Биологические свойства низкогорных почв Южного Тимана

Автор: Виноградова Юлия Алексеевна, Лаптева Елена Морисовна, Хабибуллина Флюза Мубараковна, Тетерюк Людмила Владимировна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология и география почв
Статья в выпуске: 5 т.16, 2014 года.
Бесплатный доступ
В данной статье рассмотрены особенности структуры и состава микробиоты в почвах, формирующихся на выходах известняков в долинах рек Южного Тимана. Составлен систематический список почвенных микроскопических грибов, отмечено их невысокое видовое разнообразие при существенном вкладе грибного мицелия в структуру биомассы почвенных микроорганизмов.
Выходы известняков, карбо-литоземы, микробные сообщества, микромицеты, микробная биомасса
Короткий адрес: https://sciup.org/148203324
IDR: 148203324 | УДК: 631.461:631.44:551.432
Biological properties of low-mountain soils of the Southern Timan
This paper deals with structure and composition of microbiota in soils being formed on limestone outcrops in river valleys of the Southern Timan mountain range. We have classified soil microscopic fungi. They have a low species diversity degree. Fungal mycelium provides an essential contribution in biomass structure of soil microorganisms.
Текст научной статьи Биологические свойства низкогорных почв Южного Тимана
выходах кальцийсодержащих пород представлены разнообразные типы и подтипы почв, относящиеся к разным отделам: литоземы серогумусовые остаточно-карбонатные, литоземы перегнойно-темногумусовые остаточно-карбонатные, серогумусовые остаточно-карбонатные почвы, буроземы, а также почвы, классификационное положение которых на данный момент не вполне определено. Особенности биологических свойств карбонатных почв не исследовали.
Цель данной работы заключалась в изучении специфики состава и структуры комплекса микробиоты и видового разнообразия сообществ микроскопических грибов в почвах, формирующихся на выходах известняков в долинах рек Южного Тимана (на примере долины р. Омра).
ОБЪЕКТЫ И МЕТОДЫ
Район исследований расположен на территории Троицко-Печорского р-на Республики Коми, приурочен к среднетаежной подпровинции Ти-манской таежной провинции ВосточноЕвропейской таежной зоны [5]. Детальная характеристика района представлена в работах [3, 16]. Непосредственным объектом данных исследований послужили почвы, развитые на выходах известняков в долине среднего течения р. Омра – притока второго порядка р. Печора (62°44,3' с.ш., 55°53,2' в.д.). Долина реки в этом месте имеет V-образную форму поперечного сечения, ее ширина 50-70 м, глубина вреза реки 30-50 м.
Ключевые участки для проведения комплексных исследований выделены в пределах осыпного склона южной экспозиции. Крутизна склона около 35-45°. Ключевые участки приурочены к верхней части осыпного склона (I), средней части конуса осыпи (II) и нижней части склона – к зоне накопления элюво-делювия известняков (III). На каждом участке выполнено определение флористического состава фитоценозов [9, 4]. В период с 20 мая по 20 сентября 2004 г. проведено измерение температуры почвы с помощью температурных датчиков-логгеров (DS1921G, точность 0,5 С, интервал измерений 1,5 ч), установленных на поверхности почвы и в корнеобитаемом слое – на глубине 7-8 см от поверхности почвы. В августе 2011 г. заложены разрезы для морфологического описания почв и отбора проб в соответствии с генетическими горизонтами для физикохимических и микробиологических исследований.
Пробы почв для микробиологических исследований до начала исследований хранили при температуре минус 18-20°С. Суммарную численность бактерий и спор микроскопических грибов, длину мицелия, биомассу основных компонентов микробных сообществ определяли с использованием метода люминесцентной микроскопии [8]. Таксономическую принадлежность микромицетов идентифицировали после их выделения в чистые культуры с использованием определителей [21, 12], а также интерактивных «ключей» и информационного сайта интернет-ресурсов . Для характеристики структуры комплексов микромицетов использовали показатели частоты встречаемости. Для сравнения видового состава микромицетов применяли кластерный анализ (метод группировки по Варду, мера расстояния - Жаккар) [19].
