Биология редкого вида - курильского чая (Pentaphylloides fruticosa) - в культуре на Севере
 - в культуре на Севере")
Автор: Мифтахова С.А., Скроцкая О.В., Зайнуллина К.С.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 2 (30), 2017 года.
Бесплатный доступ
Изучены биологические особенности образцов разного географического происхождения редкого и ценного лекарственного, декоративного растения Pen- taphylloides fruticosa (курильский чай кустарниковый). Показано, что анатомическое строение листовой пластинки P. fruticosa характеризует его как типично мезоморфное и светолюбивое растение. Установлено, что длительность онтогенетических состояний у изучаемых особей курильского чая в культуре на Севере сокращается. Отмечено промежуточное онтогенетическое состояние, когда у растений первого года жизни, имеющих признаки виргинильного онтогенетического состояния, наблюдается цветение. Дана генетическая характеристика коллекционных образцов P. fruticosa.
Биологические особенности, генетическая характеристика, среднетаежная подзона республики коми
Короткий адрес: https://sciup.org/14992887
IDR: 14992887 | УДК: 582.711.71:631.526(470.13)
Biology of the rare species of Pentaphylloides fruticosa (Rosaceae) in culture in the North
The purpose of this work was studying the biology of development, definition of a pool of flavonoids in medicinal raw materials and the genetic characteristics of the introduced plants of different samples of P. fruticosa (L.) O.Schwarz in conditions of the middle taiga subzone of the Komi Republic. Objects of research work are plants of five samples of P. fruticosa of various geographical origin (Gorno-Altaysk - a natural sample, Novosibirsk (The Central Siberian botanical garden), three samples of local reproduction of different age) and two samples collected in the Inta region of the Komi Republic on the right and left banks of the river Lemva. The standard modern methods of research were used: morpho- logical, ontogenetic, anatomic, genetic, biochemical. It is established that dura- tion of the ontogenetic states of plants in conditions of culture is reduced in comparison with natural ones. At the same time the intermediate ontogenetic state is noted, when along with blossoming there are signs of a virgin ontogenet- ic state. Features of structure of a flower are revealed and the process of polli- nation of the introduced samples is traced. Plants annually form full seeds, that speaks about resistance of their reproductive organs to new soil climatic condi- tions and the ability to self-renewal. It is shown that the anatomic structure of a sheet plate of P. fruticosa characterizes the plant as typically mesomorphic and photophilous. This information is useful for development of methods of its cul- tivation. For the purpose of studying the intraspecific diversity of collection samples of P. fruticosa the genetic analysis was carried out. The obtained ma- terial is necessary for further work on specification of taxonomical structure of the genus Dasiphora, including in the region.
Текст научной статьи Биология редкого вида - курильского чая (Pentaphylloides fruticosa) - в культуре на Севере
Коллекции генетических ресурсов растений, в том числе редких и исчезающих видов, сохраняемых в ботанических садах, имеют большое значение, являясь основой для научных исследований. Их создание и сохранение считается необходимым условием надежной консервации и эффективного использования генетических ресурсов. Изучение этих видов на молекулярно-генетическом уровне позволит решить главную проблему в сохранении биоразнообразия – отбор наиболее типичных представителей популяций, а также таксономическую дифференциацию объектов исследования [1], но вместе с тем и привлечение образцов из популяций растений со специфичными характеристиками генофондов [2]. Анализ ДНК напрямую раскрывает геном и может дать устойчивые характеристики растения, нейтральные по отношению к среде обитания и пригодные для идентификации генотипов, регистрации и маркирования хозяйственно-ценных генов и признаков [3]. Ключевую роль в этом играют оценка биоразнообразия, представленного в коллекции, а также характер и уровень генетического разнообразия растений каждого образца коллекции.
