Биология и становление жизненной формы копытеня зибольда -Asarum sieboldii Miq.(сем. Aristolochiaceae Juss.)
")
Автор: Безделева Тамара Аркадьевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 3, 2016 года.
Бесплатный доступ
Резюме. Asarum sieboldii Miq. - ценное лекарственное растение, произрастающее на юге Приморского края. Жизненная форма копытеня Зибольда - коротко-длиннокорневищный симподиально нарастающий травянистый поликарпик с розеточным побегом. У вида формируется 2 типа корневищ: укороченные эпигеогенные и смешанные гипогеогенно-эпигеогенные. Для вида характерна розеточная симподиальная модель побегообразования. У A. sieboldii, кроме проростков с двумя семядолями, развиваются проростки с одной семядолей. В онтоморфогенезе вида выделено 9 онтогенетических состояний и 6 фаз морфогенеза.
Жизненная форма, побег, корневище, лист, онтогенетическое состояние, фаза морфогенеза
Короткий адрес: https://sciup.org/146116725
IDR: 146116725 | УДК: 581.41(571.63)
Biology and development of life forms of Asarum sieboldii Miq. (Aristolichiaceae Juss.)
Asarum sieboldii Miq. is valuable medicine plant, which grows in the South of Primorie (Russian Far East). The life form of A. sieboldii is short- or longrhizomed policarpous herbaceous plant with sympodial growth and rosette sprout. Species forms two types of rhizomes: shortened epigeogenous and mixed hypogeogenous and epigeogeonous. Species is characterized by the rosette sympodial model of the sprouting. Besides having two-cotyledontous sprouts, A. sieboldii has single-cotyledontous ones. Morphogenesis of the plant consists of 9 ontogenetic stages and 6 phases.
Текст научной статьи Биология и становление жизненной формы копытеня зибольда -Asarum sieboldii Miq.(сем. Aristolochiaceae Juss.)
Введение. Род Asarum L. сем. Aristolochiaceae Juss. включает около 70-100 видов, широко распространенных в умеренной зоне северного полушария: в Азии, Европе и Северной Америке, с центром разнообразия в Восточной Азии, где сосредоточено наибольшее число видов (около 65 видов произрастают в Японии, Китае, Корее и на Тайване) (Харкевич, 1987; Huang et al., 2003). Большинство видов Asarum. мелкие, низкие, многолетние корневищные травы, растущие в тенистых, влажных широколиственных лесах.
Копытень Зибольда - один из видов рода Asarum L., ареал которого в пределах Российского Дальнего Востока ограничивается югом Приморского края; за пределами России встречается в Японии, Китае и Корее. Вид обитает во влажных, тенистых хвойных и хвойношироколиственных лесах на богатой гумусом почве, а также хорошо себя чувствует во вторичных широколиственных лесах.
С древности многие виды азиатских копытней используются в традиционной медицине Китая, Японии, Кореи и других стран в качестве лекарственных средств. Биологически активные вещества, накапливаемые различными видами копытня, обладают широким спектром фармакологической активности (Копытько и др., 2013). Копытень Зибольда также считается ценным лекарственным растением, сырье из которого используется в Тибетской медицине (Воробьев и др.,
1966) и особенно ценится в Китае, где его считают диким или горным женьшенем. В корнях Asarum sieboldii Miq. содержится - 1,4%, а в корневищах - 2,85-3,2% эфирного масла. Корневища и корни копытня содержат алкалоид азарин, гликозиды, в том числе сердечной группы и флавоноиды, смолистые и дубильные вещества, органические кислоты и их соли, крахмал, слизь стероидные сапонины (Горяев, 1952). При жевании корня появляется сильное слюноотделение и чувство онемения во рту (Куренцова, 1941; личные наблюдения). В экспериментальных условиях обнаружено стимулирующее влияние настоя к. Зибольда на центральную нервную систему (Ивон, 1971). Корневища и корни А. sieboldii в традиционной медицине используются в терапии различных инфекционных заболеваний (Копытько и др., 2013). По данным многих авторов (Фруентов, 1974; Шретер, 1975; Журавлев и др., 2004) отвар копытеня широко используется в традиционной медицине. Применение корневищ копытня в традиционной восточной медицине при лечении бронхиальной астмы подтверждено экспериментально. В русской народной медицине иногда используют как рвотное и для отрезвления. При этом необходимо помнить о ядовитости растения. Кроме того, А. sieboldii очень декоративен и может быть рекомендован для озеленения городских территорий.