Количественный химический анализ почв выполнен в аккредитованной лаборатории «Экоана-лит» и отделе почвоведения Института биологии Коми НЦ УрО РАН в соответствии с общепринятыми в почвоведении методами [14]. Величину рН водной вытяжки измеряли на иономере «Анион-4100» при соотношении почва : вода 1 : 2,5 для минеральных и 1 : 25 - органогенных горизонтов. Содержание общего органического углерода в органогенных горизонтах определяли на CHNS-О-элементном анализаторе ЕА 1110 (Италия), в минеральных - методом Тюрина со спектрофотометрическим окончанием, общего азота - на CHNS-О-анализаторе, обменных катионов - вытеснением КС1 с последующим атомноабсорбционным определением на «Хитачи 18060», гидролитическую кислотность - титриметрически по Каппену.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В высотном градиенте исследованного склона происходит закономерное преобразование состава растительности и, связанное с ним и с мощностью элюво-делювия, изменение почвенного покрова. В верхней части склона (участок I) древостой изрежен, представлен Betula pubescens и Picea obovata высотой до 2,0 м. В кустарниковом ярусе отмечены единичные экземпляры Salix sp. Травяно-кустарничковый ярус высотой 0,3 м имеет общее проективное покрытие около 60%. В нем доминирует Carex rhizina (cop3), с обилием cop2 встречаются Dryas punctata, Carex alba; cop1 - Thymus talijevii, Festuca ovina; sp - Carex ornithopoda, Dendranthema zawadskii, Euphrasia frigida, Aster alpinus, Campanula rotundifolia, Par- nassia palustris и др. виды. Моховолишайниковый ярус слабо развит (проективное покрытие 7-10%), образован в основном зелеными мхами (Hylocomium splendens, Rhytidiadelphus triqestrus) и лишайниками (Cladonia silvatica, Cladonia rongiferina, Cladonia stellaris).
Этот участок склона характеризуется максимальным уклоном - 35° и минимальной (20-25 см) мощностью мелкозема, перекрывающего плотные породы известняка. Активный боковой массоперенос, слабое развитие моховолишайникового покрова и обилие травянистых растений обусловили формирование в верхней части осыпного склона почв, морфологическое строение которых может быть выражено формулой: . В
0-6 6-15 15-20(25)
профиле почвы четко выделяется небольшой по мощности органогенный горизонт Оао, плотно переплетенный корнями травянистых растений. В его нижней части присутствует примесь минеральных частиц за счет активной жизнедеятельности почвенных беспозвоночных животных, верхняя часть представлена преимущественно остатками мхов, лишайников и травянистых растений. Под одернованным подстилочноторфяным горизонтом залегает хорошо структурированный, с обилием корней темногумусовый горизонт AU характерного темно-серого, близкого к черному, цвета. Он постепенно переходит в серовато-бурый с включением обломков карбонатов горизонт BCca, подстилаемый плитняком известняка. В системе классификации почв России [7] такие почвы относятся к отделу литоземов, типу карбо-литоземов темногумусовых. Данные химического анализа (табл. 1) подтверждают принадлежность почвы к типу карбо-литоземов темногумусовых. Диагностическими критериями темногумусового горизонта AU являются высокое содержание гумуса (5,6-6,4%), нейтральная реакция среды (7,67-7,72 ед. рН), высокое содержание обменных оснований (56,7-60,6 ммоль/100 г почвы).
Ключевой участок II располагается в средней части склона. Здесь крутизна склона сохраняется на уровне 35-40°. Однако мощность мелкозема, перекрывающего плитняк плотных пород, несколько больше - до 20-25 см. Соответственно растительный покров на этом участке представлен еловым редколесьем (8Е2Б) с примесью Betu-la pubescens и Pinus sylvestris. Высота древостоя 8-9 м, сомкнутость крон 0,1-0,2. Присутствует разреженный подрост ели высотой 0,5-1,0 м, кустарники - Juniperus communis, Spiraea media, Salix sp. В травяно-кустарничковом ярусе (общее проективное покрытие 80%) произрастают c обилием cop2 - Carex alba; cop1 - Carex ornithopoda, Carex rhizina, Arctostaphylos uva-ursi, Dryas punctata, Festuca ovina, Aster alpinus, Cypripedium cal- ceolus, Epipactis atrorubens, Vaccinium uliginosum, Vaccinium vitis-idaea; sp – Antennaria dioica, Equi-setum scirpoides, Gymnadenia conopsea, Gymnocar-pium robertianum, Pinguicula alpina, Polygala ama- rella. Мохово-лишайниковый ярус (проективное покрытие 60%) образован Hylocomium splendens, Pleurozium schreberi и др. зелеными мхами.
