Биология и экология Liparis loeselii (L.) Rich. (Orchidaceae juss.) в Тверской области
 Rich. (Orchidaceae juss.) в Тверской области")
Автор: Хомутовский Максим Игоревич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 3-7 т.15, 2013 года.
Бесплатный доступ
Изучены особенности биологии и экологии Liparis loeselii (L.) Rich. на территории Тверской области. Представлены результаты наблюдений за изменением положения особей в пространстве. Описано вегетативное размножение вида. За 4 года численность исследованной ценопопуляции возросла в 2,5 раза. Местообитание L. loeselii рекомендовано к охране.
Орхидные, ценопопуляция, онтогенез, семенная продуктивность, вегетативное размножение
Короткий адрес: https://sciup.org/148202170
IDR: 148202170 | УДК: 582.594
Biology and ecology of Liparis loeselii (L.) Rich. (Orchidaceae juss.) in the Tver region
Features of biology and ecology Liparis loeselii (L.) Rich. have been studied in Tver Region. The results of observations on the movement of individuals in space are presented. The vegetative propagation of individuals was described. The population size has increased in 2.5 times for the four years of observation. The plant community with population of L. loeselii must be protected.
Текст научной статьи Биология и экология Liparis loeselii (L.) Rich. (Orchidaceae juss.) в Тверской области
L. loeselii – травянистый многолетник высотой 5 – 20 см с надземный клубнем побегового происхождения, располагающемся в толще мха, и двумя сближенными у основания листьями ланцетной формы [9]. Длина листа варьирует от 2 до 20 см. Соцветие – кисть с 1 – 10 (иногда 20) желтовато-зелеными цветками, обычно обращеными вверх (рис. 1). Листочки околоцветника около 4 мм длины, узколанцетные. Губа длиной менее 5 мм, яйцевидная, согнута посередине. Колонка длинная, с коротким наклонным пыльником на верхушке; рыльце цельное, находится на передней поверхности колонки непосредственно под пыльником. Цветение обычно проходит во второй половине июня – первой половине июля [6, 16]. Популяции L. loeselii , как правило, немногочисленны [3, 18]. Вид произрастает по низинным осоково-гипновым и осоково-сфагновым болотам с богатым минеральным питанием, обычно расположенных на выходах известняков и других карбонатных пород, а также на приозерных сплавинах [9]. В Нидерландах и Англии вид встречается на зарастающих прибрежных дюнах [19, 20]. Мелиорация земель и осушение болот, увеличение рекреационной нагрузки по берегам водоемов приводят к его исчезновению [21]. Не смотря на это, процветающие популяции L. loeselii отмечали на заброшенных торфяных карьерах в Ленинградской [22] и Липецкой [23] областях.
При культивировании растения держатся стабильно, ежегодно цветут и плодоносят. Формирующиеся семена полноценны, вегетативного размножения не наблюдали [24]. Его относят к группе видов, не способных к естественному вегетативному размножению, которые длительно сохраняются в культуре при создании специфических условий, имитирующих естественные, но их численность при этом не увеличивается [9].
По сравнению с такими видами как Dacty-lorhiza fuchsii (Druce) Soó, D. maculata (L.) Soó, Platanthera bifolia (L.) Rich. биология L. loeselii изучена недостаточно. В связи с этим целью настоящей работы становится изучение популяци- онной и репродуктивной биологии вида на территории Тверской области.
