Биология цветения Rhododendron japonicum и R. viscosum при интродукции на юге Приморского края

Автор: Кокшеева И.М., Творогов С.П., Нехайченко Д.В.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 5, 2018 года.
Бесплатный доступ
В статье приведены данные о структурных и функциональных особенностях генеративной сфе-ры восточноазиатского вида Rhododendron japoni-cum (A. Gray) Suring. и североамериканского Rh. viscosum (L.) Torr. при интродукции в Приморском крае (г. Владивосток). Исследованы морфологиче-ские особенности цветка, типы опыления и су-точная ритмика распускания цветков. Наличие соцветий, крупных и ярко окрашенных цветков, присутствие аромата, - все это подтверждает принадлежность видов рода Rhododendron к энто-мофилии. Отражены результаты принудительно-го само- и перекрестного опыления. Показано, что перекрестное опыление в пределах вида способ-ствовало формированию более качественных се-мян, чем при опылении в пределах одного расте-ния. Определены сроки распускания цветков и про-должительность цветения видов рода Rhododen-dron. Начало цветения Rh. japonicum и Rh. viscosum отмечалось в конце мая, массовое - на 7-10-й день, и завершалось в среднем на 24-28-й день. Выявлено два типа суточной ритмики распускания цветков: утренний (Rh...
Биология цветения, опыление, циркадные ритмы
Короткий адрес: https://sciup.org/140238109
IDR: 140238109 | УДК: 582.912.42(571.63)
Flowering biology of Rhododendron japonicum and R.viscosum at their introduction in the south of Rrimorsky region
The data on structural and functional features of generative sphere of East Asian type of Rhododendron japonicum (A. Gray) of Suring and North American Rh. viscosum (L.) Torr. are provided in the study at an intro-duction in Primorsky Region (Vladivostok). Morphologi-cal features of the flower, types of pollination and daily rhythmics of blooming of flowers are investigated. The availability of inflorescences, large and brightly colored flowers, the presence of aroma, all this confirms the belonging species of the genus Rhododendron to ento-mophilia. The results of forced self- and cross-pollination are reflected. It is shown that cross-pollination within species contributes to the formation of more qualitative seeds, than at pollination within a single plant. The terms of blooming of flowers and duration of blossoming of species of the genus Rhododendron are determined. The beginning of blossoming of Rh. japoni-cum and Rh. viscosum was noted at the end of May, massive blossoming was on the 7th-10th day and ended on the average on 24-28th...
Текст научной статьи Биология цветения Rhododendron japonicum и R. viscosum при интродукции на юге Приморского края
Введение . Рододендроны – одна из перспективных групп растений, пользующихся популярностью во всем мире. При интродукции в условиях юга Приморского края особый интерес представляют рододендрон японский Rh. japonicum (Gray) Suring. и рододендрон клейкий Rh. viscosum (L.) Torr. как зимостойкие и высоко декоративные виды. Представитель флоры Японии (о-ва Хонсю, Сикоку, север Кюсю) – Rh. japonicum широко известен в озеленении своими крупными и яркими цветками. В природе он произрастает на солнечных травянистых склонах гор или среди высоких кустарников на высоте 100–2000 м над уровнем моря. В Ботаническом саде-институте ДВО РАН г. Владивосток (БСИ ДВО РАН) вид впервые введен в культуру в 1979 г. из Прибалтики [1]. В настоящее время культивируется более 50 экземпляров растений различного возраста. Rhododendron viscosum – представитель североамериканской флоры произрастает вдоль тихоокеанского побережья от штата Мэн до Южной Каролины. Места его обитания – болота, прибрежные равнины, горы, влажные леса и сырые просеки. Первые экземпляры получены в коллекции БСИ ДВО РАН в 1989 г. из Ботанического сада Киевского национального университета имени Тараса Шевченко (г. Киев). В настоящее время в коллекции присутствует множество экземпляров собственной репродукции.
Семенное размножение для рододендронов является наиболее доступным способом получения посадочного материала. В условиях интродукции исследуемые виды формируют жизнеспособную пыльцу, однако регулярное плодоношение отмечено только у Rh. japonicum , семенная продуктивность у Rh. viscosum в условиях интродукции низкая [2–4].