Таблица 1. Некоторые физико-химические свойства почв, сформированных на элюво-делювии карбонатных пород в долине р. Омра
|
Горизонт |
Глубина, см |
рНН2О |
Нг** |
Гумус, % |
Nобщ., % |
|
Р-16. Карбо-литозем темногумусовый, верхняя часть склона |
|||||
|
О |
0-4 |
7.58 |
н/о |
35.4* |
1.38 |
|
Оao |
4-6 |
7.84 |
н/о |
15.7* |
0.52 |
|
AU1 |
6-10 |
7.67 |
н/о |
6.4 |
0.27 |
|
AU2 |
10-15 |
7.72 |
н/о |
5.6 |
0.32 |
|
BCca |
15-20(25) |
7.93 |
н/о |
1.9 |
0.022 |
|
Р-17. Карбо-литозем темногумусовый, средняя часть склона |
|||||
|
СЖМ*** |
0+2 |
6.40 |
н/о |
44.6* |
1.06 |
|
O |
0-4 |
7.40 |
н/о |
26.2* |
0.96 |
|
AH |
4-8 |
7.73 |
н/о |
15.1* |
0.58 |
|
AU1 |
8-13 |
7.88 |
н/о |
8.7 |
0.43 |
|
AU2 |
13-18 |
8.03 |
н/о |
3.1 |
0.171 |
|
ВСca |
18-28(35) |
8.02 |
н/о |
2.8 |
0.23 |
|
Р-18. Серогумусовая остаточно-карбонатная почва, |
нижняя часть склона |
||||
|
СЖМ*** |
0+3 |
5.67 |
28.7 |
30.9* |
1.08 |
|
Oh |
0-3 |
6.09 |
8.8 |
20.8* |
1.01 |
|
H |
3-7 |
7.10 |
0.5 |
15.1* |
1.34 |
|
AY1 |
7-12 |
7.64 |
н/о |
4.2 |
0.24 |
|
AY2 |
12-17(22) |
7.80 |
н/о |
2.4 |
0.145 |
|
BCca |
17(22)-42 |
7.93 |
н/о |
0.9 |
0.071 |
Примечание.
веществ ммоль/100 г почвы; *** - слой живого мха.
* приведено содержание углерода органических
С/N
Обменные основания, ммоль/100 г почвы
|
Са2+ |
Mg2+ |
Сумма |
|
|
29.9 |
103.7 |
30.7 |
134.4 |
|
35.2 |
51.5 |
19.5 |
71.0 |
|
16.0 |
38.8 |
17.9 |
56.7 |
|
11.8 |
43.3 |
17.3 |
60.6 |
|
5.8 |
21.5 |
7.3 |
28.8 |
|
49.1 |
51.8 |
21.6 |
73.5 |
|
31.8 |
71.3 |
26.6 |
97.9 |
|
30.4 |
52.7 |
21.3 |
74.0 |
|
13.7 |
47.1 |
19.1 |
66.2 |
|
12.3 |
36.0 |
14.6 |
50.6 |
|
8.2 |
39.3 |
14.6 |
53.9 |
|
33.4 |
30.3 |
14.7 |
45.0 |
|
24.0 |
50.8 |
21.3 |
72.1 |
|
13.1 |
106.0 |
32.8 |
138.8 |
|
11.8 |
35.7 |
14.8 |
50.5 |
|
11.2 |
34.1 |
14.9 |
49.0 |
|
8.6 |
25.5 |
14.4 |
39.9 |
|
рг.); ** |
Нг – гидролитическая |
кислотность, |
|
,
На данном участке под пологом редкостойно- го елово-березового лишайниково- зеленомошного леса формируются почвы, профиль которых близок по строению к почве участка I: О — АН — AU^- ВСса-^^-Мса.
0-4 4-3 8-18 18-28(33)
Отличительной особенностью данной почвы является несколько большая мощность мелкозема (до 20-25 см) и органогенного горизонта (до 8 см). Нижняя часть подстилочно-торфяного горизонта (АН) переплетена корнями растений, струк- турирована, содержит примесь минеральных частиц и значительное количество копролитов, свидетельствующих об активной жизнедеятельности дождевых червей и других почвенных беспозвоночных животных. Темногумусовый горизонт АU, как и на участке I, характеризуется высоким содержанием гумуса (8,7-3,1%) и обменных оснований (66,2-50,6 ммоль/100 г. почвы). Реакция среды слабощелочная в верхней части профиля и щелочная – в нижней (табл. 1). В соответствии с особенностями строения и свойств почва средней части склона также диагностирована нами как карбо-литозем темногумусовый.