МАТЕРИАЛ И МЕТОДЫ
Ценопопуляция Liparis loeselii была изучена в 2009–2012 гг. в окр. г. Андреаполь Тверской области. На заложенных пробных площадках (по 1×10 м) проводили геоботанические описания согласно общепринятым методикам [25, 26]. Подсчет всех особей на пробных площадях проводили с учетом их онтогенетического состояния (рис. 2). Онтогенетические состояния вида выделяли на основе собственных исследований [17, 27] и согласно работам М.Б. Фардеевой [12, 28]. Для исследования морфологических особенностей особей в ЦП проводили измерения параметров у всех растений L. loeselii разных онтогенетических состояний. Измеряли следующие показатели: высоту растения, длину соцветия, число цветков (ЧЦ), число плодов (ЧП), процент образования плодов (ЧП/ЧЦ×100%), число листьев на растении, длину и ширину второго снизу листа, число жилок листа, число корней, их длина, а также диаметр псевдобульбы. Так ювенильные особи (j) L. loeselii характеризовались одним развитым листом 1,1 – 2,4 см длиной и 0,3 – 0,5 см шириной с двумя – четырьмя жилками (рис. 2). Имматурные растения (im) имели один – два листа длиной 2,0 – 6,5 см и 0,6 – 1,2 см шириной с четырьмя – восемью, реже десятью жилками. Для особей, находящихся в виргинильной стадии (v), характерно наличие двух листьев (длина 4,0 – 8,8 см и ширина 1,1 – 2,4 см) с 10 – 18 жилками. У генеративных особей (g) развиты два листа (длина, в среднем, 6,0 – 14,0 и ширина 1,7 – 3,5 см) с 18 – 28 жилками, а высота побегов колебалась в пределах от 8,0 до 28,6 см. Длина соцветия, в среднем, составляла 3,0 – 7,9 см, а количество цветков – 1 – 12 (до 17). Сенильных растений в течение наблюдаемого времени не отмечали, а так как большинство орхидей умеренной зоны, не переходя в сенильное состояние, отмирают после последнего цветения [5], то, вероятно, и в онтогенезе L. loeselii эта стадия отсутствует. Подсчет протокормов в природных условиях не проводили. За основу исследований ритмики развития орхидеи была принята методика И.Г. Серебрякова [29].
При характеристике ЦП рассчитывали общую среднюю и максимальню плотность растений на 1 кв.м; индекс восстановления ( I в ), который показывает степень семенного возобновления в цено-популяциях [30]; индекс возрастности (∆); индекс эффективности (ω). Определяли также скорость развития ( V ∆) и специфическую скорость старения ( r ∆) [30]. Для уточнения типа ЦП применяли классификацию «дельта-омега» [31]. Подсчет семязачатков в завязи цветка и семян в плоде определяли, согласно методикам В.В. Назарова [32,