На сегодняшний день актуальной остается проблема получения лекарственного сырья, особенно для северных районов России. Одним из ценных сырьевых растений, всесторонне изучаемых в настоящее время как кормовое, лекарственное и декоративное, является Pentaphylloides fruti-cosa (L.) O.Schwarz – курильский чай кустарниковый – из семейства Rosaceae Juss. реликтовое редкое растение северо-востока европейской части России. Он включен в Красную книгу Республики Коми [4], имеет статус 2(V) – редкий уязвимый вид с сокращающейся численностью. Вид произрастает только в Северном полушарии и имеет дизъюнктивный ареал, состоящий из азиатской, европейской и североамериканской частей [5]. Для более активного введения в культуру P. fruticosa недостаточно изучены некоторые вопросы его биологии (онтогенез, процессы репродукции и др.).
Целью настоящих исследований являлось изучение биологии развития и генетическая характеристика интродуцированных растений разных образцов P. fruticosa в условиях среднетаежной подзоны Республики Коми.
Материал и методы
Исследования проводились в Ботаническом саду Института биологии Коми научного центра УрО РАН, расположенном в 8 км к югу от г. Сыктывкара (62º с.ш., 50º в.д.) – подзона средней тайги. Климатические условия в районе исследований весьма суровые. Сезоны года отличаются большой неравномерностью по продолжительности, наиболее длительным является зимний. Вегетационный период начинается в последней декаде апреля, когда среднесуточная температура воздуха становится выше +5ºС. Его продолжительность составляет 150 дней, сумма суточных температур за этот период – 1800ºС. Продолжительность периода активной вегетации со среднесуточными температурами +10ºС изменяется от 90 до 110 дней, сумма суточных температур в этот период – 1450ºС. Территория относится к достаточно увлажненному району, среднегодовая сумма осадков – 500–600 мм, из которых 400–450 мм выпадает в теплый период года [6]. Почвы на участке Ботанического сада дерново-подзолистые, глееватые, среднеокульту-ренные, суглинистого механического состава. Материалом для исследования (в 2007 – 2015 гг.) послужили растения пяти образцов P. fruticosa различного географического происхождения (ГорноАлтайск – природный образец, Новосибирск (Центральный сибирский ботанический сад – ЦСБС), три образца местной репродукции разного возраста (четвертого, седьмого и десятого годов жизни) и двух образцов, собранных в Интинском районе Республики Коми на правом и левом берегах р.Лемвы, где для сохранения основной части популяции данного вида организован памятник природы
«Лемвинский» (самое северное островное местонахождение растений P. fruticosa в Европе).
Определение возрастных состояний проводили согласно классификации Т.А.Работнова [7], дополненной методиками А.А.Уранова [8] и Л.А.Жуковой [9]. При описании цветка руководствовались «Атласом по описательной морфологии высших растений. Цветок» [10]. Для изучения анатомо-морфологических признаков листа P. fruticosa фиксировали листья в полевых условиях в период цветения (конец июля). Определение количественных показателей проводили в лабораторных условиях, на временных препаратах, сделанных при помощи вибрационного микротома для мягких тканей [11]. Изучали препараты с использованием микроскопа ZEISS Axiovert 200 м. Генетический анализ растительных проб образцов P. fruticosa проведен сотрудниками Центра коллективного пользования «Молекулярная биология» Института биологии Коми НЦ УрО РАН. Степень идентичности определялась как доля (в процентах) одинаковых нуклеотидов, занимающих одни и те же позиции у каждой из сравниваемых нуклеотидных последовательностей. За высокую степень идентичности принимали 99–100%-ный уровень.
Результаты и обсуждение
P. fruticosa – сильно ветвящийся невысокий кустарник, 20–150 см высоты, шаровидной формы с перистыми желтовато-зелеными листьями, состоящими обычно из пяти, реже трех–семи ланцетных, волосистых с обеих сторон листочков. Ветви покрыты серо-бурой отслаивающейся корой. Молодые побеги цилиндрические, опушенные. Золотисто-желтые цветки обычно в щитках или небольших рыхлых верхушечных кистях. Плоды состоят из отдельных сухих волосистых семянок, иногда серповидно изогнутых. Растет на хорошо освещенных местах по берегам рек, на лугах, в кустарниковых зарослях и на каменистых склонах в центральных областях европейской части России, на Кавказе, в Сибири и на Дальнем Востоке. Встречается в Западной Европе, Китае, Монголии, Японии и в Северной Америке [12]. В Республике Коми произрастает на Приполярном Урале – верхнее и среднее течение рек Щугер, Кожим и Лемва [4].