Известно, что цветение является критическим моментом в цикле полового размножения и семенная продуктивность зависит от биологических особенностей цветения, климатических условий в этот период и от опыления. Вопросы биологии цветения рододендронов в литературе освещены недостаточно. Известна работа Е.А. Тихменева по экологии цветения и опыления Rh. aureum Georgi и Rh. redowskianum Maxim. на севере Дальнего Востока [5], где автор выявил два типа суточной ритмики распускания цветков и их корреляцию с температурой. В условиях Львова вопросами экологии и суточной ритмики цветения Rh. japonicum занимался Р.В. Карамзин [6].
Адаптация растений – интродуцентов к новым условиям часто выражается в изменении сезонного ритма развития, в частности цветения. Многие декоративные растения демонстрируют суточный ритм открытия и закрытия цветка [7–9]. Открытие цветов днем и закрытие ночью имеют некоторые преимущества для растений, поскольку многие опылители активны в течение дня, обеспечивая успешное опыление, оплодотворение и формирование плодов. Кроме того, изменениям подвергаются и взаимоотношения интродуцентов с агентами опыления, состав которых в районе интродукции может быть иным.
Цель исследования : выявление структурных и функциональных особенностей генеративной сферы Rh. japonicum и Rh. viscosum в условиях юга Приморского края. В частности, исследование направлено на изучение морфологических особенностей цветка, выявление типов опыления и определение суточной ритмики распускания цветков.
Материал и методы исследования. Исследование выполнено в Ботаническом саде-институте ДВО РАН (г. Владивосток), где Rh. japonicum и Rh. viscosum произрастают в открытом грунте. Для определения типов опыления проведен ряд экспериментов: 1 – изолирование соцветия; 2 – проведение перекрестного опыления в пределах одного растения; 3 – проведение перекрестного опыления между различными растениями одного вида. Опыление осуществляли с кастрацией венчика, пыльников и изоляцией накануне раскрытия цветков. Изоляцию в виде конвертов из Агротекса использовали на весь период цветения растений. Для опыления использовали по 5 цветков в каждом соцветии в трехкратной повторности в течение двух сезонов. Далее проводили подсчет завязавшихся плодов и определяли в них количество семян. Динамику распускания цветков изучали на пяти модельных кустах каждого вида. Динамика распускания цветков: раскрытие венчика, пыльников, готовность пестиков к восприятию и т. д. На протяжении всего периода цветения ежедневно с 600 до 2000 вели учет суточной ритмики распускания цветков, каждые 2 часа подсчитывали число распустившихся цветков, после этого цветки удаляли. Данные обрабатывались с помощью программы Excel. В статье использовали общепринятую терминологию [5, 10–12]. Под суточной ритмикой распускания цветков подразумевали количество цветков, распускающихся в течение суток. Для определения типа суточной ритмики (утренний, дневной, вечерний и ночной) принимали во внимание время распускания цветков, а не период суток, в течение которого они бывают открыты.
Результаты исследования
Особенности строения цветка и типы опыления
Цветки Rh. japonicum собраны в соцветие, которое характеризуется как видоизмененная или укороченная кисть, т. е. цветки имеют цветоножки равной длины и расположены на укороченной главной оси. Всего в соцветии наблюдали от 2 до 16 цветков с легким ароматом, и их число зависело от климатических условий в период формирования цветочной почки или в зимний период. Венчик сростнолепестный, широко воронковидный, 6–7,7 см в диаметре, от ярко-алого, лососевого до светло-оранжевого с коричневым или зеленым крапом. Кроме того, есть формы, окраска венчика которых варьирует от светлого до насыщенно желтого цвета. Тычинки изогнутые вовнутрь, длиной 4,1–5 см по 5 шт. Пестик немного длиннее тычинок – 5,4–6 см.
Цветки Rh. viscosum с легким ароматом, по 7–15 шт. собраны в кисть. Венчик сростнолепестный, воронковидный, 3,8–4,6 см в диаметре, от светло- до темно-розового цвета, трубка венчика длиной 1,4– 1,8 см. Тычинки изогнуты вовнутрь, длиной 4,4–5 см по 5 шт., пестик – 5,3–5,6 см.