Для понимания закономерностей сложения общего габитуса травянистых растений очень важен анализ в онтогенезе структуры их побеговой системы, которая формируется на основе наследственных признаков, определяющих потенциальные возможности побегообразования. Биология, онтоморфогенез, ритм сезонного развития, поведение этого вида в различных фитоценозах остаются неизученными.
Цель работы: изучение особенностей жизненной формы, онтоморфогенеза и биологии A. sieboldii в связи с перспективностью использования его в качестве лекарственного и культурного растения.
Материал и методы. Материалом для исследования послужили сборы в широколиственном лесу (окр. Академгородка, г. Владивосток), в кедрово-широколиственных лесах в окрестностях г. Владивостока и в природном заповеднике «Уссурийский» (Уссурийский р-н). Сбор материала и наблюдение за видом велись боле 10 лет.
Онтоморфогенез и жизненная форма A. sieboldii изучались по методикам, разработанным Т.А. Работновым (1950), И.Г. Серебряковым (1962, 1964), А.А. Урановым (1967, 1975) и их учениками.
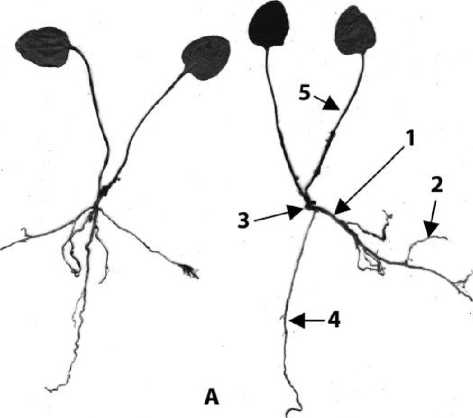
Результаты и обсуждение: Копытень Зибольда (A. sieboldii) представляет собой многолетнее травянистое поликарпическое растение с погруженным в лесную подстилку корневищем с резким специфическим запахом. Корневая система образована многочисленными придаточными корнями, равномерно отходящими по всей длине поверхности корневища. Длина придаточных корней в среднем 10-15 см, диаметр 0,2 см.
Побег розеточный, несущий в основании 2-3 чешуевидных и выше 2 ассимилирующих листа. Ассимилирующие листья длинночерешковые, почковидные, цельнокрайные, с заостренной верхушкой, снизу опушенные короткими светлыми волосками. Побег заканчивается терминальным цветком. Почка возобновления крупная, развивается в пазухе верхнего ассимилирующего листа, ветвление происходит за счет более мелких почек в пазухах низовых чешуевидных листьев. Биологической особенностью жизненной формы Л. sieboldii является формирование двух типов корневищ: укороченных эпигеогенных и смешанных, состоящих из удлиненного гипогеогенного и укороченного эпигеогенного корневища. Из годичных приростов укороченных розеточных побегов формируется эпигеогенное корневище. Из спящих почек расположенных на эпигеогенных корневищах развиваются гипогеогенные корневища, растущие вначале плагиотропно, а затем переходящие к ортотропному росту и формирующие эпигеогенное корневище.
Цветет в апреле - мае. Цветок одиночный, верхушечный, трехмерный, имеет зачаточный, недоразвитый венчик и 3 коричневобурых, мясистых треугольно-яйцевидных чашелистика. Чашелистики срастаются в бутоне боковыми сторонами. Тычинок 12, расположеных в 2 круга. Завязь нижняя. Плод - мясистая синкарпная коробочка.
A.sieboldii - вид близкородственный европейско-сибирскому A. europaea L. Однако эти виды отличаются по ритму сезонного развития: A. europaea - зимнезеленый вид - перезимовывает с зелеными листьями (Смирнова, Зворыкин, 1974), в то время как A. sieboldii - летнезеленый вид, у которого период вегетации длится с начала апреля и до конца сентября-середины октября. К концу вегетации отмирают все листья.
