Биология размножения, морфометрия и популяционная динамика пяти видов славок рода Sylvia на Куршской косе (Восточная Прибалтика)
")
Автор: Паевский Владимир Александрович
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 602 т.19, 2010 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140151922
IDR: 140151922
Breeding biology, morphometrics and population dynamics of Sylvia warblers on the Courish spit, Eastern Baltic
Текст статьи Биология размножения, морфометрия и популяционная динамика пяти видов славок рода Sylvia на Куршской косе (Восточная Прибалтика)
Зоологический институт РАН,
Университетская набережная, д. 1, Санкт-Петербург, 199034, Россия
Поступила в редакцию 29 сентября 2010*
Среди видов птиц рода Sylvia наиболее широко распространёнными и многочисленными можно считать 5 видов: ястребиную славку S. nisoria , славку-завирушку S. curruca , серую славку S. communis , садовую славку S. borin и славку-черноголовку S. atricapilla . Сведения по распространению, экологии и биологии размножения этих видов обобщены в крупных сводках по птицам Палеарктики (Cramp et al . 1992; Glutz von Blotzheim, Bauer 1991), в региональных сводках (Ко-релов 1972; Зацепина 1978; Мальчевский, Пукинский 1983; Левин, Губин 1985; и др.), а также в монографиях (Siefke 1962; Schmidt 1981; Berthold et al . 1990).
На Куршской косе Балтийского моря (Калининградская область) все пять видов славок были многочисленными в 1960-е – 1980-е годы как в гнездовое время, так и во время миграций. Сведения об их численности, срокам миграций и гнездования, а также по разным аспектам биологии размножения частично уже опубликованы сотрудниками Биологической станции «Рыбачий» Зоологического института РАН, их студентами и аспирантами (Кушнирук 1962; Белопольский, Одинцова 1969; Ефремов, Паевский 1973; Виноградова 1983, 1986, 1988; Паев-ский 1971, 1985, 1991, 1992, 2008; Соколов 1991а,б, Sokolov et al . 1998; Payevsky 1999, 2009; Матанцева, Симонов 2007, 2008). За период наших сорокалетних исследований численность славок претерпевала существенные изменения. Это побудило нас обратить особое внимание на детали популяционной динамики славок и их размножение.
В Европе особый интерес к популяционной динамике птиц был вызван резким снижением численности ряда видов (особенно насекомоядных и хищных), что в первую очередь было связано с использованием в сельском и лесном хозяйстве пестицидов и гербицидов. Возможность использовать количество пойманных с целями кольцевания птиц как меру их численности демонстрировалась одинаковыми трендами этого показателя в разных регионах, а в ряде случаев – сходством трендов местных и пролётных птиц (Hjort, Lindholm 1978; Svensson 1978; Busse, Cofta 1986; Baumanis 1990; Соколов 1991б, 1999; Соколов и др. 2001; Berthold, Fiedler 2005; и др.). Мнения большинства орнитологов сходились в том, что необходимыми условиями использования данных отлова для учёта численности должны быть следующие: а) один и тот же стандартный метод отлова на одном и том же месте, б) одни и те же сроки отлова из года в год, в) анализ только долговременных трендов по многолетним данным. Существует также возможность сравнивать данные отлова с другими формами учётов птиц. В частности, не использован в этом плане и такой простой показатель, как ежегодное количество найденных гнёзд при стационарном и долговременном изучении какой-либо популяции.
Цель данной статьи – изложить данные, полученные при изучении пяти видов славок сотрудниками Биологической станции ЗИН РАН «Рыбачий» в течение 40 лет. Для этого были проанализированы материалы по миграциям, размножению, возрастной структуре и популяционной динамике этих видов. Природные условия Куршской косы, протянувшейся с юго-запада на северо-восток и отделяющей Куршский залив от Балтийского моря, уже были подробно описаны (Паевский 1971, 2008, 2009 и др.). Исследования проведены в разных местах Куршской косы, но наибольшее число сведений по регулярно контролируемым гнёздам собрано в район полевого стационара «Фрин-гилла», расположенного в 12 км южнее посёлка Рыбачий. На этой территории неоднократно проводились прореживание леса и санитарные рубки, во время которых по недоразумению были частично вырублены также и привлекательные для славок шиповник, барбарис, можжевельник и кустистая горная сосна.