Ключевой участок III приурочен к нижней части склона (его подошве), где происходит ак- кумуляция элюво-делювия карбонатных пород. Крутизна склона в этой части около 45°. Древостой из Betula pubescens, Picea obovata, Pinus sylvestris. Высота деревьев достигает 13-14 м, сомкнутость крон до 0,5. Подрост из Alnus incana (высота 4-8 м) и Pinus sibirica. В подлеске Junipe-rus communis, Rosa acicularis, Lonicera pallasii, лиана (Atragene sibirica). В травянокустарничковом ярусе (общее проективное покрытие 60-70%, высота до 0,8-1,0 м) с обилием cop2 встречается Carex ornithopoda, cop1 – Den-dranthema zawadskii, Festuca ovina, Heracleum si-biricum, Milium effusum, Rubus saxatilis, Solidago virgaurea, Trifolium pretense, Trollius europaeus; Sp. – Angelica archangelica, Angelica sylvestris, Antennaria dioica, Cirsium heterophyllum, Galium boreale, Lathyrus vernus, Melampyrum sylvaticum, Melica nutans, Viola mauritii; sol – Aster alpinus, Campanula rotundifolia, Fragaria vesca, Polygala amarella. Мохово-лишайниковый ярус (проективное покрытие 80%) образован зелеными мхами Rhytidiadelphus triqestrus с примесью Dicranum sp.
В нижней части склона под пологом еловоберезового зеленомошного леса формируются почвы, имеющие мощность профиля около 40-45 см:
3 тт 4 10(15) 20(25) ,,
0/1--Н — AY \ , ВСса , \ у Мса. В
0-3 3-7 7-17(22) 17(22)-42
отличие от выше рассмотренных почв, в почве участка III нижняя часть подстилочно-торфяного горизонта имеет перегнойный характер, а гумусоаккумулятивный горизонт содержит меньшее количество гумуса (4,2-2,4%) и имеет серую окраску, что диагностирует наличие серогумусового горизонта AY, а не темногумусового AU, как в почвах верхней и средней частей склона. Более кислая среда подстилочно-торфяного горизонта (табл. 1), его несколько меньшая насыщенность основаниями (степень насыщенности основаниями в верхней части подстилки составила 89%, в отличие от нижнего перегнойного подгоризонта подстилки, где она достигает 100%), формирование серогумусового горизонта AY в минеральной части профиля и подстилание плотными породами на глубине ниже 30-35 см позволяют диагностировать почву участка III как серогумусовую остаточно-карбонатную грубогумусово перегнойную. Таким образом, в высотном градиенте осыпных склонов, приуроченных к выходам известняков в долине р. Омра, формируются почвы, относящиеся к двум отделам – отделу литозе-мов (карбо-литоземы темногумусовые) и отделу органо-аккумулятивных почв (серогумусовые остаточно-карбонатные почвы).
Ранее на выходах известняков в бассейне р. Сойва, притоком которой является р. Омра, были прослежены широтные закономерности и динамика поступления прямой солнечной радиации на разноориентрованные склоны, перераспределения температур с учетом мезорельефа модельных склонов разной экспозиции [3, 16]. Установлено, что растительные сообщества на южных склонах получают больше тепла, произрастают при более стабильном температурном режиме, защищены от воздействия поздних весенних и ранних осенних заморозков.
Наблюдения за температурным режимом почв на выделенных ключевых участках в долине р. Омра показали, что в рассмотренном ряду почв наиболее теплообеспечены почвы средней части склона (табл. 2). Почвы верхней и нижней частей склона по суммам среднесуточных температур близки и характеризуются меньшими величинами, по сравнению с почвой средней части склона. Расчет сумм суточных перепадов температуры свидетельствует, что почва средней части склона отличается относительно стабильным температурным режимом. Более выраженные суточные перепады температур почвы в верхней части склона могут быть обусловлены разреженностью древостоя, а в нижней части склона – влиянием русла реки и подтоплением в период паводков.
Таблица 2. Температурный режим почв, формирующихся на выходах известняков в долине р. Омра (по данным за период с 20.05.04 по 20.10.04)
|
Показатели |
Ключевой участок |
||
|
верхняя часть склона (III) |
средняя часть склона (II) |
нижняя часть склона (I) |
|
|
Сумма среднесуточных температур: выше 0 С |
1615.8 |
1692.5 (1573.4*) |
1595. 8 |
|
выше 5 С |
1 584.6 |
1 674.7 (1568.5*) |
1 578.5 |
|
выше 10 С |
1368.2 |
1433.9 (1307.9*) |
1355.6 |
|
Сумма максимальных суточных температур, °С |
2636,5 |
2601 (1956,5*) |
2835,5 |
|
Сумма минимальных суточных температур, °С |
909 |
1088,5 (1304*) |
890,5 |
|
Сумма суточных перепадов температуры, °С |
1723,5 |
1509,5 (651,5*) |
1941 |
|
Число дней со среднесуточной температурой: выше 5 С |
113 |
117 (120*) |
117 |
|
выше 10 С |
85 |
87 (87*) |
88 |
|
Длительность безморозного периода |
93 |
110 |
92 |
* - показатели для корнеобитаемого слоя почвы (глубина 7-8 см).