P. fruticosa хорошо размножается как семенами, так и черенкованием, достаточно зимостоек [13]. Для изучения онтогенеза семена местной репродукции образца из Горно-Алтайска в мае предварительно высевались в теплицу для получения рассады. В имматурном онтогенетическом состоянии растения были перенесены в открытый грунт. Длительность онтогенетических состояний у особей P. fruticosa в культуре сокращается по сравнению с природными условиями. Описание его онтогенеза в природе в Горном Алтае сделано Л.М. Шафрановой [14]. Е.К. Комаревцевой [15] была определена длительность онтогенетических состояний: p – до года, j – 2–4 года, im – 3–5 лет, v – 20–30 лет. При интродукции в среднетаежную подзону Республики Коми их продолжительность значительно сократилась: p – 11 – 13 дней, j – 20–30 дней, im – 20–30 дней, в виргинильное онтогенетическое состояние (v) боль- шая часть растений переходит в середине сентября и находится в нем до середины июля следующего вегетационного периода (второго года жизни). В природе P. fruticosa зацветает в возрасте 12–20 лет [15]. Нами установлено, что в культуре в первый год жизни особей цветение наступает, когда вегетативная сфера еще не совсем сформировалась. Наблюдается промежуточное состояние, когда наряду с появлением генеративных органов остаются признаки, характерные для растений виргинильного онтогенетического состояния. Основная масса растений вступает в генеративный период со второго года жизни. Вегетация на второй и последующие годы жизни начинается в начале мая. Цветение продолжительное и обильное – с последней декады июня и до октября. Цветки в диаметре до 2,9 см. У растений третьего года жизни до пяти – семи осей возобновления, к шестому году, когда кусты значительно разрастаются, их уже насчитывается до 20–25. На трех- и шестилетних растениях до 147 и 590 цветков на одной оси соответственно. В течение всего периода цветения куст выглядит очень декоративно, поэтому P. fruticosa рекомендуется для использования в зеленом строительстве. Растения ежегодно формируют зрелые семена, сбор которых проводится с середины августа.
Для P. fruticosa характерен половой полиморфизм. Известно, что данный вид образует цветки трех типов: мужские, женские и обоеполые. По степени половой дифференциации в естественных условиях Горного Алтая выделены следующие популяции: мономорфные (состоящие из особей с обоеполыми цветками) и гетероморфные, куда входят либо только мужские и женские (диэцичные це-нопопуляции), либо мужские, женские и обоеполые особи (триэцичные ценопопуляции). Любая особь имеет цветки только одного полового типа [16]. Однако сведения по изучению процессов репродукции Р. fruticosa на Северо-Востоке европейской части России немногочисленны. Объектом для исследований послужил образец P. fruticosa , привлеченный из Горно-Алтайска. Изучение структуры соцветий – одна из самых сложных областей морфологии. Исследователи, изучавшие этот вид в природе, отмечают, что цветки собраны в небольшие рыхлые верхушечные кисти или щитки [17], или в немногоцветковые соцветия [15]. Наблюдения за растениями P. fruticosa в условиях интродукции в среднетаежной подзоне Республики Коми показали, что в первые годы жизни они образуют в основном единичные цветки. В последующие годы – соцветия сложные в многоярусных дихазиях, очень редко -одиночные цветки. Главная ось заканчивается цветком (закрытые соцветия), также как и боковые оси обладает ограниченным ростом (монотелические соцветия).
Растения среднего и старого генеративного онтогенетических состояний завершают цветение уже к началу сентября. Распускание цветков в соцветии происходит в течение длительного периода, и они находятся на разных этапах развития. При этом бутоны в основании соцветия могут не распус- титься в течение периода вегетации. Цветок за счет удлинения цветоножки всегда оказывается выше бутонов или уже отцветших цветков. Продолжительность цветения отдельного цветка - от одного до пяти дней. Цветки растений разных образцов P.fruticosa, выращиваемых в Ботаническом саду, в диаметре от 2,6 до 3,4 см, обоеполые, актиноморфные, в основном пентамерные (рис. 1), гетерохламидные (рис. 2). Чашечка двойная раздельнолистная, подпестичная, зеленого цвета неопушенная.