33]. Были вычислены такие показатели: реальная семенная продуктивность (РСП) – число полноценных семян в плоде; потенциальная семенная продуктивность (ПСП) – число семязачатков в завязи цветка; коэффициент продуктивности (КП) – РСП / ПСП [34]. В качестве меры изменчивости признаков использовали коэффициент вариации (CV %).
Уровни варьирования признаков приняты по Г. Н. Зайцеву [35]: СV>20% – высокий, СV= 11-20% – средний, СV<10% – низкий.
При изучении семенной продуктивности собирали плоды со зрелыми семенами до начала фазы диссеменации (вскрытия плодов), измеряли их длину и ширину. Плоды хранили в бумажных пакетах в течение двух недель при температуре +24ºС, после чего закладывали в холодильник на постоянное хранение (+4ºС) [36]. Семязачатки, зрелые семена изучали с помощью светового микроскопа MICMED-5, фотоаппарата Canon Power Shot A95 и Fujifilm FinePix J37 (программы Image J), сканирующего электронного микроскопа LEO 143OVP (программа LEO SRV-32). Вычисляли среднюю длину и ширину семенной кожуры и зародыша, отношение длины к ширине, доля семян без зародыша (число семян без зароды-ша/общее число семян в плоде×100%). Объем семенной кожуры и зародыша, соотношение этих показателей и объем воздушного пространства в семени вычисляли по следующим формулам:
VS (объем семени) = 2[( w /2) 2 ×(½ l )×(1,047)], где w – ширина семенной кожуры, l – длина семенной кожуры, 1,047 – π /3;
VE (объем зародыша) = 4/3× π ×(½ l )×(½ w ) 2 , где w – ширина зародыша, l – длина зародыша;
AS (объем воздушного пространства в семени) = ( VS-VE )/ VS ×100% [37, 38]. Брали от 50 до 200 семян каждого образца. Тип семян определяли по методике R. Dressler [39]. Данные обрабатывали статистическими методами с использованием пакета Microsoft Excel.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Изученная ЦП произрастает на болоте узкой полосой 8 – 12 м шириной и около 50 м длиной (общая площадь составила около 500 кв.м). В травяном ярусе преобладают Equisetum palustre L., Carex dioica L., C. nigra (L.) Reichard, Eriophorum latifolium Hoppe, Blysmus compressus (L.) Panz. ex Link., Epipactis palustris (L.) Crantz, в моховом – Calliergon giganteum (Schimp.) Kindb. и Limprichtia cossonii (Schimp.) Anderson & al. Структура полночленной, нормального типа ЦП в первый год исследований (2009 г.) имела вид: 28j:101im:63v:36g. Максимальная плотность ЦП составила 38 особей/кв.м, а средняя – 20,3 особи/кв.м. По классификации «∆–ω»
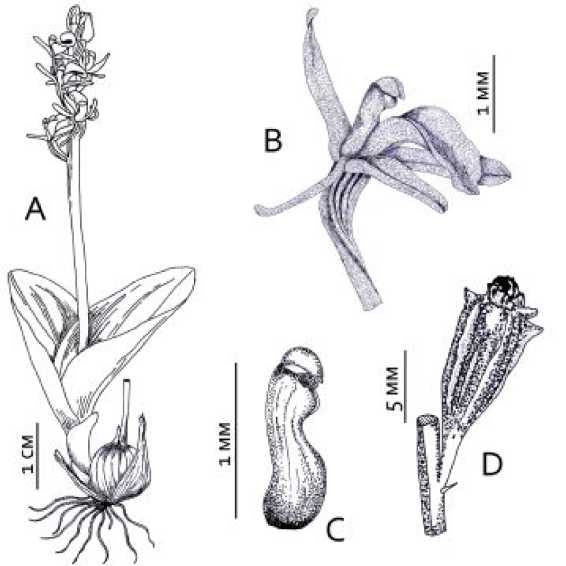
Рис. 1. Liparis loeselii .
A – внешний вид генеративного растения; B – цветок; C – колонка (вид сбоку); D – плод.
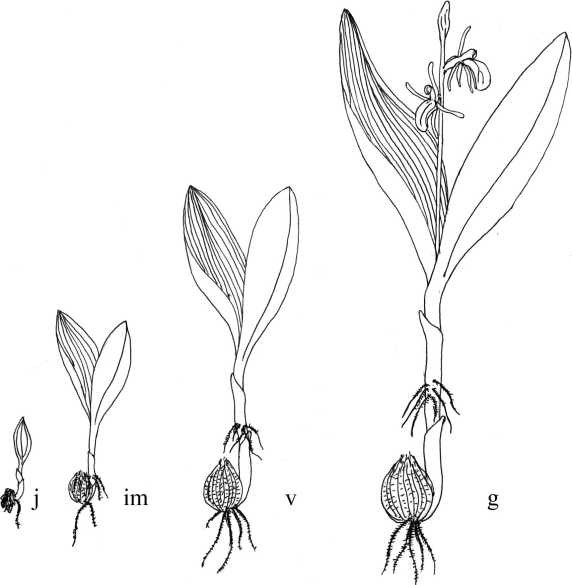
Рис. 2. Онтогенетические состояния Liparis loeselii .