Различия в экологических условиях формирования карбонатных почв в пределах склоновых поверхностей долины р. Омра нашли свое отражения в особенностях структуры и состава их микробных комплексов (табл. 3). Общей закономерностью для всех рассмотренных почв является концентрирование микроорганизмов в органогенных горизонтах. В нижележащих минеральных горизонтах численность бактерий снижается в 532, спор грибов – в 2-8 раз, длины мицелия гри- бов – в 2-12 раз, что в принципе характерно для профильного распределения микроорганизмов в почвах таежных экосистем [11]. В структуре биомассы микробных сообществ основную роль играет мицелий микроскопических грибов. На его долю во всех почвах приходится 92-96% общей биомассы микроорганизмов. Такая картина типична для почв таежных лесов [13].
Следует отметить, что рассмотренные почвы существенно отличаются друг от друга по чис- ленности и биомассе отдельных структурных компонентов блока почвенной микробиоты. Как видно из представленных данных (табл. 3), почвы верхней и нижней частей склона, характеризуются меньшими параметрами численности и биомассы прокариот, по сравнению с почвой средней части склона, для которой отмечены меньшие суточные перепады температуры, более длительный безморозный период и лучшая теплообеспе-ченность (табл. 2). В органогенном горизонте карбо-литозема темногумусового, занимающего среднюю часть склона, численность бактерий составляет 177-543 млн. кл./г почвы, их максимум приходится на верхнюю часть подстилочноторфяного горизонта, минимум – на нижнюю, перегнойную часть лесной подстилки. В слое живых мхов численность бактерий ниже, чем в верхней части органогенного горизонта. В карбо-литоземе темногумусовом, представленном в верхней части склона, органогенный горизонт достаточно однороден по содержанию прокариот – во всех его подгоризонтах численность бактерий находится на уровне 86-89 млн. кл./г почвы. В серогумусовой остаточно-карбонатной почве нижней части склона численность прокариот варьирует в пределах органогенного горизонта от 42 (в перегнойном подгоризонте) до 192 (в верхней части лесной подстилки) млн. кл./г почвы. Минеральные горизонты почв I и III участков, несмотря на различия в содержании органического углерода, идентичны по содержанию прокариот, и в 3-8 раз их численность меньше по сравнению с аналогичными горизонтами почвы средней части склона. Биомасса прокариот отражает закономерности, выявленные для профильного распределения численности бактерий в почвах осыпного склона.
Таблица 3. Численность и биомасса микроорганизмов в почвах, формирующихся на выходах известняков в долине р. Омра (X + 4)*
|
Горизонт |
Глубина, см |
Численность |
Длина мицелия грибов, м/г |
Биомасса, мкг/г |
|||
|
бактерии, млн кл./г |
споры грибов, млн кл./г |
бактерий |
мицелия грибов |
спор грибов |
|||
|
Р-16. Карбо-литозем темногумусовый, верхняя часть склона |
|||||||
|
О |
0-4 |
89±22 |
2.7±0.5 |
107.0±5.4 |
1.78±0.22 |
417.3±10.5 |
27.3±2.5 |
|
Оao |
4-6 |
86±6 |
2.2±0.9 |
73.5±11.3 |
1.71±0.06 |
286.6±22.0 |
22.3±4.3 |
|
AU1 |
6-10 |
14±5 |
0.71±0.01 |
38.7±4.3 |
0.28±0.05 |
150.8±8.3 |
7.12±0.04 |
|
AU2 |
10-15 |
7±7 |
0.94±0.14 |
30.9±2.8 |
0.15±0.07 |
120.5±5.4 |
9.4±2.1 |
|
BCca |
15-20 |
9±1 |
0.52±0.09 |
18.3±9.6 |
0.17±0.03 |
71.3±18.7 |
5.0±1.3 |
|
Р-17. Карбо-литозем темногумусовый, средняя часть склона |
|||||||
|
СЖМ** |
0+2 |
344±3 |
5.4±1.9 |
195.9±133.2 |
6.87±0.03 |
764.1±259.7 |
53.7±9.4 |
|
O |
0-4 |
543±23 |
7.1±0.7 |
237.8±43.8 |
10.86±0.23 |
927.3±85.5 |
71.4±3.3 |
|
AH |
4-8 |
177±29 |
2.1±0.2 |
66.4±7.2 |
3.55±0.29 |
259.0±14.0 |
21.0±1.0 |
|
AU1 |
8-13 |
42±15 |
0.89±0.12 |
41.6±5.6 |
0.84±0.15 |
130.0±9.2 |
9.0±0.6 |
|
AU2 |
13-18 |