Рис.1. Цветок Pentaphylloides fruticosa. Fig. 1. Pentaphylloides fruticosa flower.

Рис. 2. Двойная чашечка (лепестки удалены).
Fig. 2. A double cup (petals are removed).
Чашелистики внешнего круга продолговато-линейные с заостренной вершиной, 4,5 – 7,7 мм длиной и 1 – 2,3 мм шириной. Венчик состоит из внешнего и внутреннего кругов, несущих обычно по пять лепестков желтого цвета. Внутренние лепестки треугольные или ланцетовидные, желтовато-зеленые 6,6 – 7,9 мм длиной и 3,2 – 4,7 мм шириной. Лепестки гладкие, округлые или яйцевидные длиной 8,2–13,2 мм и шириной 9,1–14 мм. Венчик опадает после отцветания цветка, чашечка, увядая, остается при плоде. Двойной околоцветник и тычинки срастаются с цветоложем и образуют блюдцевидный гипантий. Андроцей свободный, состоит из 22– 33 прямых и отогнутых тычинок (рис. 3), прикреплен-
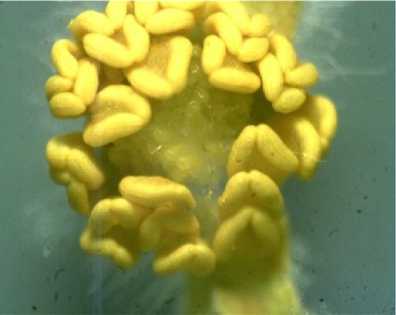
Рис. 3. Андроцей и гинецей Pentaphylloides frutico-sa в закрытом бутоне перед опылением.
Fig. 3. Androecium and gynoecium Pentaphylloides fruticosa in closed bud before pollination.
ных основанием, неравных, длиннее гинецея, подпестичных. Тычиночная нить цилиндрическая, голая, длиной 1,1 – 2,7 мм. Связник (треугольный или яйцевидный) продолжает тычиночную нить. Пыльники по длине не равные, короче тычиночных нитей, сердцевидной или яйцевидной формы. Они достигают максимального размера перед опылением в закрытом бутоне. При их раскрытии пыльца попадает внутрь цветка. Гинецей апокарпный, из 34 – 62 пестиков (рис. 4). Пестик представлен одним плодолистиком. Он состоит из верхней, одногнездной завязи яйцевидной формы, стилодия и рыльца с густым опушением. После отцветания увядшие сти-лодии остаются на завязи.

Рис. 4. Гинецей Pentaphylloides fruticosa.
Fig. 4. Gynoecium Pentaphylloides fruticosa.
Для P.fruticosa характерен утренний максимум раскрытия цветков, приходящийся на время восхода солнца. Цветки распускаются однократно, цветут от одного до пяти дней в зависимости от метеоусловий. Согласно данным литературы, P.fru-ticosa относится к энтомофилам. Желтая окраска лепестков – одно из приспособлений к ксеногамно-му опылению насекомыми. В Горном Алтае цветки посещают 56 видов насекомых из пяти отрядов – перепончатокрылых (17 видов), двукрылых (28 видов), полужесткокрылых (3 вида), жесткокрылых (6
видов) и сетчатокрылых (2 вида), 39 видов отнесены к опылителям [18].
В 2011 г. проведено наблюдение за опылением в цветках P.fruticosa . Установлено, что высыпание пыльцы происходит в бутонах (клейстогамия), она высыпается из пыльников и оседает на рыльце (рис. 5), которое к этому моменту увлажняется. В цветках нет временного разобщения в функционировании мужских и женских генеративных органов (одновременное созревание пыльников и рылец) – адихогамное развитие. Явление клейстогамии представляет крайнюю форму автогамии и встречается довольно редко, если не является обязательным. В основном клейстогамия вы-

Рис. 5. Опыление в закрытом бутоне.