Отлов птиц с целью кольцевания и прижизненного обследования проводился с помощью стационарных рыбачинских ловушек, действующих с апреля до ноября (их описание см.: Дольник, Паевский 1976). Многие из окольцованных птиц неоднократно ловились в последующие годы (например, один самец славки-завирушки попадался в ловушки 18 раз на протяжении 8 лет). За 1959-1994 годы на полевом стационаре всего было поймано 25120 особей пяти видов славок, из них славки-завирушки 8932 (в среднем за год 248), садовой 6299 (175), черноголовой 3826 (106), серой 3754 (104) и ястребиной 2309 особей (в среднем 109 за год в 1959-1974 и 28 – в 1975-1994 годах). Количество данных, используемых для анализа разных аспектов размножения и популяционной динамики, указано в соответствующих таблицах и рисунках. У пойманных в ловушки птиц регистрировались два измерения: длина крыла (максимальная длина вытянутого и уплощенного крыла) с точностью до миллиметра, и масса тела с точностью до десятой доли грамма. Яйца, птенцы и гнёзда измерялись и взвешивались с точностью до сотой доли. Пол и возраст пойманных птиц с 1972 года определяли по методике Svensson (1970), которая была впоследствии проверена и дополнена нами (Виноградова и др. 1976). Расчёты успешности размножения проведены по методу первоначального определения ежедневной скорости гибели содержимого гнёзд (Mayfield 1975). Поскольку пролёт славок по косе продолжался и в тот период, когда многие местные особи уже приступили к гнездованию, проводилось разделение пойманных птиц на местных и пролётных. Для этого были установлены конкретные сроки для каждого вида, когда пойманные птицы давали в последующие годы наибольшую долю повторных отловов (Соколов 1991б).
Сроки мигр аций и размножения
Результаты определения средних дат основных явлений летней жизни славок приведены в таблице 1. Видовые различия в сроках весеннего прилёта и осеннего отлёта достаточно чёткие. Попарное сравнение сроков прилёта по средним многолетним данным показало, что их различия во всех случаях статистически значимы (величина t от 3.5 до 11.2, P < 0.01 и P < 0.001), за исключением пары черноголовка – славка-завирушка, где значимых различий между видами нет.
Таблица 1. Сроки миграций и размножения славок на Куршской косе по данным за 1959-1994 годы
|
Показатели |
S. atricapilla |
S. borin |
S. communis |
S. curruca |
S. nisoria |
|
Пределы Медиана |
14.4 - 10.5 30.4 |
Первая пои 5.5 - 19.5 12.5 |
мка весной 25.4 - 14.5 4.5 |
14.4 - 11.5 1.5 |
8.5 - 26.5 18.5 |
|
13.5 - 29.5 |
Сроки весенней миграции 20.5 - 4.6 18.5 – 31.5 |
11.5 – 31.5 |
28.5 – 9 6 |
||
|
Число гнёзд Самое раннее по годам Пределы Медиана |
Срок появления первого яйца 257 289 38 421 15.5 – 9.6 23.5 – 9.6 — 10.5 – 8.6 15.5 – 10.7 23.5 – 13.7 25.5 – 29.6 10.5 – 13.7 4.6 8.6 12.6 30.5 |
325 22.5 – 7.6 22.5 – 27.6 5.6 |
|||
|
Пределы Медиана |
Пер 7.7 – 16.8 23.7 |
вая молодая 6.7 – 17.8 24.7 |
птица в ловушке 6.7 -19.8 25.6 -24.7 24.7 8.7 |
6.7 – 26.7 15.7 |
|
|
Пределы Медиана |
22.9 - 17.11 24.10 |
Последняя птица осенью 13.9 – 28.10 22.8 – 3.10 12.10 17.9 |
4.9 – 22.10 26.9 |
29.7 – 26.9 25.8 |
|
Средние сроки осеннего отлёта разных видов также достоверно различаются во всех случаях при P < 0.001. Наибольшая ежегодная длительность летнего пребывания оказалась у черноголовки (прилетает раньше других видов славок, улетает позже других). Наименьшая длительность – у ястребиной славки (прилетает позже других видов, улетает раньше других). Хотя весеннее появление славки-завирушки столь же раннее, как и у черноголовки, её осеннее исчезновение происходит в среднем на месяц раньше, чем у черноголовки. Садовая и серая славки занимают среднее положение по обсуждаемым датам, хотя отдельные особи садовой славки задерживались осенью до последних чисел октября (что, несомненно, связано с ярко выраженной способностью этого вида переходить осенью на питание ягодами, что также характерно и для черноголовки, и неоднократно наблюдалось в Прибалтике). В целом длительность пребывания на территориях размножения и послегнездовых кочёвок, рассчитанная по датам поимок первых и последних птиц, составляет у черноголовки в среднем 178 дней, у садовой славки 153 дня, у славки-завирушки 149 дней, у серой славки 136 дней и только 100 дней у ястребиной славки.
Пути осенней миграции как местных славок, так и пролётных через Куршскую косу, пролегают у садовой славки в юго-западном направлении, через Италию, а у черноголовки и ястребиной славки – в юговосточном направлении, через восточное Средиземноморье (Паевский 1971; Bolshakov, Shapoval, Zelenova 2001).