В онтогенезе A. sieboldii нами выделены 4 возрастных периода и 9 онтогенетических состояний: латентный период - покоящиеся семена; предгенеративный период - проросток, ювенильное, имматурное и виргинильное состояния; генеративный период - молодое, зрелое и старое генеративные состояния; постгенеративный период -субсенильное и сенильное состояния.
Латентный период. Семена многочисленные продолговатообратнояйцевидные, имеют форму полумесяца, почти гладкие, бурые, с вдающимся швом и расположенным в нем мясистым придатком, превышающим по длине семя (рис. 1). Длина семени 3-3,5 мм, ширина 1,5 мм. Вес 1000 семян 4 г 250 мг. Семена этого вида с недоразвитым зародышем. По данным С.В. Нестеровой (1990) доразвиваются семена к. Зибольда в течение 35-40 дней и уже осенью трогаются в рост -зародышевый корешок выходит за пределы семенной кожуры, растет вплоть до заморозков, углубляется на 2,5-3 см в почву и в таком состоянии зимует. По наблюдениям Г.П. Рысиной (1973) семена Asarum europaeum L. также имеют недоразвитый зароды и прорастают в сентябре - октябре: корешок выходит за пределы семенной кожуры и достигает больших размеров. Семядоли остаются внутри семени; в таком состоянии проросшие семена перезимовывают.

Рис. 1. Семена Asarum sieboldii
Предгенеративный период. Проросток. Проростки A. sieboldii появляются в мае - июне. Прорастание надземное: семядоли выносятся на дневную поверхность на черешках длиной до 3 и более см. Листовая пластинка семядоли яйцевидная с тупой верхушкой, 1 см длины и 0,6-0,7 см ширины. Корневая система в начале вегетационного периода часто представлена только главным корнем, достигающим длины 3-4(7) см. Гипокотиль до 3 мм длины и почти 1 мм толщины, хорошо отличается от корня по цвету и диаметру. Снизу вверх диаметр гипокотиля увеличивается. К концу вегетационного периода на главном корне некоторых проростков образуется 1-2 боковых корня, и от гипокотиля отходит придаточный корень. На верхушке побега, между основаниями черешков семядолей, располагается верхушечная почка, которая дает начало побегу будущего года (рис. 2А). Фаза проростка длится до конца вегетационного периода, который заканчивается в сентябре месяце. У А. еигораеа, по данным Рысиной (1973), семядоли отмирают только к осени второго года жизни.
Биологической особенностью A. sieboldii является наличие у него кроме 2-семядольных также 1-семядольных проростков, у которых одна из семядолей не развивается (рис. 2Б).

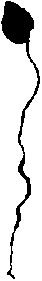
Б
Рис. 2 . Проростки Asarum sieboldii А - с двумя семядолями; Б - с одной семядолей 1 — главный корень, 2 — боковой корень, 3 - гипокотиль, 4 - придаточный корень, 5 — семядоля
Ювенильное онтогенетическое состояние длится несколько лет, начиная со второго года развития. Из верхушечной почки проростка весной следующего года развивается годичный побег ювенильного растения.