49±23 |
0.91±0.14 |
- |
0.98±0.23 |
- |
9.12±0.7 |
|
ВСca |
18-28(35) |
36±3 |
0.94±0.24 |
- |
0.72±0.03 |
- |
9.37±1.21 |
|
Р-18. Серогумусовая остаточно-карбонатная почва, нижняя часть склона |
|||||||
|
СЖМ** |
0+3 |
89±6 |
3.0±0.1 |
220.1±19.8 |
1.79±0.06 |
90.0±6.4 |
29.8±06 |
|
Oh |
0-3 |
192±3 |
4.9±0.3 |
333.8±56.7 |
3.84±0.03 |
1301.9±110.5 |
48.9±1.6 |
|
H |
3-7 |
42±4 |
1.26±0.02 |
50.9±9.3 |
0.84±0.04 |
198.4±18.1 |
12.6±0.1 |
|
AY1 |
7-12 |
13±1 |
0.61±0.06 |
28.9±7.8 |
0.25±0.01 |
113.1±15.2 |
6.1±0.3 |
|
AY2 |
12-17(22) |
6±1 |
0.68±0.06 |
27.2±6.3 |
0.12±0.01 |
110.0±12.4 |
6.8±0.3 |
|
BCca |
17(22)-42 |
- |
0.76±0.02 |
23.0±2.4 |
- |
89.6±4.7 |
7.6±0.1 |
Примечание. * A - среднее арифметическое; Δ – стандартное отклонение; ** - слой живого мха. Знаком « » отмечено отсутствие показателя.
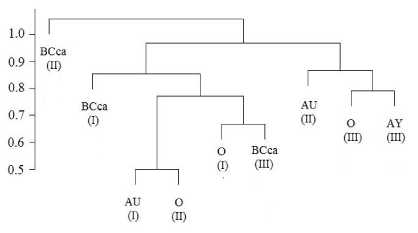
Рис. Дендрограмма сходства видового состава микроскопических грибов в почвах, формирующихся на выходах известняков в долине р. Омра (кластеризация – по Варду, мера расстояния – Жаккар).
Для мицелия почвенных микроскопических грибов отмечено увеличение его длины и биомассы в направлении от верхней части склона (I) к его подошве (III), что связано с некоторым возрастанием в этом направлении кислотности почв. В целом в подстилочно-торфяных горизонтах длина мицелия варьирует в пределах от 50,9 до 333,8 м/г почвы, а его биомасса – от 198 до 1302 мкг/г почвы. Максимальными показателями отличаются верхние подгоризонты лесных подстилок, где идут первые этапы трансформации растительных остатков с участием гидролитических ферментов грибов, минимальными – нижние, перегнойные. В минеральных горизонтах длина мицелия и его биомасса снижаются на порядок по сравнению с верхними органогенными горизонтами. Следует обратить внимание на тот факт, что в почве средней части склона, наиболее теплообеспеченной и характеризующейся щелочной реакцией среды, в горизонтах на глубине ниже 15 см, где величина рН водных вытяжек превысила 8 единиц, грибной мицелий отсутствует. В структуре микробных комплексов в этих горизонтах были отмечены только клетки прокариот и споры грибов. Учитывая, что микроскопические грибы приурочены преимущественно к почвам с кислой реакцией среды [15], отсутствие мицелия грибов в нижних горизонтах минеральной части профиля почвы средней части склона, отличающихся щелочной реакцией среды, вполне закономерно.
Изучение видового разнообразия микроскопических грибов показало, что в совокупности в рассмотренных почвах комплекс микромицетов представлен 39 видами, относящимися к 16 родам, двум отделам Zygomycota, Ascomycota и неформальному классу анаморфных грибов (включая два «вида» стерильных форм мицелия). Максимальной видовой насыщенностью характеризуются роды Penicillium (10 видов), Mortierella (4) , Paecilomyces (4) , Trichoderma (3). Низкое видовое разнообразие микромицетных комплексов, а также основного кислотообразующего рода Penicillium , обусловлено, прежде всего, нейтральной реакцией среды почв, формирующихся на элюво-делювии известняков. Для сравнения, в кислых подзолистых почвах, развитых на водоразделах в подзоне средней тайги на территории Республики Коми, таксономический список микроскопических грибов включает до 58 видов, при этом до 60% видового состава микромицетных комплексов представлено видами рода Penicillium [17]. Наиболее богата по видовому разнообразию микромицетов почва нижней части склона. Здесь выявлено 29 видов микроскопических грибов. Минимальным числом видов микромицетов характеризуется почва средней части склона – 17 видов. Почва верхней части склона занимает промежуточное положение по видовому разнообразию микроскопических грибов – 20 видов.