Анализ ежегодных вариаций сроков весенней миграции птиц на Куршской косе (Sokolov et al . 1998) показал, что чем выше температура воздуха в мае, тем раньше прилетают черноголовка и славка-завирушка, но у серой и садовой славок такой зависимости не найдено.
Сроки откладки первого яйца у разных видов славок, в отличие от сроков прилёта, в среднем почти не различаются (табл. 1). При одновременном сравнении сроков откладки у всех пяти видов по критерию Фридмана не было получено статистически значимых различий ( χ 42 = 8.27, n.s.), хотя у славки-завирушки сроки оказались достоверно более ранними, чем у серой славки при P < 0.05. Видовая специфика интервала между датой прилёта и датой появления первого яйца в среднем весьма определённа: 19 дней у ястребиной, 27 у садовой, 29 у славки-завирушки, 37 у черноголовки и 39 дней – у серой славки.
Ежегодные вариации сроков откладки яиц и вылупления у славок вполне определённы и достоверны. Как было показано нами ранее (Sokolov, Payevsky 1998), более высокие температуры воздуха весной коррелируют с более ранним вылуплением (значимая отрицательная связь обнаружена для славки-завирушки с температурой апреля, для садовой и ястребиной – с температурой мая, и только у серой славки – с температурой июня). За все годы исследований самое раннее появ- ление первого яйца отмечено у черноголовки и садовой славки в 1990 году, а самое позднее у всех видов славок – в 1987 году.
Видовая специфика средних сроков отлова первых молодых птиц почти не выражена (табл. 1). Достоверно раньше других видов появляются лишь молодые птицы у славки-завирушки ( P < 0.05), что соотносится с более ранней откладкой яиц. Годовые же вариации в сроках отлова первых молодых оказались высоко значимыми. Ранее было показано (Sokolov, Payevsky 1998), что чем выше температура воздуха весной, в апреле и мае, тем раньше регистрируются первые молодые птицы у всех видов славок.
Таблица 2. Длина крыла и масса тела славок, пойманных в апреле, мае и июне в разные периоды лет
|
Годы |
Длина крыла, мм |
Масса тела, г |
||||
|
n |
Пределы |
Среднее |
n |
Пределы |
Среднее |
|
|
S. |
atricapilla 33 |
|||||
|
1967-1974 |
185 |
72-81 |
76.4 ± 0.1 |
178 |
13.8-22.8 |
17.6 ± 0.1 |
|
1990-1994 |
207 |
71-81 |
76.1 ± 0.1 |
207 |
14.5-22.2 |
17.9 ± 0.1 |
|
S. |
atricapilla . . |
|||||
|
1967-1974 |
163 |
71-81 |
76.0 ± 0.2 |
157 |
14.6-22.7 |
17.8 ± 0.1 |
|
1990-1994 |
186 |
72-81 |
76.2 ± 0.1 |
186 |
15.6-24.2 |
18.9 ± 0.1 |
|
S. borin |
||||||
|
1967-1974 |
517 |
74-85 |
79.5 ± 0.1 |
448 |
14.9-23.5 |
18.8 ± 0.1 |
|
1990-1994 |
344 |
73-85 |
79.8 ± 0.1 |
344 |
15.6-24.9 |
19.1 ± 0.1 |
|
S. |
communis 33 |
|||||
|
1967-1974 |
146 |
69-78 |
73.4 ± 0.2 |
107 |
12.4-18.0 |
14.3 ± 0.1 |
|
1990-1994 |
165 |
68-81 |
73.6 ± 0.1 |
165 |
11.5-17.7 |
14.9 ± 0.1 |
|
S. |
communis . . |
|||||
|
1967-1974 |
120 |
67-76 |
71.6 ± 0.2 |
96 |
12.5-19.1 |
14.6 ± 0.1 |
|
1990-1994 |
133 |
68-77 |
72.3 ± 0.1 |
133 |
13.0-19.0 |
15.1 ± 0.1 |
|
S. curruca |
||||||
|
1967-1974 |
286 |
58-72 |
65.1 ± 0.2 |
162 |
10.1-14.2 |
11.7 ± 0.1 |
|
1990-1994 |
408 |
61-72 |
65.4 ± 0.1 |
408 |
8.6-15.4 |
11.6 ± 0.1 |
|
S. nisoria 33 |
||||||
|
1967-1974 |
217 |
83-93 |
88.0 ± 0.1 |
204 |
20.1-28.7 |
23.8 ± 0.1 |
|
1987-1994 |
28 |
84-91 |
87.8 ± 0.4 |
28 |
20.6-26.8 |
23.9 ± 0.4 |
|
S. nisoria . . |
||||||
|
1967-1974 |
143 |
80-91 |
87.4 ± 0.2 |
136 |
21.1-28.9 |
24.1 ± 0.1 |
|
1987-1994 |
33 |
82-90 |
85.9 ± 0.3 |
33 |
20.5-27.3 |
24.2 ± 0.3 |
Размеры и масса половозрелых птиц
Поскольку все пойманные птицы измерялись и взвешивались, было проведено сравнение длины крыла и массы тела в два периода, разделённых промежутком в 15 лет: 1967-1974 (среднее за 8 лет) и 1990-1994 (среднее за 5 лет); лишь для ястребиной славки вследствие падения её численности второй период был увеличен до 8 лет (19871994). Проверялось предположение, что рост и снижение численности популяций может сопровождаться изменением физических параметров, в частности размеров. Результаты сравнения приведены в таблице 2. В большинстве случаев и размеры, и масса тела отличались лишь на несколько десятых долей, и на первый взгляд были одинаковы. Только в двух случаях изменения были явными: средняя масса тела самок черноголовки достоверно увеличилась на 1.1 г (t = 6.25, P < 0.001), а средняя длина крыла самок ястребиной славки достоверно уменьшилась на 1.5 мм (t = 4.11, P < 0.001). Хотя статистически значимыми при P < 0.05 и P < 0.01 оказались и другие изменения (увеличение размера крыла у садовой славки и самок серой славки и увеличение массы тела у садовой и серой славок), объяснить это какими-либо явлениями жизни славок не представляется возможным.