Особенности строения цветков: крупные, ярко окрашенные цветки в соцветиях, наличие ярких пятен, указателей нектара на лепестках, присутствие аромата, – говорят об их принадлежности к энтомофилии. В условиях юга Приморского края цветки рододендронов посещали пчелы, шмели, осы, мухи-журчалки и т. д. Насекомые, посещая цветок, добывали нектар, ориентируясь на указатели нектара, погружались в глубь цветка, однако некоторые предпочитали прокалывать цветок снаружи, в месте нахождения нектара. Стоит отметить, что насекомые при посещении цветка не касались пыльников, поскольку тычиночные нити и пыльники выходят далеко за пределы венчика. Однако они касались тычиночных нитей, способствуя высыпанию пыльцы из пыльников и перекрестному опылению (аллогамии). Аллогамия у рододендронов осуществлялась при помощи абиотических (ветер) и биотических агентов (насекомые). Кроме перекрестного опыления, у рододендронов наблюдались случаи самоопыления. Первый случай самоопыления выявлен еще в бутоне, когда в результате соприкосновения пыльца из пыльников попадала на рыльце. Второй случай самоопыления наблюдался в период цветения. Так, после раскрытия цветка столбик пестика располагался выше тычиночных нитей и выходил за их пределы. Через несколько дней столбик опускался ниже тычиночных нитей, и свисающая из пыльников пыльца часто попадала на собственное рыльце. Знание о преимуществе перекрестного или самоопыления для конкретного вида дало возможность правильно использовать механизмы опыления и оплодотворения при селекционной и семеноводческой работе.
Результаты эксперимента по выявлению типов опыления показали, что при изоляции цветков самоопыление произошло только у Rh. viscosum и составило 78 %. При опылении в пределах одного растения (гейтоногамия) у Rh. japonicum завязываемость плодов составила 75 %, а у Rh. viscosum – 50 % . Опыление в пределах вида (ксеногамия) проводили только у Rh. japonicum , и оно составило 68 %. Таким образом, рододендронам присуща как автогамия, так и аллогамия. С целью выявления более перспективной формы перекрестного опыления (гейто-но- или ксеногамии) для семенной продуктивности растения Rh. japonicum проводили измерение плодов и подсчет количества семян. При гейтоногамии формировались более мелкие коробочки (2,1 см длиной и 0,7 см шириной), а при ксеногамии – более крупные (2,5 см длиной и 1 см шириной). Подсчет семян показал, что при опылении в пределах одного растения количество семян в коробочке в среднем составило 195 шт., а при опылении в пределах вида – 444 шт. Таким образом, ксеногамия является наиболее перспективной формой аллогамии, в результате которой формировалось большее количество семян.
Особенности распускания цветков
Исследование биологии цветения рододендронов в условиях юга Приморского края показало, что начало цветения Rh. japonicum и Rh. viscosum отмечалось с 26 по 29 мая при среднесуточной температуре 12 °С. Период цветения одного цветка составлял 5–7 дней, а соцветия – в среднем до двух недель. Массовое цветение обоих видов наблюдалось на 7–10-й, а окончание распускания цветков происходило в среднем на 24–28-й день (рис. 1, 2).
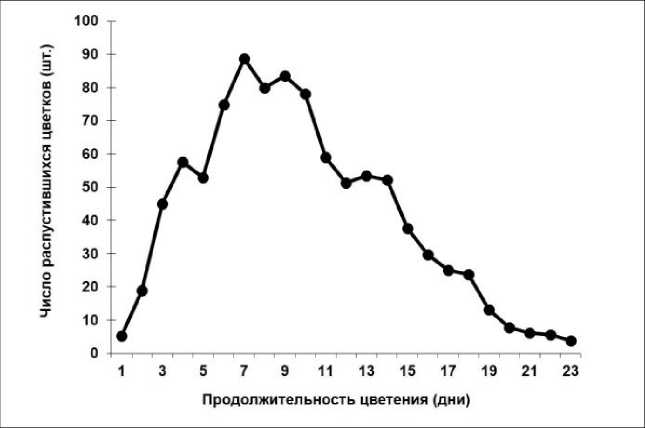
Рис. 1. Динамика цветения Rhododendron japonicum
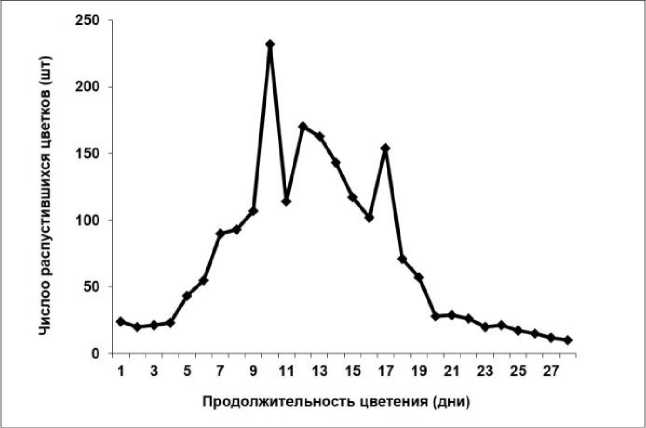
Рис. 2. Динамика цветения Rhododendron viscosum
При этом продолжительность цветения всего растения составляла в среднем месяц и зависела от размеров куста. У Rh. japonicum общее количество цветков на одном растении составляло в среднем 952 (355–1935) шт. Для Rh. viscosum характерно большее количество цветков на одно растение – 1580 (208–3354) шт.