Fig. 5. Pollination in closed bud.
зывается неблагоприятными условиями внешней среды. Наблюдения проведены в разные периоды вегетационного сезона (в середине июля и начале октября). В июле, в период сухой жаркой погоды, ярко окрашенный венчик растений привлекал насекомых, которые активно посещали цветки в течение всего дня. В начале октября опыление цветков насекомыми было минимальным. Однако, чтобы подтвердить явление клейстогамии необходимо проводить дополнительные исследования.
Наличие многопорядковых соцветий, высокое обилие цветков и их последовательное цветение в течение длительного периода приводят к образованию фертильных семян и самосеву у P.fru-ticosa. Способность интродуцентов к размножению и самовозобновлению служит одной из характеристик успешности интродукции, а устойчивость репродуктивных органов растений к неблагоприятным факторам внешней среды является важнейшим показателем онтогенетической адаптации.
Лист растения в наибольшей степени связан с окружающей средой, поэтому его строение сильнее отражает изменения условий среды. Внешнее разнообразие листьев сопровождается таким же разнообразием их анатомического строения, которое связано как с таксономическим положением растений, так и с приспособлением их в процессе эволюции к различным местообитаниям [19]. Размеры листовой пластинки P.fruticosa в период цветения от 2 до 3 см длиной и от 3 до 3,5 см шириной. Ее толщина 10–16 мкм. Покровная ткань представ- лена эпидермой с утолщенными наружными стенками клеток. Эпидерма одно-двухслойная, из крупных клеток неправильной формы. Их размеры на адаксиальной стороне от 2,25 до 1,2 мкм, на абаксиальной – от 1,0 до 0,7 мкм. Эпидерма с обеих сторон имеет волоски, причем с адаксиальной стороны листа их меньше. Между верхним и нижним эпидермисом находится основная фотосинтезирующая ткань – мезофилл, которая дифференцирована на палисадную и губчатую. Свет – ведущий экологический фактор, влияющий на нормальное развитие и функционирование фотосинтетического аппарата. Для выявления действия светового фактора на формирование ассимиляционного аппарата основное внимание уделяется толщине мезофилла, развитию палисадной и губчатой ткани. Мезофилл листа у растений P. fruticosa – дорсивент-ральный, что характерно для мезоморфных растений. Палисадная паренхима хорошо выражена и состоит из двух–трех слоев клеток, которые вытянуты в направлении, перпендикулярном поверхности эпидермиса, и имеют призматическую форму. Размеры столбчатого мезофилла от 4,2 до 6,5 мкм, губчатого от 2,7 до 3,7 мкм. Губчатая паренхима имеет высокий парциальный объем межклетников. Структурный тип листа представляет достаточно консервативный признак и не затрагивается при изменении экологических условий произрастания растений, меняются только его отдельные показатели, такие как толщина листа, мезофилла, коэффициент палисадности [20]. Анатомическое строение листовой пластинки P. fruticosa характеризует его как типично мезоморфное и светолюбивое растение, что следует учитывать при культивировании ценного лекарственного и декоративного растения.