Предпочитаемые биотопы и плотность гнездования
В большинстве случаев гнездовые территории разных видов славок на Куршской косе находились в одних и тех же биотопах. Наиболее плотно были заселены славками участки смешанного леса с подлеском из смородины и ежевики, а также опушки высокоствольных участков с кустарниковым бордюром. В таких биотопах встречалось наибольшее количество гнёзд садовой, черноголовой и ястребиной славок. Только славка-завирушка могла заселять внутренние части молодого соснового леса, и только серая славка иногда гнездилась в очень низких ивовых кустиках на морских дюнах. Большинство гнездовых территорий ястребиной славки находилось в открытых биотопах: небольшие прогалины и низкорослый кустарник на опушках сосновых и смешанных участков леса. В таких же местах гнездился в 1960-х и 1970-х годах и сорокопут-жулан Lanius collurio , и их гнездовые территории часто перекрывались. Численность этих обоих видов резко снизилась в 1980-х годах и они практически одновременно исчезли с исследуемой территории.
Плотность гнездования наиболее детально была исследована у ястребиной славки (Виноградова 1986, 1988). В 1974-1978 годах она составляла 16-26, в среднем 20 пар на 1 км 2 , а затем, в 1982-1986 годах, в связи с резким падением численности этого вида на Куршской косе, снизилась до 4.0, а позже – до 0.4 пар на 1 км 2 . В 1980 году у шести гнездовых пар ястребиных славок размер территорий составлял от 700 до 1250 м 2 , в среднем 1050 м 2 . В 1986 году было найдено последнее гнездо ястребиной славки. В последующие годы лишь в 1997-м были снова обнаружены два гнезда этого вида.
Совместная гнездовая плотность черноголовой и садовой славок в наиболее благоприятных для них местах составляла 0.94 пар/га, а на остальной площади не превышала 40 пар/км 2 для каждого из этих видов. Гнездовая плотность славки-завирушки в наиболее предпочитаемых ею местах достигала 2 пар/га, а на остальной площади в среднем составляла около 60 пар/км 2 .
Расположение и размеры гнёзд
Расположение гнезд было исследовано по 657 гнёздам S. curruca , 566 гнездам S. nisoria , 384 гнёздам S. atricapilla , 344 гнёздам S. borin и 63 гнёздам S. communis . Гнезда славок располагались на 28 видах растений и предпочитаемыми были: для черноголовки – красная смородина ( χ 12 = 18.3, P < 0.001), для садовой – она же и ежевика ( χ 12 = 27.2, P < 0.001), для серой – ежевика ( χ 12 = 27.5, P < 0.001), для славки-завирушки – можжевельник, обыкновенная и горная сосна ( χ 12 = 57.2, P < 0.001), для ястребиной – ежевика, красная смородина, горная и обыкновенная сосна ( χ 12 = 21.7, P < 0.001). Однако частота распределения гнёзд на разных видах растений в разные годы варьировала в очень широких пределах (по критерию Фридмана χ 62 = 4.7, n.s.). Доля гнёзд на красной смородине достоверно увеличилась в 1981-1995 годах ( χ 12 = 9.8, P < 0.01), а на обыкновенной сосне резко уменьшилась ( χ 12 = 16.3, P < 0.001). Рост числа гнёзд на можжевельнике в 1981-1995 годах проявился в основном у славки-завирушки. Все эти изменения, несомненно, связаны с лесохозяйственными работами – прореживанием леса и обрубанием нижних ветвей у обыкновенной и горной сосен.