Двулетние ювенильные особи имеют корни длиной от 3,5 до 4,8 (5) см. Гипокотиль достигает 0,5-0,7 см длины. Годичный побег розеточный, несущий (2) 3 чешуевидных листа и 1 ассимилирующий лист. Длина листового черешка до 3,7 см. Лист почковидный с сердцевидным основанием и небольшой выемкой на верхушке, 1-1,7 см
-81 - длиной и 1,3-1,7 см шириной. В основании черешка ассимилирующего листа находится небольшая выемка, в которой скрыта верхушечная почка побега (рис. 3). В ювенильном состоянии растение находится до 5(6) лет. Пятилетняя ювенильная особь незначительно отличается по структуре от двулетней особи. Все еще сохраняется главный корень. Отличие 5-6летних особей от 2-летних в том, что у них от основания гипокотиля всегда отходят придаточные корни, достигающие больших размеров в сравнении с главным, а также наблюдается ветвление главного корня. Число боковых корней колеблется от 3-4 до 10, длина их 0,5-3,5 см. Длина придаточных корней от 2,3 до 8,7 см. Гипокотиль (0,3) 0,5-0,7(1) см длины; на нем намечается коленчатый изгиб в месте отхождения придаточного корня. Иногда боковые корни отходят от главного с той же стороны, с которой отходит придаточный корень от гипокотиля, что способствует втягиванию верхушечной почки в глубь почвы — корни работают как втягивающие (контрактильные). Побег в течение первых лет развития нарастает моноподиально, за счет верхушечной почки, формирующей розеточный побег. Годичный прирост 1-2 мм длины и несет (2) 3 чешуевидных листа и 1 ассимилирующий лист. К концу ювенильного возрастного состояния увеличиваются размеры листа: длина листового черешка - до 5,8 см, длина листовой пластинки - до 8 мм, ширина - до 10 мм. По краю листовой пластинки располагаются прозрачные волоски, направленные к верхушке листа. Характерной особенностью ювенильных растений является наличие хорошо заметной выемки на верхушке листа. С возрастом увеличивается длина придаточных корней, в то время как длина главного корня увеличивается незначительно. Его раст затем и вовсе прекращается.
Имматурное онтогенетическое состояние. В имматурном состоянии приостанавливается нарастание главного корня в длину и питание растения; его прикрепление к субстрату выполняют придаточные корни.
Побег с неполным циклом развития в течение нескольких лет нарастает моноподиально, формируя ежегодно розеточный побег, который несет на себе 2-3 чешуевидных листа и 1 ассимилирующий лист (рис. 4А). Через несколько лет верхушечная меристема прекращает свою деятельность и моноподиальное нарастание сменяется на симподиальное. Возобновление происходит за счет боковой почки, развивающейся в пазухе верхнего чешуевидного листа. Однако особенностью изучаемого вида является то, что побеги следующих порядков несколько лет нарастают моноподиально, причем наблюдается смена моноподиального и симподиального нарастания (рис. 4Б).
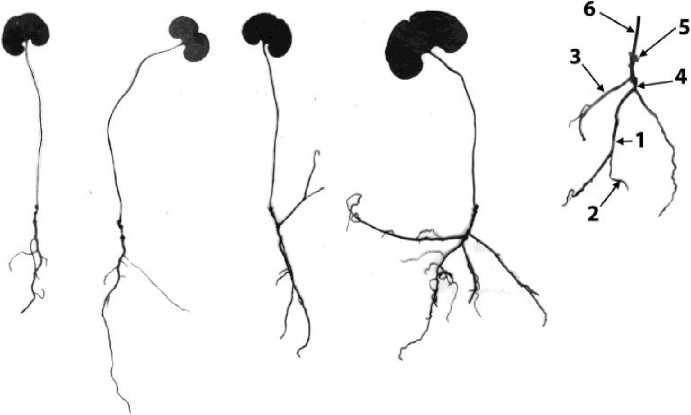