Дана генетическая характеристика коллекционных образцов P. fruticosa . Получены последовательности рибосомальной ДНК (общая длина ITS-участков варьировала от 585 до 600 п.н.). Анализ сходства ITS-фрагментов рибосомальной ДНК коллекционных образцов P. fruticosa Ботанического сада Института биологии с идентичными последовательностями, депонированными в базе данных GenBank [21], показал, что все представленные образцы имеют общую видовую принадлежность к Potentilla fruticosa (синонимы: Pentaphylloides fru-ticosa , Dasiphora fruticosa , Dasiphora fruticosa var. albicans ) рода Dasiphora L. ( Potentilla ) (см. таблицу). Степень идентичности полученных сиквенсов соответствует 99 –100%-ному уровню последовательности нуклеотидов из NCBI-GenBank. Отмечена высокая гомология (99 %) последовательностей природного образца P. fruticosa , полученного из ГорноАлтайска, и коллекционных экземпляров ЦСБС с нуклеотидными последовательностями видов Dasi-phora davurica , Dasiphora glabra и Dasiphora phyllocalyx . В базе данных последовательность природного образца P. fruticosa из Интинского района Республики Коми имеет гомологи по шести сиквен-сам вида Potentilla fruticosa ( Dasiphora fruticosa , Pentaphylloides fruticosa ), а также вида Dasiphora davurica со степенью сходства в 99 %. Полученные
Результаты анализа 99 %-го сходства ITS-последовательностей между коллекционными образцами Ботанического сада ИБ и идентичными образцами, зарегистрированными в базе данных GenBank
The analysis of 99% similarity of ITS-sequences between collection samples of the Botanical Garden of the Institute of Biology and identical samples registered in the GenBank database
|
Виды рода Pentaphylloides Hill в базе данных GenBank |
Образцы Pentaphylloides fruticosa в коллекции Ботанического сада Института биологии |
||
|
Горный Алтай (природный образец) |
ЦСБС |
Интинский р-н, Республика Коми (природный образец) |
|
|
Р. fruticosa |
+ |
+ |
+ |
|
Р. davurica |
+ |
+ |
+ |
|
Р. glabra |
+ |
+ |
- |
|
Р. phyllocalyx |
+ |
+ |
- |
(«+» — есть сходство, «-» — нет сходства) (“+” – similarity, “-“ – no similarity результаты по рибосомальным ITS последовательностям в дальнейшем послужат материалом для составления филогении у Dasiphora L., а также являются новыми дополнительными информативными маркерами для занесения их в базу данных NCBI-GenBank.
Заключение
Изучены биологические особенности растений образцов P.fruticosa разного географического происхождения при интродукции в среднетаежной подзоне Республики Коми. Установлено, что продолжительность онтогенетических состояний у особей в условиях культуры сокращается по сравнению с природными. Так, если по данным Е.Л. Ко-марцевой (2005) растения P.fruticosa в естественных местообитаниях вступают в фазу цветения на 12 – 20-е годы жизни, то при интродукции на Севере это может происходить в первый год жизни. При этом отмечено промежуточное онтогенетическое состояние, когда, наряду с цветением, имеются признаки виргинильного онтогенетического состояния. В результате сравнительного морфологического анализа выявлены особенности строения цветка и прослежен процесс опыления интродуцируемых образцов. Растения изученных образцов ежегодно формируют фертильные семена, что говорит о возможности интродуцентов к самовозобновлению и устойчивости их репродуктивных органов к новым почвенно-климатическим условиям. Это является одним из важнейших показателей онтогенетической адаптации. Отмечено, что анатомическое строение листовой пластинки P. fruticosa характеризует растение как типично мезоморфное и светолюбивое, что является необходимой информацией для разработки приемов его выращивания.
С целью изучения внутривидового разнообразия коллекционных образцов P. fruticosa проведен их генетический анализ. Сравнение последовательностей их ITS-фрагментов рибосомной ДНК с данными NCBI-GenBank установлено, что все ис- следуемые растения идентифицируются на 99 – 100%-ном уровне как P. fruticosa, природный образец P. fruticosa из Горно-Алтайска и коллекционные экземпляры ЦСБС гомологичны (99%) Dasiphora davurica, Dasiphora glabra и Dasiphora phyllocalyx. Природный образец P. fruticosa из Интинского района Республики Коми имеет высокую степень генетического родства с видом Dasiphora davurica. Полученный материал необходим для дальнейшей работы по уточнению таксономического состава рода Dasiphora, в том числе и в регионе.
Работа выполнена в рамках государственного задания по теме «Некоторые аспекты репродуктивной биологии ресурсных видов растений в культуре на европейском Северо-Востоке России» № 115012860039.
Авторы выражают искреннюю благодарность научным сотрудникам Центра коллективного пользования «Молекулярная биология» Института биологии Коми НЦ УрО РАН к.б.н. Е.А.Юшковой, к.б.н. И.С.Боднарь, к.б.н. Д.М.Шадрину за выполнение генетической характеристики коллекционных образцов Pentaphylloides fruticosa.