Относительно высоты расположения гнёзд выяснено, что средняя высота зависела как от вида славок, так и от года её регистрации (двухфакторный дисперсионный анализ, F = 10.29, P < 0.001). Наиболее высоко располагала гнёзда славка-завирушка (0.2-5.5, в среднем 1.29 м), наиболее низко – серая славка (0.05-0.6, в среднем 0.22 м). Те же показатели у других видов таковы: черноголовка – 0.1-2.9, в среднем 0.80 м, садовая – 0.1-2.5, в среднем 0.51 м, ястребиная – 0.1-2.5, в среднем 0.7 м. Достоверное изменение высоты расположения гнёзд выявлено у 2 видов. У ястребиной славки эта высота постепенно увеличивалась с 1959 по 1985 год ( r = 0.97, P < 0.01), а у славки-завирушки с 1959 по 1980 год увеличивалась ( r = 0.96, P < 0.001), а с 1980 по 1995 год уменьшалась ( r = -0.98, P < 0.01).
Некоторые данные были собраны и по размеру гнёзд у черноголовки и садовой славки. Размеры, см ( H – высота гнезда, h – глубина лотка, D – диаметр гнезда, d – диаметр лотка) были следующими. По 14 гнёздам черноголовки: H 6.5-9.3, в среднем 7.48, h 3.3-5.5, в среднем 3.90, D 9.3-10.6, в среднем 9.84, d 5.5-6.3, в среднем 5.91. По 12 гнёздам садовой славки: H 6.0-9.2, в среднем 8.29, h 5.0-6.2, в среднем 5.48, D
9.5-12.4, в среднем 11.24, d 6.0-7.0, в среднем 6.30. Вес пустых гнёзд, высушенных при температуре 60°С, составил у черноголовки от 3.94 до 7.21, в среднем 5.03 г ( n = 14), у садовой славки – от 6.07 до 9.93, в среднем 7.66 г ( n = 8).
Размеры и масса яиц
Измерения (в мм) 58 яиц у черноголовой, 43 у садовой и 26 у славки-завирушки дало следующие результаты. У S. atricapilla длина 17.0-20.4, в среднем 18.75±0.11, ширина 13.3-15.8, в среднем 14.46± 0.06; у S. borin длина 18.1-21.0, в среднем 19.75±0.68, ширина 13.415.4, в среднем 14.61±0.09; у S. curruca длина 16.1-18.1, в среднем 16.9 ±0.12, ширина 11.0-14.1, в среднем 12.73±0.1. Масса (в г) яиц в день их снесения составила у черноголовой ( n = 63) 1.82-2.99, в среднем 2.23± 0.03, у садовой ( n = 52) 2.08-2.73, в среднем 2.37±0.02, у славки-завирушки ( n = 31) 1.30-1.57, в среднем 1.43±0.01. Взвешивание яиц в день их снесения (5 кладок черноголовки и 6 кладок садовой) обнаружило тенденцию увеличения массы каждого последующего яйца, однако статистически значимым это оказалось только для 3 кладок садовой славки (ранговый коэффициент корреляции Спирмэна r s от 0.90 до 1.00, P < 0.05, P < 0.01) и одной кладки черноголовки ( r s = 1.00, P = 0.0). Тем не менее масса первого яйца и у черноголовки, и у садовой славки всегда была меньше веса последнего из отложенных яиц. Масса птенцов сразу же после вылупления составлял у черноголовки от 1.58 до 2.25, в среднем 1.94±0.05 г ( n = 18), а у славки-завирушки – от 1.05 до 1.46, в среднем 1.24±0.04 г ( n = 12).
Поведение насиживания и наседные пятна
Поведение насиживания и наседные пятна славок были предметом специальных исследований, результаты которых опубликованы (Ефремов, Паевский 1973), поэтому здесь будут кратко приведены только их основные положения. У всех пяти видов славок ночью на гнезде оставались только самки, и продолжительность их пребывания на гнезде в тёмное время суток составляет от 5 до 8.6 ч, не обнаруживая достоверных видовых отличий. Продолжительность однократного насиживания в светлое время суток очень вариабельна у всех видов и у обоих полов. У славки-завирушки и у серой славки продолжительность общего пребывания самца на гнезде увеличивалась к концу периода насиживания. Самцы всех видов участвовали не только в насиживании кладки, но и в обогревании птенцов. И самки, и самцы славки-завирушки и ястребиной славки при очередной смене партнёра приносили коконы пауков, которыми постепенно обрамляли гнездо. Участие самцов в насиживании, выраженное в виде средней доли суток, приходящейся на насиживающего самца, оказалось одинаковым у ястребиной, серой и славки-завирушки (26.6%, 27.6%, 27.5%) и статистически значимо (P < 0.001) более высоким у них, чем у черноголовой и садовой славок (6.3%, 5.0%).