Рис. 3 . Ювенильные особи Asarum sieboldii 1—главный корень, 2 — боковой корень, 3—придаточный корень, 4 — гипокотиль, 5 —чешуевидные листья, 6 —черешок ассимилирующего листа
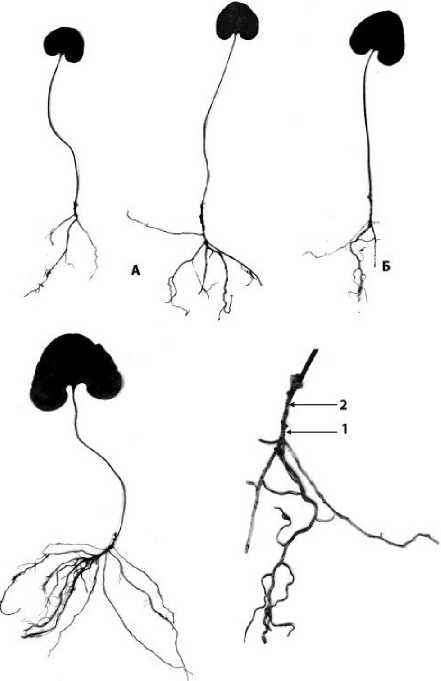
Рис. 4 . Имматурные особи Asarum sieboldii К — особи с моноподиально нарастающим простым побегом, Б — особь с симподиально ветвящимся побегом 1 —полициклический побег первого порядка;
2 —полициклический побег второго порядка
Основной особенностью данного онтогенетического состояния является изменение формы листовой пластинки, которая теряет выемку на верхушке. Лист в имматурном состоянии длинночерешковый, почковидный с тупой округлой верхушкой. В этом возрастном состоянии черешок и листовая пластинка становятся опушенными. На верхней стороне листовая пластинка опушена очень короткими волосками по жилкам; с нижней стороны лист опушен по всей поверхности короткими светлыми волосками. В пазухе чешуевидных листьев закладываются почки, остающиеся некоторое время спящими. Длительность имматурного онтогенетического состояния составляет несколько лет; к его концу из годичных приростов розеточных побегов формируется укороченное эпигеогенное корневище - резид; происходит увеличение размеров листа и корневой системы.
Особи в ювенильном и иногда в имматурном возрастных состояниях могут быть отнесены к одноосной стержнекорневой фазе морфогенеза. Часть особей имматурного онтогенетического состояния относятся к одноосной короткокорневищной фазе морфогенеза. В данных возрастных состояниях особи Л. sieboldii моноцентрические.
Виргинилъное онтогенетическое состояние. Побег остается розеточным и несет на себе 2-3 чешуевидных листа и 1 ассимилирующий лист. Нижний чешуевидный лист - 0,4 см, верхний -0,7 см длины. Ассимилирующий лист длинночерешковый, почковидный с приостренной верхушкой (рис. 5). И черешок, и листовая пластинка опушенные. Черешок достигает 13 см длины, расширенное основание его окружает верхушечную почку - почку возобновления. Листовая пластинка до 6 см длины и до 6,2 см ширины, глубина выемки в основании листовой пластинки достигает 2,1 см, ширина 2,6 см на выходе и 1 см возле черешка. В пазухе чешуевидных листьев закладываются почки, которые некоторое время остаются спящими.
Монокарпический побег многолетний, розеточный с неполным циклом развития: в течение нескольких лет нарастает моноподиально, затем верхушечная почка заканчивает цикл развития и возобновление происходит за счет почки, расположенной в пазухе верхнего чешуевидного листа. Годичные приросты 0,2-0,5 см длины формируют укороченное наклонное корневище, по которому можно определить относительный возраст особи. Абсолютный возраст корневища и особи определить сложно, т.к. корневище, нарастая на верхушке, отмирает в основании. От годичных приростов отходят придаточные корни до 10 и более см длины, ветвящиеся в своей нижней половине. Главный корень в виргинильном возрастном состоянии полностью отмирает.
