Биоморфологические аспекты естественного возобновления Quercus robur L. на песчаных отложениях в долине р. Большой Черемшан (Ульяновская область)
")
Автор: Стаменов М.Н.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 4 т.19, 2025 года.
Бесплатный доступ
С использованием современных биоморфологических подходов исследовано строение кроны у молодых особей Quercus robur L., произрастающих на выходах песков с редкой травянистой растительностью на склоне надпойменной террасы р. Большой Черемшан (Мелекесский район Ульяновской области). Всего изучено 106 деревьев. Локус имматурных особей нормальной и пониженной жизненности расположен в верхней части склона вблизи опушки леса. В остальных частях склона имматурные растения встречаются крайне редко и имеют пониженную и низкую жизненность. Виргинильные и молодые генеративные особи произрастают вдоль всего склона, и имеют в основном нормальную жизненность. В имматурном онтогенетическом состоянии преобладают растения с ортотропным стволом и редкими восходящими ветвями. Строение кроны в виргинильном и молодом генеративном состояниях сводится к трем типам. 1 – ортотропный ствол и восходящие ветви, 2 – ортотропный ствол и преимущественно плагиотропные ветви, 3 – переходящий от ортотропного к плагиотропному направлению роста ствол, плагиотропные и восходящие ветви. Преобладают особи типа 1. У большинства особей всех онтогенетических состояний в приземной части ствола сформирован комплекс осей, включающий один или несколько следующих элементов: либо плагиотропные/восходящие ветви, либо стелющиеся по поверхности субстрата оси, либо дополнительные стволы, уступающие по развитию основному. У двух имматурных особей выявлен ксилоподий.
Экологическая морфология, дуб черешчатый, крона, ствол, ветви, ксилоподий, надпойменная терраса, пески
Короткий адрес: https://sciup.org/148332081
IDR: 148332081 | УДК: 581.44/.52+582.632.2 | DOI: 10.24412/2072-8816-2025-19-4-227-237
Biomorphological aspects of natural regeneration of Quercus robur L. on sandy sediments in the valley of the river Bolshoi Cheremshan (Ulyanovsk region)
Using modern biomorphological approaches, the crown structure of young Quercus robur L. specimens growing on sand outcrops with sparse herbaceous vegetation on the slope of the floodplain terrace of the Bolshoy Cheremshan River (Melekessky District, Ulyanovsk Region) was studied. A total of 106 trees were studied. The locus of immature specimens of normal and low vitality is located in the upper part of the slope near the forest edge. In other parts of the slope, immature plants are extremely rare and have low and low vitality. Virginile and young generative specimens grow along the entire slope and have mostly normal vitality. In the immature ontogenetic state, plants with an orthotropic trunk and rare ascending branches predominate. The crown structure in the virginile and young generative states is reduced to three types. 1 – orthotropic trunk and ascending branches, 2 – orthotropic trunk and predominantly plagiotropic branches, 3 – trunk transitioning from orthotropic to plagiotropic growth direction, plagiotropic and ascending branches. Type 1 individuals predominate. Most individuals of all ontogenetic stages have a complex of axes in the ground part of the trunk, including one or more of the following elements: either plagiotropic/ascending branches, or axes creeping along the surface of the substrate, or additional trunks inferior in development to the main one. Xylopodium was found in two immature individuals.
Текст научной статьи Биоморфологические аспекты естественного возобновления Quercus robur L. на песчаных отложениях в долине р. Большой Черемшан (Ульяновская область)
Среди лесообразующих видов Европейской части России особенно широкой экологической амплитудой характеризуется дуб черешчатый (Quercus robur L.) (Lositskiy, 1981). Его естественный ареал охватывает огромное пространство от южной тайги до полупустынь. Популяции вида существуют в поймах рек, на надпойменных террасах речных долин, в балках и на водоразделах. Лесоводы выделяют у Q. robur целую серию экотипов и экологических форм, связанных с определенными климатическими и почвенно-гидрологическими условиями (Selektsiya…, 1982). Для лучшего понимания адаптивных возможностей видов и прогнозирования динамики их популяций особенно важно исследовать морфологические девиации в условиях, которые существенно отличаются от оптимума. Так, для Q. robur в ряду благоприятности местообитаний по почвенному богатству одно из краевых положений занимают сообщества на песчаных и супесчаных отложениях (Lositskiy, 1981; Selektsiya…, 1982; Novosel’tsev, Bugaev, 1985). В сосняках и сосново-широколиственных лесах надпойменных террас рек и зандровых равнин Q. robur присутствует в различных ярусах древостоев (Zakamskaya et al., 2012; Grishutkin, 2014; Suslova, 2019; Evstigneev, Korotkova, 2024). Гораздо реже распространены дубовые леса на массивах песков за пределами лесной зоны. Такие дубравы получили название аренных. Наиболее крупные и хорошо описанные площади аренных дубовых лесов расположены на севере Ростовской области (Sokolova, 2019).