Наседные пятна у самцов обсуждаемых пяти видов оказались выраженными у всех исследованных гнездовых особей, за исключением серой славки. Это подтвердилось и при осмотре всех самцов серой славки, пойманных в ловушки. Ни у одного самца S. communis из пойманных в период гнездования не было обнаружено выпадения перьев. Среди самцов славок наиболее чётко наседные пятна были выражены у славки-завирушки и ястребиной славки. Наседные пятна самцов претерпевали все стадии развития, как и у самок – выпадение перьев, гиперваскуляризацию и утолщение кожи, кожный отёк, стадию восстановления.
К постоянному насиживанию все исследованные виды приступали обычно после откладки четвертого, реже пятого яйца. Продолжительность регулярного насиживания до начала вылупления птенцов составляла у черноголовки и садовой славки 11-12 сут, у славки-завирушки также 11-12, но иногда 13 сут, у ястребиной славки – 12-13, однажды 14 сут. Последовательность вылупления не всегда точно соответствовала очерёдности откладки яиц.
Величина кладки и успешность размножения
У всех пяти исследованных видов славок максимальная величина кладки составляла 6 яиц. Таких кладок было 85 (19.6%) у славки-завирушки, 19 (5.4%) у ястребиной, 15 (5.2%) у черноголовой, 5 (11.9%) у серой и 3 (1.0%) у садовой славки. Средняя величина кладки по пятидневкам в течение сезона размножения достоверно уменьшалась у славки-завирушки от 5.50 до 4.46, составляя в общем среднем за все годы 5.13; у черноголовой – от 5.03 до 3.91, в среднем 4.73; у садовой – от 5.04 до 4.57, в среднем 4.76; у ястребиной – от 4.91 до3.91, в среднем 4.90; у серой в общем среднем 4.95.
Частичные потери при размножении вызывались эмбриональной смертностью и некоторой долей неоплодотворённых яиц. В среднем на каждую полную кладку приходилось неразвившихся яиц у серой славки 0.09, у ястребиной 0.22, у славки-завирушки 0.23, у черноголовой 0.30 и у садовой 0.33. От всех отложенных яиц, сохранившихся до вылупления, неразвившиеся яйца составили от 1.8% у серой до 7.0% у садовой славки. Основные потери яиц и птенцов вызывались хищничеством. Из хищников, разоряющих гнёзда славок, на Куршской косе отмечены куница, горностай, ласка, белка и бродячие кошки.
Усреднённые данные по величине кладок славок, успешности вылупления и выкармливания птенцов приведены в таблице 3. Изменения величины кладки и успешности размножения в разные годы были
Таблица 3. Результативность размножения славок за 1959-1995годы
Успешность размножения у славки-черноголовки была статистически значимо выше в 1980-е по сравнению с 1970-ми и 1990-ми годами, а у садовой славки выше в 1960-е, чем в 1970-е и 1980-е. И лишь у ястребиной славки успешность размножения в 1970-е годы была выше, чем в 1960-е и одинакова с успешностью размножения в 1980-е годы.
Отмеченные изменения средней величины кладки и успешности размножения в популяциях славок Куршской косы, скорее всего, связано с многолетними колебаниями весенних температур воздуха. Ранее было установлено (Sokolov, Payevsky 1998), что повышение средних температур воздуха апреля и мая в 1960-е и 1980-е годы по сравнению с 1970-ми и 1990-ми годами привело к более раннему гнездованию птиц, в том числе и славок. Можно полагать, что и бóльшая величина кладок, и более высокая успешность размножения также связаны с более комфортными условиями гнездования.
Возрастная структур а популяций, смертность и продолжительность жизни
По данным повторных отловов меченых особей в последующие после кольцевания годы была рассчитана возрастная структура для трёх видов славок. В весенне-летний период, до появления птенцов, годовалые птицы (рождения прошлого года) составляли 53.5±2.0% у ястребиной славки, 60.1±3.5% у садовой славки и 58.1±2.2% у славки-завирушки. Двухлетние птицы составляли, соответственно, 22.0±1.7, 21.2 ±2.9, 22.8±1.8, а трёхлетние – 11.7±1.3, 10.1±2.1, 10.4±1.3%. Доли птиц более старшего возраста для каждой возрастной категории составляли 6% и менее. Особей старше 5 лет на каждую тысячу птиц приходится 32 у ястребиной, 16 у садовой и 24 у славки-завирушки.