Следует отметить, что микромицетные комплексы органогенных горизонтов, где аккумулируется основная масса растительного опада, отличаются максимальным разнообразием микроскопических грибов, с преимущественной их концентрацией в почве нижней (16 видов) и верхней (15 видов) частей склона. В органогенном горизонте почвы средней части склона, представленном остатками зеленых мхов, листьями толокнянки, дриады и других видов трав, видовое разнообразие микромицетов снижено до 7-9 видов. В нижних минеральных горизонтах исследованных типов почв присутствует от 2 до 9 видов микро-мицетов. Более высокое разнообразие микоценозов подстилочно-торфяных горизонтов и их резкое снижение по мере перехода к минеральным горизонтам, обусловлено как снижением общего содержания органического вещества, так и кислотности среды, что связано близким залеганием карбонатных пород (табл. 2). Аналогичные закономерности распределения микромицетов по профилю выявлены при исследовании карбонатных почв Северо-Западного Кавказа [6].
Для карбонатных почв Южного Тимана характерны такие виды, как Catenularia pidopliczkoi, Zygodesmus marginatus, Hormodendrum hordei. Перечисленные виды можно рассматривать в качестве видоспецифичных для карбонатных почв, т.к. они не были встречены нами в почвах равнинных территорий Республики Коми, формирующихся в различных экологических условиях как зональных, так и интразональных ландшафтов [17. 18]. Из наиболее часто встречающихся видов рода Penicillium в карбонатных почвах, развитых на выходах известняков в долине р. Ом-ра, следует отметить Penicillium thomii и P. сyclopium , из видов рода Mortierella – M . alpine и M. verticillata, из рода Cladosporium – С. сladosporioides , рода Geomyces – G . pannorum. Специфичными для почвы средней части осыпного склона являются виды рода Paecilomyces ( P. varioti, P. farinosus, P. carneus ), нижней части склона – виды рода Monocillium ( M. humicola, M. exsolum ).
Статистическая обработка данных по видовому составу микроскопических грибов с использованием кластерного анализа выявила определенную специфичность микоценозов рассмотренных нами почв (рис.), обусловленную (1) своеобразием формирования почв на склоновых поверхностях, сопровождающихся боковым массоперено-сом вещества вниз по склону; (2) влиянием кар-бонатности почвообразующих пород (элюво-делювия известняков); (3) особенностями температурного режима почв и (4) характером растительного покрова. Оказались наиболее близки по видовому составу (образовали единый кластер) верхние горизонты (О и AY) почвы нижней части склона, формирующейся под пологом елово- березового зеленомошного леса, наиболее кислой в рассматриваемом ряду почв. Как видно из рисунка, к ним тяготеет гумусоаккумулятивный горизонт (AU) почвы склона, что может быть обусловлено в какой-то мере возможным перемещением грибных пропагул с внутрипочвенным стоком и их аккумуляцией в нижней части склона. Переходный горизонт BCca почвы средней части склона занимает обособленное положение, что связано с щелочной реакцией среды этого горизонта и возможным существованием здесь строго определенных видов микроскопических грибов. Все остальные горизонты карбо-литоземов, формирующихся в верхней и средней частях склона, включая нижний минеральный горизонт BCca почвы нижней части склона, образуют второй кластер.
ВЫВОДЫ
-
1) Установлено, что в пределах южных склонов на элюво-делювии карбонатных пород в долине р. Омра, формируются почвы, относящиеся к карбо-литоземам темногумусовым (верхние и средние части осыпных склонов) и серогумусовым остаточно-карбонатным почвам (нижняя часть склонов);
-
2) В структуре микробных комплексов почв, формирующихся на выходах известняков, как и в почвах таежных экосистем, развитых на бескар-бонатных породах, ведущую роль играет мицелий микроскопических грибов (92-96% общей микробной биомассы). Количественные параметры и глубина распространения мицелия грибов определяются кислотно-основными условиями среды: при величине рН водных вытяжек более 8 ед. рН микроскопические грибы присутствуют только в виде спор;
-
3) Видовое разнообразие микромицетных комплексов в почвах, формирующихся на элюво-делювии известняков, существенно ниже по сравнению с зональными подзолистыми почвами;
-
4) На параметры численности и биомассы прокариот в карбонатных почвах решающее влияние оказывает не тип почвы, а условия теп-лообеспеченности.