Л.А. Животовского [31] ЦП является молодой (табл. 1). Возрастной спектр левосторонний, среднединамичный с преобладанием имматурных и виргинильных особей (рис. 3). Число генеративных особей в 4,5 раза меньше вегетативных. Однако, в 2012 г. их число резко возросло с 70 (в 2011 г.) до 168 и в спектре появился второй пик. В ЦП, произрастающей на территории ООПТ «Татарско-Ахметьевское болото» (Республика Татарстан) доля ювенильных особей в разные по влажности годы варьирует от 10 до 25% [28]. В ЦП, обнаруженной в 2006 г. в Киеве (Украина), ювенильные особи составили 9,3% от общей численности [40]. Изменение же численности данной группы особей в нашей ЦП в разные годы наблюдений не значительно (от 12 до 17%). Согласно литературным данным [41, 42], для видов рода Liparis характерны ЦП с бимодальным типом спектра. Несмотря на это, соотношение возрастных групп не всегда одинаково и может резко меняться от года к году. Так в ЦП, произрастающей в заповеднике Миельно (Mielno) на территории Польши, в 1995 г. преобладали генеративные особи (более 60%), тогда как уже в следующем 1996 г. численность группы снизилась и от общего числа особей ЦП составила всего 38,8% [43]. ЦП в Киеве (в 2006 г.), имела одновершинный правосторонний возрастной спектр, где преобладали также генеративные особи (51,1%) [40]. Эта ЦП произрастала на неохраняемой территории и могла подвергаться антропогенному воздействию. Данный фактор может объяснить такое соотношение возрастных групп в ЦП. Подобное наблюдается и в ЦП, произрастающей на трансформированной территории – в зарастающем карьере [44].
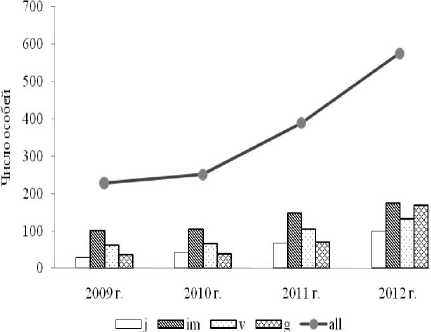
Рис. 3. Динамика возрастного спектра ЦП Liparis loeselii на минеротрофном ключевом болоте (г. Андреаполь, Тверская обл.) в 2009–2012 гг.
Однако, не только антропогенный фактор может влиять на уровень численности в ЦП. Многолетние наблюдения (1995–2010 гг.) Л. Беднорца за ЦП в заповеднике Миельно (Польша) показали, что численность особей снижается вследствие постепенного естественного зарастания болота древесными (Salix, Alnus) и травянистыми (Carex) видами растений. Численность ЦП из заповедника в первые годы наблюдений увеличивалась с 73 особей (1995 г.) до 106 (1997 г.), а после начала снижаться. В 2002 г. численность данной ЦП составила 29 особей [43], а в 2010 г. – всего 2 особи [45]. Геоботанические описания фитоценоза с участием L. loeselii, сделанные в заповеднике в 1995 г. и в 2002 г., различались. На всех трех участках, где проводили наблюдения, увеличилось количество древесных видов (Salix cinerea L., S. pentandra L., Alnus glutinosa (L.) Gaertn., Betula pubescens Ehrh.) [43, 45]. На болоте, где произрастает изучаемая нами ЦП, также присутствуют виды рода Salix (Salix aurita L., Salix rosmarinifolia L., Salix triandra L.). Увеличение проективного покрытия кустарникового яруса изменяет как освещенность местообитания, так и водный режим, а высокая чувствительность L. loeselii к изменению любых факторов, вероятно, может привести к снижению численности ЦП. Таким образом, естественное изменение количественного и видового состава ценоза также может менять соотношение возрастных групп и размер ЦП L. loeselii.
Полученная нами морфологическая характеристика особей ЦП (табл. 2) в целом совпадает с данными полученными в Республике Татарстан [28], но отличается от данных, полученных в Челябинской области и на Украине. Так генеративные особи из Ильменского заповедника (Челябинская область) по сравнению с нашими данными (табл. 2) имеют более крупные листья (длина второго листа в среднем 13,51 см, а ширина – 3,05 см) и корни, длина которых больше почти в 2 раза (в среднем 4,13 см) [46]. Генеративные особи ЦП из Киева, наоборот, небольших размеров: высота растения в среднем 11,8 см, длина листа – 6,7 см, а его ширина – 1,4 см. Но, несмотря на это, число цветков на побеге практически не отличается и составляет в среднем 6,0 [40].