Максимальная зарегистрированная продолжительность жизни у ястребиной славки была 9 лет, у славки-завирушки 8 лет, у садовой славки 7 лет, причём этого максимума достигали только самцы. Это находится в полном соответствии с результатами изучения нами дифференцированной по полу выживаемости. В отличие от других животных, у птиц более высокая выживаемость оказывается присуща самцам (Payevsky et al . 1997; Паевский 2008). Средняя ожидаемая продолжительность жизни после достижения половозрелости составляла 1 год и 6 месяцев у ястребиной и 1 год и 3 месяца у садовой и славки-завирушки. Средняя ежегодная выживаемость у славки-завирушки составила 47.7% (Паевский 1992), а у ястребиной славки – 49.7% (Payevsky et al . 1997), но у последней значения ежегодной выживаемости год от года непрерывно снижались (Payevsky et al . 2003).
Половая структура популяции была определена только у славки-черноголовки. Доля самцов весной и летом составила 56.6±1.6%, осенью – 55.2±1.7% (Паевский 1993).
Популяционная динамика
Тенденции изменения численности славок на Куршской косе, наряду с другими видами птиц, неоднократно приводились в разных публикациях биостанции «Рыбачий». Для 1960-1976 годов было показано (Дольник, Паевский 1979), что у садовой и серой славок численность снижалась, у славки-завирушки увеличивалась, у черноголовки испытывала небольшие колебания, а у ястребиной до 1969 года увеличивалась, а затем снижалась. Отлов в последующие годы дал возможность проследить изменения численности до 1986 года (Паевский 1990). Снижение численности продолжалось у ястребиной славки, а у серой, садовой и черноголовой славок появилась тенденция к популяционному росту, хотя статистически значимой она была лишь для птиц, пойманных в летне-осенний период. Рассмотрение численности сугубо местных птиц (пойманных в июне и июле) показало, что между температурой воздуха в апреле и мае и численностью молодых птиц есть положительная связь у черноголовой, серой и садовой славок, тогда как у ястребиной и славки-завирушки такой связи не обнаружено (Sokolov 1999). Тенденции изменения численности славок и по данным отлова, и по количеству найденных гнёзд за 1974-1990 годы были определены по коэффициенту корреляции временных рядов (коэффициент односторонней регрессии) и по коэффициенту ранговой коре-ляции Кендэла, где один ряд переменных представлен числом лет, а другой – количеством птиц или гнёзд. Результаты представлены в общем виде на рисунках 1-5, а в подробностях расчётов – в таблице 4. Оба типа коэффициентов корреляционной зависимости в большинстве случаев дали сходный результат.
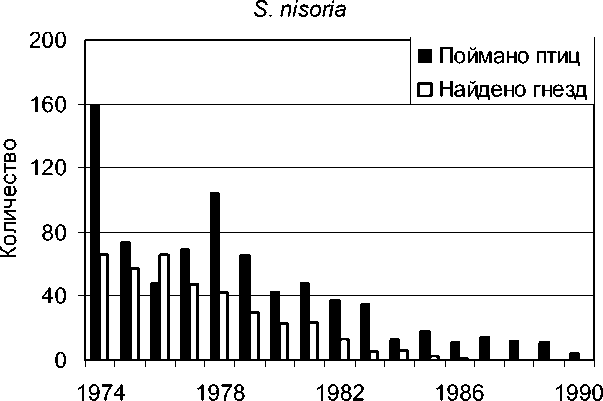
Рис. 1. Количество отловленных особей и найденных гнёзд у ястребиной славки.
S. curruca
-
■ Поймано птиц
-
□ Найдено гнезд
1974 1978 1982 1986 1990
Рис. 2. Количество отловленных особей и найденных гнёзд у славки-завирушки.
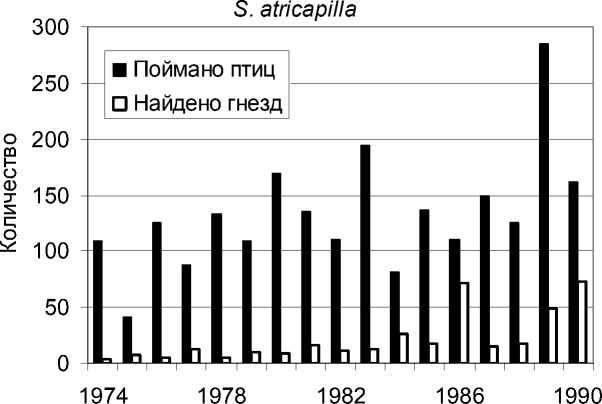
Рис. 3. Количество отловленных особей и найденных гнёзд у славки-черноголовки.
1974 1978 1982 1986 1990
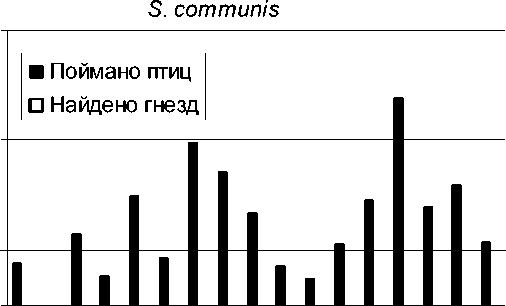
Рис. 4. Количество отловленных особей и найденных гнёзд у серой славки.