Работа выполнена при финансовой поддержке проекта программы Президиума РАН 12-Т-4-1006 « Экологические качества эталонных почв Европейского Северо-Востока России, их биоор-ганический потенциал как критерий продуктивности и охраны в свете подготовки Красной книги почв Республики Коми».
Список литературы Биологические свойства низкогорных почв Южного Тимана
- Атлас почв Республики Коми. Сыктывкар, 2010. 356 с.
- Добровольский Г.В. Сохранение почв как незаменимого компонента биосферы/Г.В. Добровольский, Е.Д. Никитин. М.: Наука, МАИК «Наука/Интерпериодика», 2000. 185 с.
- Елсаков В.В. Роль рельефа в формировании растительности карстовых ландшафтов европейского северо-востока России/В.В. Елсаков, Л.В. Тетерюк//Исследование Земли из космоса. 2012. № 3. С. 78-93.
- Ипатов В.С. Описание фитоценоза. Методические рекомендации. СПб., 1998. 93 с.
- Исаченко А.Г. Экологическая география Северо-Запада России. СПб.: Изв. РГО, 1995. Ч. 1. 206 с.
- Казеев К.Ш. Влияние карбонатности пород на биологические свойства горных почв северо-западного Кавказа/К.Ш. Казеев, М.А. Кутровский, Е.В. Даденко, Л.С. Везденеева, С.И. Колесников, В.Ф. Вальков//Почвоведение. 2012. № 3. С. 327-335.
- Классификация и диагностика почв России/Составители: Л.Л. Шишов, В.Д. Тонконогов, И.И. Лебедева, М.И. Герасимова. Смоленск: Ойкумена, 2004. 342 с.
- Методы почвенной микробиологии и биохимии/Под. ред. Д.Г. Звягинцева. М.: Изд-во МГУ, 1991. 304 с.
- Нешатаев Ю.Н. Методы анализа геоботанических материалов. Л., 1987. 192 с.
- Почвы и почвенный покров Печоро-Илычского заповедника (Северный Урал)/Отв. ред. С.В. Дегтева, Е.М. Лаптева. Сыктывкар, 2013. 328 с.
- Регуляторная роль почвы в функционировании таежных экосистем/Отв. ред. Г.В. Добровольский. Москва: Изд-во Наука, 2002. 364 с.
- Саттон Д. Определитель патогенных и условно патогенных грибов/Д. Саттон, А. Фотергиль, М. Ринальди. М.: Мир, 2001. 486 с.
- Стольникова Е.В. Микробная биомасса, ее активность и структура в почвах старовозрастных лесов Европейской территории России/Е.В. Стольникова, Н.Д. Ананьева, О.В. Чернова//Почвоведение. 2001 № 4. С. 479-494.
- Теория и практика химического анализа почв/Под ред. Л.А. Воробьевой. М.: Изд-во МГУ, 2006. 400 с.
- Терехова В.А. Микромицеты в экологической оценке водных и наземных экосистем. М.: Наука, 2007. 215 с.
- Тетерюк Л.В. Роль рельефа в формировании терморежима и биоразнообразия реликтовых экосистем на известняках европейского северо-востока России/Л.В. Тетерюк, В.В. Елсаков, Е.М. Лаптева//Экология. 2012. № 6. С. 410-417.
- Хабибуллина Ф.М. Почвенная микобиота естественных и антропогенно нарушенных экосистем Северо-Востока европейской части России: Автореф. дис. докт. биол. наук. Сыктывкар, 2009. 40 с.
- Хабибуллина, Ф.М. Разнообразие микромицетов в почвах пойменных лугов/Ф.М. Хабибуллина, Е.М. Лаптева, Ю.А. Виноградова//Микология и фитопатология. 2009. Т. 43, Вып. 3. С. 200-206.
- Шипунов, А.Б. Наглядная статистика. Используем R!/А.Б. Шипунов, E.М. Балдин, П.А. Волкова, А.И. Коробейников, С.А. Назарова, С.В. Петров, В.Г. Суфиянов. М.: ДМК Пресс, 2012. 298 с.
- Юдин Ю.П. Реликтовая флора известняков Северо-Востока европейской части СССР//Материалы по истории флоры и растительности СССР. М., 1963. Вып. 4. С. 493-587.
- Ramirez C. Manual and atlas of the Penicillia. Amsterdam; New York; Oxford: Elsevier Biomedical Press, 1982. 874 p.