Таблица 1. Динамика ЦП Liparis loeselii на минеротрофном ключевом болоте в 2009–2012 гг.
|
год |
n |
Х ср ( Х max ) |
X п |
X Г |
I в |
∆ |
ω |
V ∆ |
r ∆ |
Тип ЦП |
|
2009 |
228 |
20,3 (38) |
19,1 |
0,6 |
5,3 |
0,13 |
0,35 |
0,013 |
0,103 |
Молодая |
|
2010 |
251 |
19,2 (35) |
18,4 |
0,8 |
5,6 |
0,12 |
0,34 |
» |
||
|
2011 |
389 |
28,6 (46) |
26,2 |
2,6 |
4,6 |
0,13 |
0,34 |
» |
||
|
2012 |
575 |
39,4 (73) |
28,1 |
11,3 |
2,4 |
0,17 |
0,41 |
» |
Примечание к табл. 1 : n – численность ЦП; Х ср – общая средняя и максимальная ( Х max) плотность растений, осо-бей/1кв.м; X п – плотность прегенеративной фракции, особей/1кв.м; XГ – плотность генеративной фракции, особей/1кв.м; I в – индекс восстановления; ∆ – индекс возрастности; ω – индекс эффективности; V ∆ – скорость развития; r ∆ – специфическая скорость старения.
Таблица 2. Морфометрическая характеристика онтогенетических состояний Liparis loeselii (Тверская область, окр. г. Андреаполь)
|
Признак |
Онтогенетическое состояние |
|||
|
j |
im |
v |
g |
|
|
M±m |
M±m |
M±m |
M±m |
|
|
Длина листа, см |
1,85±0,09 |
3,87±0,11 |
6,58±0,16 |
10,45±0,53 |
|
Ширина листа, см |
0,43±0,01 |
0,80±0,01 |
1,51±0,04 |
2,60±0,12 |
|
Число листьев |
1 |
1–2 |
2 |
2 |
|
Число жилок1 |
2–4 |
4–10 |
10–18(20) |
18–28 (32) |
|
Диаметр псевдобульбы, см |
0,34±0,09 |
0,49±0,05 |
0,81±0,29 |
0,86±0,21 |
|
Число корней |
1 |
2–3 |
4–6 |
5–6 (8) |
|
Длина корней |
0,57±0,08 |
0,90±0,15 |
1,56±0,21 |
2,11±0,17 |
|
Высота растения, см |
– |
– |
– |
16,24±0,80 |
|
Длина соцветия, см |
– |
– |
– |
4,95±0,39 |
|
Число цветков |
– |
– |
– |
7,29±0,71 (17) |
Примечание к табл. 2 : 1 срединная жилка не учитывается; M – среднее арифметическое значение, m – стандартная
|
----7 4a |
5b R |
|||
|
6* -, |
||||
|
i |
||||
|
1# |
■3 |
|||
|
4 |
5 |
■ |
||
|
t 11 |
♦ 2 )•■ |
■ |
8a |
|
|
9a |
||||
|
.w |
||||
|
6 |
27 1 4 |
5 • |
♦ 12» |
|
|
n 2e |
IF |
8a |
||
|
> |
||||
2009 г. 2010 г. 2011г
|
l.l: |
e 5 1[ |
' |
||
|
-1 |
e KI |
|||
|
2e |
4b |
|||
|
9a |
||||
|
1 |
•■ |
12» |
10a |
4s |
|
Ha |
141 |
■7 |
||
|
2e |
4» |
8* |
||
|
9 |
||||
|
1^ ■ |
1 |
5* |
IP |
|
|
1U |
15» |
7 |
||
|
8« |
||||
|
* |
||||
Список литературы Биология и экология Liparis loeselii (L.) Rich. (Orchidaceae juss.) в Тверской области
- Татаренко И.В. Биоморфология орхидных (Orchidaceae Juss.) России и Японии//Автореф. дисс. … докт. биол. наук. М., 2007. 48 с.