S. borin
-
■ Поймано птиц
-
□ Найдено гнезд
ПШ1Ш1ШШ!
1974 1978 1982 1986 1990
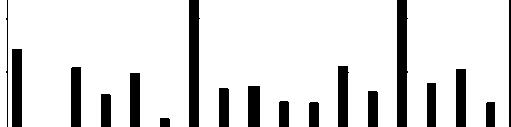
Рис. 4. Количество отловленных особей и найденных гнёзд у садовой славки.
Таблица 4. Тенденции изменения численности славок за 1974-1990годы по данным отлова и по количеству найденных гнёзд*
|
Показатели |
S. atricapilla |
S. borin |
S. communis |
S. curruca |
S. nisoria |
|
Ежегодное общее количество пойманных птиц |
|||||
|
Пределы |
42-285 |
66-603 |
44-239 |
177-493 |
4-159 |
|
Среднее |
135 |
203 |
125 |
371 |
45 |
|
τ |
+ 0.415* |
- 0.022 |
+ 0.272 |
+ 0.022 |
- 0.852*** |
|
r |
+ 0.543* + 0.208 + 0.401 Количество взрослых местных |
+ 0.157 птиц |
- 0.842 *** |
||
|
Пределы |
3-29 |
1-16 |
1-19 |
25-35 |
4-110 |
|
Среднее |
11 |
7 |
11 |
39 |
29 |
|
τ |
+ 0.260 |
+ 0.243 |
+ 0.084 |
- 0.111 |
- 0.739*** |
|
r |
+ 0.151 + 0.410 Количество молод |
+ 0.266 - 0.181 ых местных птиц |
- 0.786 ** |
||
|
Пределы |
1-80 |
1-40 |
2-26 |
35-268 |
0-46 |
|
Среднее |
16 |
17 |
11 |
162 |
15 |
|
τ |
+ 0.622*** |
+ 0.537** |
+ 0.463* |
+ 0.044 |
- 0.682** |
|
r |
+ 0.678** + 0.687** + 0.589* + 0.016 Количество найденных гнёзд |
- 0.805*** |
|||
|
Пределы |
4-73 |
8-57 |
0-11 |
5-99 |
0-66 |
|
Среднее |
21 |
20 |
3 |
33 |
22 |
|
τ |
+ 0.736*** |
+ 0.543** |
+ 0.393* |
- 0.274 |
- 0.929*** |
|
r |
+ 0.711** |
+ 0.634* |
+ 0.494 |
- 0.366 |
- 0.940*** |
* – коэффициент ранговой корреляции Кендэла τ и коэффициент корреляции временных рядов r , уровни значимости: * – <0.05, ** – <0.01, *** – <0.001.
В эти годы статистически стабильной оказалась численность славки-завирушки, а катастрофическое снижение численности наблюдалось у ястребиной славки. Статистически значимый популяционный рост отмечен у черноголовой, садовой и серой славок – и по данным отлова молодых птиц, и по количеству найденных гнёзд.
Самым впечатляющим результатом нашего анализа явилось несомненное сходство тенденций, выявленных по исходным данным разного типа: по общему количеству пойманных птиц, по количеству только местных взрослых, по количеству только местных молодых, по количеству найденных гнёзд.
Сравнение изменений численности в разных странах, близких к Куршской косе (Польша, Швеция, Дания), показало сходный тип популяционных тенденций для многих видов птиц, в том числе и для славок. Основной предполагаемый в те годы фактор, вызывающий изменения численности – антропогенное воздействие, главным образом загрязнение среды ядохимикатами. Возможно, что у ряда видов птиц в годы особенно интенсивного использования ДДТ и других химических препаратов в сельском и лесном хозяйствах произошло со- вмещение этого влияния с периодом естественного спада численности при циклических её изменениях. Важен и другой аспект – конкуренция между близкими видами. Вполне возможно, что снижение численности доминировавших в конкуренции видов может привести к росту численности других видов, для которых положительный эффект ослабления конкуренции важнее, чем воздействие химического загрязнения.
Заключение
Сопоставление показателей изученных популяций славок Куршской косы с аналогичными показателями других популяций показало отсутствие во многих случаях каких-либо заметных расхождений. Размеры и масса тела, величина кладок были в пределах значений, указанных для европейских популяций. То же самое можно сказать и об основных чертах биологии славок (Cramp et al . 1992). Тем не менее, детальный сбор и анализ популяционных параметров на протяжении многих лет изучения обычных видов птиц даёт возможность не только получать непреходящей ценности фактические данные, но и обсуждать ряд общебиологических закономерностей, таких, например, как соотношение выживаемости и возрастно-половой структуры популяций, влияния изменений погоды и климата на изменения численности птиц и т.п.
