраплоидных популяций Gladiolus tenuis (Iridaceae) методом ISSR-маркирования // Систематика и эволюционная морфология растений: материалы конф. М., 2017. С. 232-236
- Кутлунина Н.А., Князев М.С. Эволюционный потенциал клональных видов: миф или реальность? // Наука в России. 2012. № 4. C. 45-52
- Кутлунина Н.А., Князев М.С., Беляев А.Ю. Гигантский клон гладиолуса тонкого Gladiolus tenuis Bieb. (Iridaceae) в Уральской части ареала // Труды Института биоресурсов и прикладной экологии. Оренбург, 2010. Вып. 9. С. 7980
- Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990. 352 с
- Паушева З.П. Практикум по цитологии растений. М.: Колос, 1980. 304 с
- Пономарев А.Н. Изучение цветения и опыления растений // Полевая геоботаника, 1960. Т. 2. С. 9-19
- Сацыперова И.Ф. Основные аспекты и методы изучения репродуктивной биологии травянистых растений при их интродукции // Проблемы репродуктивной биологии семенных растений. СПб., 1993. Вып.8. С. 25-35
- Сацыперова И.Ф., Рабинович А.М. Проект общесоюзной программы исследований по интродукции лекарственных растений // Растительные ресурсы. 1990. Т. 26, вып. 4. С. 587-597
- Седельникова Л.Л. Биологические закономерности развития луковичных и клубнелуковичных геофитов при интродукции в лесостепную зону Западной Сибири: автореф. дис.... д-ра биол. наук. Новосибирск, 2004. 32 с
- Семенова Г.П. Редкие и исчезающие виды флоры Сибири: биология, охрана. Новосибирск: Гео, 2007. 408 с
- Тамберг Т.Г. Gladiolus L. - Гладиолус, или Шпажник // Декоративные травянистые растения для открытого грунта СССР: Класс Однодольных. Л., 1977. Т. 1. С. 177-198
- Френкель Р., Галун Э. Механизмы опыления, размножения и селекция растений. М.: Колос, 1982. 384 с
- Хохлов С.С., Зайцева М.И., Куприянов П.Г. Выявление апомиктичных форм во флоре цветковых растений СССР. Саратов: Изд-во СГУ, 1978. 224 с
- Шумихин С.А. Антэкологические исследования в селекции декоративных геофитов // Ботанические сады как центры сохранения биоразнообразия и рационального использования растительных ресурсов. Материалы междунар. конф. М., 2005. С. 554-556
- Черткова М.А., Шумихин С.А. К вопросу антэко-логии Gladiolus imbricatus L. и Gladiolus tenuis Bieb. в условиях Предуралья // Материалы II Междунар. науч. конф. «Лекарственные растения: фундаментальные и прикладные проблемы». Новосибирск, 2015. С. 44-46
- Fryxell P.A. Mode of reproduction of higher plants // Bot. Rev. 1957. Vol. 23, № 3, P. 135-233
- Kutlunina N., Permyakova M., Belyaev A. Genetic diversity and reproductive traits in triploid and tetraploid populations of Gladiolus tenuis (Iridaceae) // Plant Systematics and Evolution. 2016. Vol. 303, № 1. P. 1-10