- Пушай Е.С. Дементьева С.М. Биология, экология и распространение видов сем. Orchidaceae Juss. в Тверской области: Монография. Тверь: Твер. гос. ун-т, 2008. 206 с.
- Красная книга Российской Федерации (растения и грибы). М.: Товарищество научных изданий КМК, 2008. 855 с.
- Флора европейской части СССР. Л.: Наука, 1976. Т. 2. 236 с.
- Вахрамеева М.Г., Денисова Л.В., Никитина С.В., Самсонов С.К. Орхидные нашей страны. М.: Наука, 1991. 224 с.
- Ефимов П.Г. Род Liparis (Orchidaceae) на территории России//Ботан. журн. 2010. Т. 95, № 10. С. 1458-1480.
- Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (подписана 3 марта 1973 г. в г. Вашингтоне)//СИТЕС в России. Нижний Новгород, 1995. С. 6-52. (Охрана живой природы; Вып. 5).
- Варлыгина Т.И. Аннотированный список растений, включенных в приложения Бернской Конвенции и Директивы по охране природных местообитаний и дикой фауны и флоры//Информационно-аналитические материалы по состоянию охраны растений, животных и их местообитаний в странах Западной Европы и России (на примере Бернской Конвенции, Директивы по охране птиц и Директивы по охране природных местообитаний и дикой фауны и флоры). М., 2008. С. 91-96.
- Мамаев С.А., Князев М.С., Куликов П.В., Филиппов Е.Г. Орхидные Урала: систематика, биология, охрана. Екатеринбург: УрО РАН, 2004. 124 с.
- Куликов П.В. Конспект флоры Челябинской области (сосудистые растения). Екатеринбург -Миасс: «Геотур», 2005. 537 с.
- Баранова О.Г., Рогова Т.В., Бакин О.В. Флористические находки в республике Татарстан, Россия//Ботан. журн. 2000. Т. 85, № 4. С. 148-152.
- Фардеева М.Б. Диагнозы онтогенетических состояний Liparis loeselii (L.) Rich. (Orchidaceae)//Ботанические заметки (Казань). 2010. № 1. С. 6-10.
- Аверьянов Л. В. Орхидные (Orchidaceae) Средней России//Turczaninowia. 2000. Т. 3(1). С. 30-53.
- Благовещенский В.В., Раков Н.С. Конспект флоры высших сосудистых растений Ульяновской области. Ульяновск, 1994. 114 с.
- Гафурова М.М. Дополнения в Красную книгу Чувашской Республики (редкие и исчезающие растения) по гербарным материалам//Материалы Всерос. науч.-практ. конф. «Изучение растительных ресурсов Волжско-Камского края»: сб. науч. тр. конф. Чебоксары Чувашской Республики, 3-5 октября 2008 г./Гл. ред. д.б.н. Папченков В.Г. Чебоксары, 2008. С. 18-23.
- Ефимов П.Г. Орхидные северо-запада европейской России (Ленинградская, Псковская, Новгородская области). М.: Товарищество научных изданий КМК. 2-е изд., испр. и доп. 2012. 220 с.
- Хомутовский М.И. Характеристика ценопопуляции Liparis loeselii (L.) Rich. в окр. г. Андреаполь (Тверская область)//Современные проблемы популяционной экологии, геоботаники, систематики и флористики: материалы международной научной конференции, посвященной 110-летию А.А. Уранова (Кострома, 31 октября -3 ноября 2011 г.): в 2 т. Т. 1/отв. ред. и сост. Ю.А. Дорогова, Л.А. Жукова, И.Г. Криницын, В.П. Лебедев. Кострома: КГУ им. Н.А. Некрасова, 2011. C. 233-236.