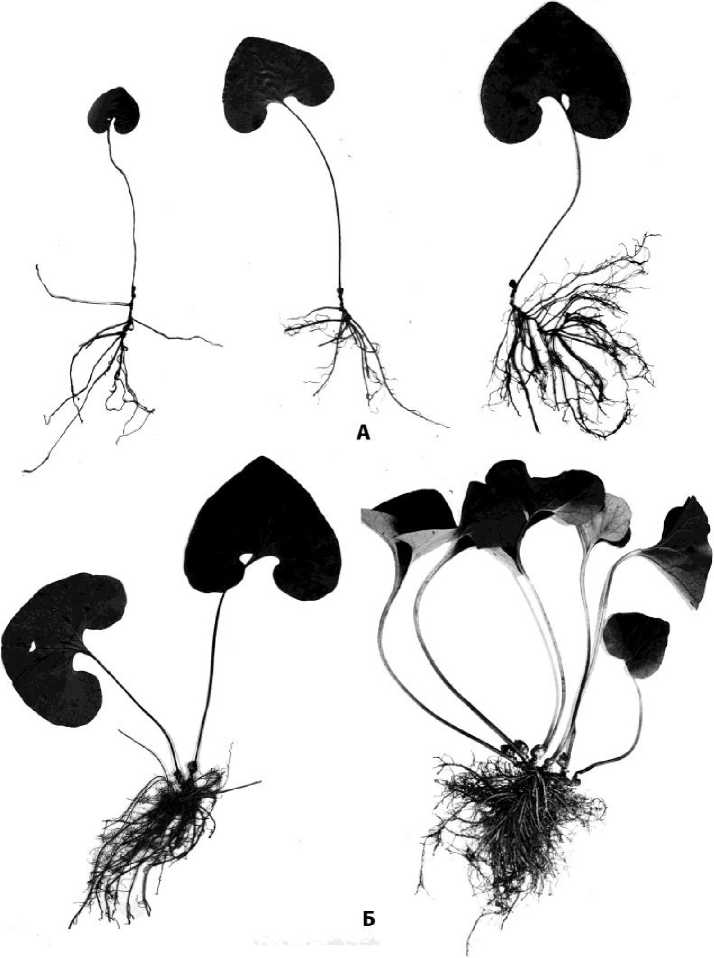
Рис. 5 . Виргинильные особи Asarum sieboldii К - простые (неветвящиеся) моноцентрические особи Б - ветвящиеся неявно полицентрические особи
Особенностью данного онтогенетического состояния является начало ветвления особи. Из спящих почек, расположенных на корневищах развиваются боковые побеги, которые также нарастают моноподиально и формируют корневища второго порядка (рис. 5Б).
Часть особей этого возрастного состояния относятся к одноосной короткокорневищной фазе морфогенеза (рис. 5А), другая часть особей относится к многоосной короткокорневищной фазе морфогеннеза (рис. 5Б).
Генеративный период. Молодое генеративное онтогенетическое состояние. Особь, достигая генеративного остояния, формирует генеративные побеги, несущие 2-3 чешуевидных и 2 ассимилирующих листа и заканчивающиеся одиночным цветком (рис. 6А). По окончании генерации цикл развития побега завершается, и моноподиальное нарастание сменяется на симподиальное. Возобновление происходит за счет боковой почки, формирующейся в пазухе одного из ассимилирующих листьев генеративного побега. Таким образом, для A. sieboldii характерна симподиальная розеточная модель побегообразования. Годичные приросты образуют укороченное эпигеогенное корневище. С возрастом наряду с эпигеогенными начинают формироваться и гипогеогенные корневища в результате плагиотропного подземного нарастания побегов. Спящие почки, раскрываясь, дают начало побегам, которые некоторое время нарастают плагиотропно и формируют удлиненные гипогеогенные корневища, а затем, переходя к ортотропному росту, образуют надземный розеточный побег, который в течение нескольких лет нарастает моноподиально, давая начало розеточным годичным приростам и эпигеогенному корневищу. В результате формируется сложное корневище, состоящее из удлиненной гипогеогенной части и укороченной эпигеогенной. Удлиненные корневища ветвятся, образуют на своих концах парциальные кусты; особь захватывает новые территории, в результате чего формируется куртина (рис. 7), состоящая из нескольких парциальных кустов. У куртины наблюдается несколько центров побегообразования, особь становится полицентрической, наблюдается ее вегетативное разрастание.
Взрослое генеративное онтогенетическое состояние (G2). Во взрослом генеративном состоянии особь достигает максимального развития. Продолжается ветвление корневищ и формирование более разветвленной куртины. На особи развивается до 5-10 генеративных побегов и несколько вегетативных (рис. 6Б). Генеративные побеги несут по 3 чешуевидных и по 2 ассимилирующих листа. Заканчивается побег одним крупным цветком. Черешок листа до 17-18см длины; л. пластинка 9,6-12,8 см длины и 11,4-15,3 см ширины. Корневая система разрастается формируя большое количество придаточных корней до 20 и более см длины и 1-1,5 мм в диаметре.