При наблюдении за цветением на первый взгляд создавалось впечатление, что цветки непрерывно распускались в течение суток. Однако при детальном изучении суточной ритмики установлено, что цветки Rh. japonicum распускались утром в течение одного-двух часов. На рисунке 3 показан ход распускания цветков, где наибольшее число распустившихся цветков фиксировалось в 8 часов утра.
Далее, в течение дня, происходило резкое снижение распускания цветков и активное посещение их опылителями. Таким образом, для Rh. japonicum характерен утренний тип распускания цветков (см. рис. 3). Наши данные полностью противоречат результатам работы Р.В. Карамзина [6], который указывает на ночной тип распускания цветков Rh. japon-icum на Украине. Различия в сроках распускания цветков, на наш взгляд, обусловлены генетически закрепленными циркадными ритмами. Известно, что растения, лишенные экзогенных сигналов времени (температура, освещение), сохраняют суточные ритмы, указывая на их связь с эндогенными биологическими циркадными часами [13]. Таким образом, в большинстве случаев цветки открываются и закрываются, реагируя на циркадные часы [14]. В то же время циркадные часы, влияющие на время цветения, являются генетически закрепленными [15]. Поэтому мы предполагаем, что у представителя флоры Японии – Rh. japonicum распускание цветков генетически закреплено и не изменяется при интродукции растения в другой часовой пояс, а смещается на ночное время.
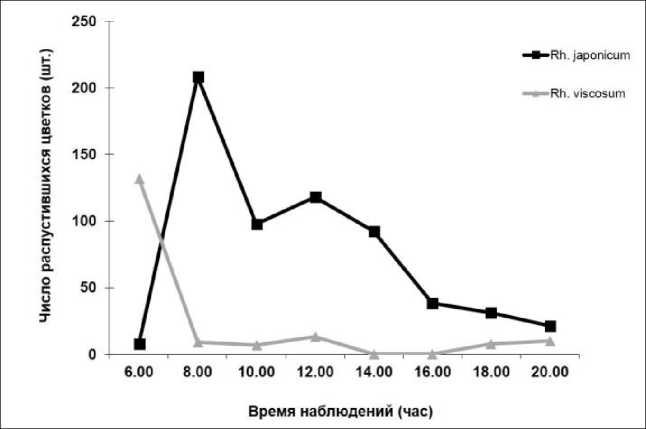
Рис. 3. Суточная динамика цветения Rhododendron japonicum и viscosum.
Противоположная ситуация наблюдается у североамериканского интродуцента – Rh. viscosum , у которого массовое распускание цветков осуществляется в ночное время (ночной тип распускания цветков) (см. рис. 3). Такой характер суточной периодичности цветения при отсутствии потенциальных опылителей в ночное время можно рассматривать как признак, сформировавшийся в иной биоценоти-ческой обстановке, либо как проявление действия циркадных часов. Поскольку данные о времени распускания цветков Rh. viscosum в условиях естественного ареала (Северная Америка) отсутствуют, можно предположить, что при интродукции этого вида в другой часовой пояс (российский Дальний Восток) также происходит смещение распускания цветков на ночное время.