- Варлыгина Т.И., Голубева М.А., Сорокин А.И. Состояние популяций некоторых видов орхидных Сусанинского болота в Костромской области//Охрана и культивирование орхидей. Материалы IX Международной конференции (26-30 сентября 2011г.) М.: Товарищество научных изданий КМК, 2011. С. 80-85.
- Grootjans A.P., Geelen H.W.T., Jansen A.J.M., Lammerts E.J. Restoration of coastal dune slacks in the Netherlands//Hydrobiologia. 2002. Vol. 478 (1-3). P. 181-203.
- Pillon Y., Qamaruz-Zaman F., Fay M. F., Hendoux F., Piquot Y. Genetic diversity and ecological differentiation in the endangered fen orchid (Liparis loeselii)//Conservation Genetics. 2007. Vol. 8 (1). P. 177-184.
- Красная книга Тверской области. Тверь: Вече Твери, АНТЭК, 2002. 256 с.
- Ефимов П.Г., Конечная Г.Ю. Новые находки редких видов сосудистых растений в карбонатных районах Северо-Запада европейской части России//Псковск. регионологич. журн. 2009. № 8. С. 52-62.
- Хлызова Н.Ю., Скользнева Л.Н., Недосекина Т.В. Редкие виды Cypripedium calceolus и Liparis loeselii (Orchidaceae) и на территории Липецкой области//Ботан. журн. 2009. Т. 94. № 10. С. 1594-1597.
- Коновалова Т.Ю., Шевырева Н.А. Опыт выращивания видов орхидных флоры Московской области в открытом грунте//Восстановление и мониторинг природной флоры. М.: Товарищество научных изданий КМК. 2010. С. 32-39.
- Ценопопуляции растений (Основные понятия и структура)/Под ред. А.А. Уранова, Т. И. Серебряковой. М., 1976. 216 с.
- Ценопопуляции растений (очерки популяционной биологии)/Под ред. Т.И. Серебряковой. М., 1988. 184 с.
- Хомутовский М.И. Антэкология, семенная продуктивность и оценка состояния ценопопуляций некоторых видов орхидных (Orchidaceae Juss.) Валдайской возвышенности: Дис.. канд. биол. наук. М., 2012. 237 с.
- Фардеева М.Б. Онтогенез липариса Лезеля (Liparis loeselii (L.) Rich.)//Онтогенетический атлас растений: научное издание. Том VI/Мар. гос. ун-т; отв. ред. проф. Л.А. Жукова. Йошкар-Ола, 2011. С. 188-192.
- Серебряков И.Г. О ритме сезонного развития растений подмосковных лесов//Вестн. МГУ. 1947. № 6. С. 75-108.
- Жукова Л.А. Популяционная жизнь луговых растений. Йошкар-Ола: РИИК «Ланар», 1995. 224 с.
- Животовский Л.А. Онтогенетическое состояние, эффективная плотность и классификация популяций//Экология. 2001. № 1. С. 3-7.
- Назаров В.В. Определение реальной семенной продуктивности у Dactylorhiza romana и D. incarnata (Orchidaceae Juss.)//Ботан. журн. 1988. Т. 73. № 2. С. 231-233.
- Назаров В.В. Методика подсчета мелких семян и семяпочек (на примере сем. Orchidaceae)//Ботан. журн. 1989. Т. 74. № 8. С. 1194-1196.
- Левина Р.Е. Репродуктивная биология семенных растений (Обзор проблемы). М.: Наука, 1981. 96 с.
- Зайцев Г.Н. Методика биометрических расчетов//Математическая статистика в экспериментальной ботанике. М., 1973. 256 с.
- Буров А.В., Широков А.И., Коломейцева Г.Л. О создании банка семян редких видов орхидных умеренной зоны//Биологический вестник, Т. 8, № 1, 2004, С. 8-11.
- Arditti J., Michaud J.D., Haeley P.L. Morphometry of orchid seeds. I. Paphiopedium and native California and related species of Cypripedium//Amer. J. Bot. 1979. Vol. 66, № 10. P. 1128-1137.