Увеличиваются размеры и разветвленность особи. В данном онтогенетическом состоянии формируется куртина до 20-25 см в диаметре, несущая несколько парциальных кустов на концах корневищ. Парциальные кусты могут быть однопобеговые и многопобеговые. Побеги находятся на разной ступени развития: на концах одних корневищ развиваются вегетативные побеги, на концах других побеги достигают генеративного состояния.
По определению А.Б. Безделева и Т.А. Безделевой (2006) жизненная форма A. sieboldii - летнезеленый травянистый тонко-длиннокороткокорневищный симподиально нарастающий поликарпик с розеточным побегом. К этому необходимо добавить, что монокарпический побег этого вида полициклический и в течение нескольких лет нарастает моноподиально.
Особи молодого и зрелого генеративного возрастных состояний относятся к фазе морфогенеза - куртина.



Рис. 6 . Генеративные особи Asarum sieboldii
А — молодая генеративнаяособь, Б — средневозрастно генеративные особи, В — старая генеративная особь
Старое генеративное состояние характеризуется деградацией корневой системы (уменьшением числа и размеров придаточных корней), сокращается число генеративных побегов, формирующихся на особи, и цикл развития монокарпических побегов до побегов с неполным циклом развития (рис. 6В). В результате нарастания корневища на верхушке и его отмирания на другом конце наблюдается распад особи на несколько самостоятельных. Формируется клон, состоящий из разновозрастных парциальных кустов рамет, заканчивающих цикл развития. В данном состоянии наблюдается незначительное вегетативное размножение.
Постгенеративный период. Субсенильное онтогенетическое состояние. В этом возрастном состоянии растение теряет способность формировать генеративные побеги, инаблюдается дальнейшее сокращение мощности корневой системы и уменьшение длины и диаметра корневищ. Особи в субсенильном возрастном состоянии отличаются крупными листовыми пластинками и образованием побегов с неполным циклом развития.

Рис. 7 . Куртина Asarum sieboldii
В сенильном возрастном состоянии уменьшается длина корневищ и остается только несколько придаточных корней. Особенностью субсенильных и сенильных растений является развитие крупных ассимилирующих листьев: до 10,2-11см дл. и до13,8-14,б см шир.; уменьшение длины черешка до 7,0-9,7 см. Кроме того, верхушка листа теряет приостренность и становится тупой, округлой.
Выводы. Проведенное изучение онтоморфогенеза жизненной формы и биологических особенностей Asarum sieboldii показало, что:
-
1. Жизненная форма A. sieboldii - длинно-короткокорневищный полицентрический симподиально нарастающий травянистый поликарпик с розеточным полициклическим монокарпическим побегом и мощной корневой системой. Вид является вегетативно подвижным травянистым
-
2. Для вида характерны короткие эпигеогенные, длинные гипогеогенные и смешанные гипогеогенно-эпигеогенные корневища.
-
3. Монокарпический побег розеточный полициклический. Наиболее длительный период развития монокарпического побега наблюдается в предгенеративном периоде; далее цикл развития монокарпических побегов постепенно сокращается вплоть до побегов с неполным циклом развития у растений в субсенильном и сенильном онтогенетических состояниях
-
4. Для вида характерна розеточная симподиальная модель побегообразования.
-
5. Биологической особенностью A. sieboldii формирование наряду с 2-семядольными 1-семядольных проростков.
-
6. В онтогенезе A. sieboldii веделено 4 периода и 9 онтогенетических состояний
-
7. В морфогенезе A. sieboldii нами выделены латентный период и следующие фазы развития: проросток, одноосная стержнекорневая, одноосная короткокорневищная, многоосная короткокорневищная, куртины и клона.
многолетником, максимально приспособленным к жизни в хвойношироколиственных и широколиственных лесах.
Выражаю глубокую благодарность Волонец Е.Б. - ст. н. с. БПИ ДВ О РАН и Калинкиной В.А. — ст. н. с. БСИ ДВО РАН за предоставленные фотографии
Безделева Т.А. Биология и становление жизненной формы копытеня Зибольда - Asarum Siebolidii Miq. (сем. Aristolochiaceae Juss.) / Т.А. Безделева // Вести. ТвГУ. Сер.: Биология и экология. 2016.№ 3. С. 77-91.