Выводы. Исследование структурных особенностей цветка подтверждает принадлежность видов рода Rhododendron к энтомофилии. Наблюдения в период цветения показали, что кроме аллогамии исследуемым видам присуща контактная и гравитационная автогамия. Эксперименты по принудительному опылению подтвердили способность видов к само- и перекрестному опылению. При этом в случае аллогамии качество семян при гейтоногамии выше, чем при ксеногамии. Изучение функциональных особенностей репродуктивной сферы, а именно биологии цветения Rh. japonicum и Rh. viscosum, позволило выявить два типа распускания цветков: утренний и ночной. Различие в сроках цветения во- сточноазиатского и североамериканского видов, вероятно, обусловлено генетически закрепленными циркадными ритмами, которые сохраняются и смещают сроки распускания цветков при интродукции растений в районах с другим часовым поясом. Учитывая, что рододендроны – энтомофильные растения, ночной тип распускания цветков отрицательно сказывается на семенной продуктивности.
Список литературы Биология цветения Rhododendron japonicum и R. viscosum при интродукции на юге Приморского края
- Петухова И.П. Рододендроны на юге Приморья. -Владивосток, 2006. -131 с.
- Кокшеева И.М. Качество семян в условиях юга Приморья//Биологическое разнообразие. Интродукция растений. -СПб., 2003. -С. 391-393.
- Кокшеева И.М. К методике определения жизнеспособности пыльцы у представителей рода Rhododendron L. (Ericaceae)//Ботан. журн. -2004. -Т. 89. -№ 6. -С. 147-150.
- Кокшеева И.М. Репродуктивная биология видов рода Rhododendron L. при интродукции в Ботаническом саду-институте ДВО РАН//Вестн. КНУ. -2009. -Вып. 19-31. -С. 127-130.
- Тихменев Е.А. Цветение и опыление некоторых Вересковых (Ericaceae) на севере Дальнего Во-стока//Ботан. журн. -1979. -Т. 64. -№ 4. -С. 595-601.
- Карамзин Р.В. Биология цветения и плодоношения рододендрона японского (Rhododendron japonicum (Gray) Suring.)//Охрана, изучение и обогащение растительного мира. -1980. -Вып. 7. -С. 51-58.
- Bünning E., Zimmer R. Zur Deutung der Phasenver schiebungen und "transients" nach exogener Störung endo gener Rhythmen//Planta. -1962. -Vol. 59. -P. 1-14.
- Ewusie J.Y., Quaye E.C. Diurnal periodicity in some common flowers//New Phytol. -1977. -Vol. 78. -P. 479-485.
- Doorn W.G. van, Meeteren U. van Flower opening and closure: a review.//J. Exp. Bot. -2003. -Vol. 54. -P. 1801-1812.
- Шамурин В.Ф. Суточная ритмика и экология цветения некоторых степных растений//Ботан. журн. -1958. -Т. 43. -№ 4. -С. 548-557.
- Пономарев А.Н., Верещагина В.А. Антэкологический очерк темнохвойного леса//Проблемы биогеоценологии, геоботаники и ботанической географии. -Ленинград: Наука, 1973. -С. 56-60.
- Демьянова Е.И. Антэкология: учеб. пособие по спецкурсу. -Пермь: Изд-во Перм. гос. ун-та, 2010. -116 с.
- McClung C. Robertson. Plant Circadian Rhythms//The Plant Cell. -2006. -Vol. 18. -P. 792-803.
- Jianfang Bai, Saneyuki Kawabata Regulation of Diurnal Rhythms of Flower Opening and Closure by Light Cycles, Wavelength, and Intensity in Eustoma grandiflorum//The Horticulture Journal. -2015. -Vol. 84 -№ 2. -P. 148-155.
- Chuan Liu, Xuefeng Qu, Yanhao Zhou, Gaoyuan Song, Naghmeh Abiri, Yuhui Xiao, Fan Liang, Daiming Jiang, Zhongli Hu, Daichang Yang Os-PRR37 confers an expanded regulation of the diur-nal rhythms of the transcriptome and photoperiodic flowering pathways in rice//Plant, Cell and Enviro-ment. -2018. -Vol. 41 -№ 3. -P. 630-645.