- Healey P.L., Michaud J.D., Arditti J. Morphometry of orchids seeds. III. Native of California and related species of Goodyera, Piperia, Platanthera, and Spiranthes//Amer. J. Bot. 1980. Vol. 67. P. 508-518.
- Dressier R.L. Phylogeny and classification of the orchid family. Portland, Oregon: Dioscorides press, 1993. 314 p.
- Шевченко М.С., Тимченко I.А., Парнiкоза I.Ю. Унiкальне мiсцезнаходження Liparis loeselii (L.) Rich. в м. Києвi//Укр. ботан. журн. 2007. Т. 64. № 3. С. 438-443.
- Татаренко И.В. Орхидные России: жизненные формы, биология, вопросы охраны. М.: Аргус, 1996. 207 c.
- Фардеева М.Б. К изучению популяции Liparis loeselii (L.) Rich. на территории РТ//Вопросы общей ботаники: традиции и перспективы. Ч.2./Материалы Международной научной конференции, посвященной 200-летию Казанской ботанической школы. Казань, 2006. С.125-127.
- Bednorz L. Population dynamics of Liparis loeselii (L.) L.C. Rich. in the nature reserve «Mielno» -some results from a 8 year study//Electronic Journal of Polish Agricult. Univ. Biology. 2003. Vol. 6 (2). -http://www.ejpau.media.pl/series/volume6/issue2/biology/art-06.html.
- McMaster, R.T. The population biology of Liparis loeselii, Loesel's twayblade, in a Massachusetts wetland//Northeastern Naturalist. 2001. Vol. 8(2). P. 163-178.
- Bednorz L. Regression of Liparis loeselii population in the nature reserve ‘Mielno' (Wielkopolska)//Rocz. AR Pozn. 390, Bot. Stec. 2011. 15. P. 31-33.
- Лесина С.А., Куянцева Н.Б. Онтогенетические состояния и биоэкологические особенности Liparis loeselii (L.) Rich. (Ильменский гос. заповедник, Южный Урал)//Биоразнообразие и биоресурсы Урала и сопредельных территорий: материалы V Всерос. науч.-практ. конф. Оренбург, 2010. С. 87-90.
- Смирнова Е.С. Морфология побеговых систем орхид ных. М.: Наука, 1990. 208 с.
- Залукаева Г.Л. Особенности онтогенеза тропических и субтропических орхидей в оранжерейной культуре: Дис.. канд. биол. наук. М., 1990. 245 с.
- Назаров В.В. О возникновении дочерних групп у ятрышника пурпурного//Природоохранные исследования экосистем горного Крыма: Сб. науч. ст. Симфирополь: СГУ, 1986. С. 23-25.
- Catling P.M. Rain-assisted autogamy in Liparis loeselii (L.) L.C. Rich. (Orchidaceae)//Bulletin of the Torrey Botanical Club, 1980. Vol. 107. No. 4. P. 525-529.
- Хомутовский М.И. Эффективность опыления некоторых видов орхидных Валдайской возвышенности//Охрана культивирование орхидей. Материалы IX Международной конференции (26-30 сентября 2011 г.). М.: Товарищество научных изданий КМК, 2011. С. 456-461.
- Назаров В.В. Репродуктивная биология орхидных Крыма: Дис. … канд. биол. наук. СПб., 1995. 294 с.
- Коломейцева Г.Л., Антипина В.А., Широков А.И., Хомутовский М.И., Бабоша А.В., Рябченко А.С. Семена орхидей: развитие, структура, прорастание. М.: ГЕОС, 2012. 352 с.
- Хомутовский М.И. Вегетативное размножение Liparis loeselii (L.) Rich. (Orchidaceae Juss.)//Симбиоз-Россия 2012: материалы V Всероссийского с международным участием медико-биологического конгресса молодых ученых, Тверь, 3-8 декабря 2012 г./Тверской государственный университет, Тверская государственная медицинская академия. Тверь: Издательство «Заповедник Времени», 2012. C. 444-445.