Список литературы Биология и становление жизненной формы копытеня зибольда -Asarum sieboldii Miq.(сем. Aristolochiaceae Juss.)
- Безделев А.Б., Безделева Т.А. 2006. Жизненные формы семенных растений российского Дальнего Востока. Владивосток: Наука. 296 с.
- Воробьев Д.П., Ворошилов, В.Н., Горовой П.Г., Шретер, А.И. 1966. Определитель растений Приморья и Приамурья. М.-Л.: Наука. 490 с.
- Горяев М.И. 1952. Эфирные масла флоры СССР. Алма-Ата: Изд-во АН КазССР, 380 с.
- Ивон В.Г. 1971. Некоторые показатели состояния ЦНС при сравнительном изучении препаратов из копытня Зибольда и к. европейского//Биологически активные вещества флоры и фауны Дальнего Востока и Тихого океана. Владивосток: Примор. кн. изд-во. С. 53.
- Журавлев Ю.Н., Воронкова Н.М., Баркалов В.Ю., Воронков А.А. 2004. Лекарственные растения Курильских островов. Владивосток: Дальнаука. 306 с.
- Копытько Я.Ф., Щиревич Н.Н., Сокольская., Т.А., Маркарян А.А., Даргаева Т.Д. 2013. Применение, химический состав и стандартнзация растительного сырья лекарственных средств из растений рода Asarum L.//Химко-фармацевтический журн. Т. 47 № 3. С. 33-44.
- Куренцова Г.Э. 1941. Лекарственные растения советского Дальнего Востока//Тр. Дальневост. Горнотаеж. станции им. акад. В.Л. Комарова. Ворошилов-Уссурийский: АН СССР. Т. 4. С. 131-226.
- Нестерова С.В. 1990. Прорастание и изменение жизнеспособности семян некоторых травянистых растений//Тезисы докладов Всесоюзного научного совешания (к 30-летию Ставропольского ботанического сада) «Охрана, обогащение, воспроизводство и использование растительных ресурсов» Ставрополь. C. 178-180.
- Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах//Тр. Ботан. ин-та им. В.Л. Комарова АН СССР. Сер. 3. Геоботаника. Вып. 6. С. 7-204.
- Рысина Г.П. 1973. Ранние этапы онтогенеза лесных травянистых растений Подмосковья. М.: Наука. 215 с.
- Серебряков И.Г. 1962. Экологическая морфология растений: жизненные формы покрытосеменных и хвойных. М.: Высшая школа. 378 с.
- Серебряков И.Г. 1964. Жизненные формы высших растений и их изучение//Полевая геоботаника. М.-Л.: Наука. Т. 3. С. 146-205.
- Смирнова О.В., Зворыкин К.В. 1974. Копытень европейский//Биологическая флора Московской области. М.: Изд-во Моск. ун-та. Вып. 1. С.41-51.
- Уранов А.А. 1967. Онтогенез и возрастной состав популяций//Онтогенез и возрастной состав популяций цветковых растений. М.: Наука. С.3-9.
- Уранов А.А. 1975. Возрастной состав ценопопуляции как функции времени и энергетических волновых процессов//Науч. докл. высш. школы. Биол. науки. № 2. С.7-34.
- Фруентов Н.К. 1974. Лекарственные растения Дальнего Востока. Хабаровск: Кн. изд-во. 400 с.
- Харкевич С.С. 1987. Сем Кирказоновые -Aristolochiaceae Juss.//Сосудистые растения советского Дальнего Востока. Владивосток: Наука. Т. 2. С.19-21.
- Шретер А.И. 1975. Лекарственная флора советсконо Дальнего Востока. М.: Медицина, 1975. 328 с.
- Huang S.M., Kelly L.M., Gulbert M.C. 2003. Asarum//Flora of China/Z.Y. Wu and P.H. Raven (eds.), Vol. 5. Science Press, Reijing and Missouri Botanical Garden Press, St. Louis. P. 246-257.