Список литературы Биология редкого вида - курильского чая (Pentaphylloides fruticosa) - в культуре на Севере
- Оценка молекулярно-генетического полиморфизма редких и охраняемых видов растений Республики Беларусь/О.В.Дзюбан, З.Е.Грушецкая, В.Н.Тихомиров, В.Д.Поликсенова, В.И.Парфенов//Актуальные проблемы генетики и молекулярной биологии в рамках фестиваля науки: Тез. докладов Всерос. молодежной конференции «Научные и научно-педагогические кадры инновационной России». Уфа, 2012. 68 с
- Боронникова С.В. Молекулярно-генетический анализ генофондов редких и исчезающих видов растений Пермского края: Автореф. дис. … доктора биол. наук. Уфа, 2009. 44 с
- Павловская Н.Е., Гагарина И.Н., Прудникова Е.Г. Применение ПЦР-метода для маркирования сельскохозяйственных растений//Вестник ОрелГАУ. 2009. Т.18. №3. С. 32-35
- Красная книга Республики Коми. Сыктывкар, 2009. 791 с
- Юзепчук C.В. Лапчатка -Potentilla L.//Флора СССР. М.; Л., 1941. Т. 10. С. 68-223
- Атлас по климату и гидрологии Республики Коми. М., 1997. 116 с
- Работнов Т.А. Методы определения возраста и длительности жизни у травянистых растений//Полевая геоботаника. М.; Л., 1960. Т.2. 500 с.
- Уранов А.А. Онтогенез и возрастной состав популяций//Онтогенез и возрастной состав популяций цветковых растений. М., 1967. С. 3-8
- Жукова Л.А. Популяционная жизнь луговых растений. Йошкар-Ола, 1995. 224 с
- Федоров А.А., Артюшенко З.Т. Атлас по описательной морфологии высших растений. Цветок. Л., 1975. С.177
- Скупченко В.Б. Вибрационная микротомия мягких тканей. Сыктывкар, 1979. 56 с. (Сер. препр. сообщений «Новые научные методики»/Коми филиал АН СССР; Вып. 2)
- Деревья и кустарники СССР/Под ред. П.И.Лапина. М., 1966. С. 395
- Скупченко Л.А., Мишуров В.П., Волкова Г.А., Портнягина Н.В. Интродукция полезных растений в подзоне средней тайги Республики Коми (Итоги работы Ботанического сада за 50 лет; Т. III). СПб., 2003. 214 с
- Шафранова Л.М. Жизненные формы и морфогенез Potentilla fruticosa L. в различных условиях произрастания//Бюл. МОИП. 1964. Т. 71. Вып. 4. С. 101-110
- Комаревцева Е.К. Онтогенез и структура популяций Pentaphylloides fruticosa (Rosaceae) Горного Алтая//Растит. ресурсы. 2005. Т. 41. Вып. 1. С. 27-34
- Годин В.Н. Половая структура ценопопуляций Pentaphylloides fruticosa (Rosaceae) в естественных условиях Горного Алтая//Ботан. журнал. 2002. Т. 87. № 9. С.92-99
- Иванова З.Я. Курильский чай. М., 2005. 56 с
- Годин В.Н. Половой полиморфизм как фактор адаптации Pentaphylloides fruticosa (L.) O. Schwarz в Алтае-Саянской горной области: Автореф. дис.... доктора биол. наук. Новосибирск, 2009. 31 с
- Ботаника с основами фитоценологии: Анатомия и морфология растений/Т.И.Серебрякова, Н.С.Воронин, А.Г.Еленевский, Т.Б.Батыгина, Н.И.Шорина, Н.П.Савиных. М., 2007. 543 с
- Оганезова Г.Г. Экологический тип листа и история происхождения некоторых лилейных//Современные проблемы экологической анатомии растений: Материалы I Всесоюз. совещ. по экологической анатомии растений. Ташкент, 1987. С. 66-69
- GenBank Home. 2015; http://www.ncbi.nlm. nih.gov/genbank/