Представляется особенно интересным исследовать потенциал развития Q. robur на песчаных субстратах с минимальным развитием плодородного слоя почвы. Обследуя в октябре 2024 г. сообщества долины р. Большой Черемшан в пределах Ульяновской области, мы обнаружили ценопопуляцию (ЦП) Q. robur , произрастающую на песчаном массиве надпойменной террасы. Поэтому целью нашего сообщения мы поставили краткий обзор количественных и качественных характеристик строения кроны молодых деревьев Q. robur в данной ЦП.
Материалы и методы
Исследования проводили в заволжской части Ульяновской области, вблизи с. Ерыклинск Мелекесского района (рис. 1). Климат района исследований умеренно-континентальный. Годовое количество осадков составляет 420 мм, а гидротермический коэффициент – 0.9, что свидетельствует о недостаточном увлажнении (Antonova, 2015). Район исследований относится к Черемшано-Майнскому почвенно-экологическому району Заволжского низменно-равнинного округа (Antonova, 2015) и Черемшанском лесному ландшафтному району (Priroda…, 1963).
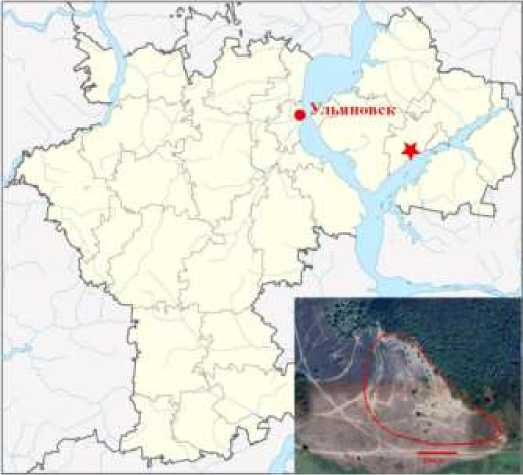
Рис. 1. Район исследований. Звездочкой показано расположение ценопопуляции (ЦП) Quercus robur на карте Ульяновской области, на врезке – спутниковый снимок участка надпойменной террасы р. Большой Черемшан, в котором расположена исследуемая ЦП
Fig. 1. Research area. The location of the Quercus robur coenopopulation (CP) on the map of the Ulyanovsk region is shown by an asterisk; the inset shows a satellite image of the floodplain terrace area of the Bolshoy Cheremshan River, where the studied CP is located
Описываемая ЦП Q. robur расположена на высоте 70–95 м н.у.м и занимает юго-западный склон надпойменной террасы р. Большой Черемшан. Фитоценоз представляет собой зарастающий молодыми особями деревьев и кустарником участок нарушенной песчаной степи (рис. 2). В травяном ярусе преобладает Calamogrostis epigejos (L.) Roth. Его проективное покрытие составляет 80–90% от общей площади, занятой травянистой растительностью. Кроме того, в травостое регулярно встречаются Bromus tectorum L., Artemisia abronatum L., Verbascum lychnitis L., Hieracium sp., Centaurea sp., изредка – Silene tatarica Pers., Linaria vulgaris Mill., Erigeron annuus (L.) Desf.и др. В средней и нижней частях склона местами интенсивно разрастается Chamaecytisus ruthenicus (Fisch. ex Woł.) Klásk. Древесная растительность состоит преимущественно из молодых деревьев Q. robur, которые растут как поодиночке, так и разреженными группами. Во втором случае размер дубовых колков соcтавляет 20–50 м2, сомкнутость крон в пределах колка не превышает 0.3–04. Все деревья полностью освещены. Также на склоне произрастают единичные особи Malus domestica (Suckow) Borkh., Ulmus laevis Pall. и Acer platanoides L. высотой 1–4 м, локусы Betula pendula Roth и Populus tremula L. высотой 2–5 м, сомкнутостью 0.7–0.8 и площадью 10–25 м2. До 40% общей площади сообщества занимают участки незадернованных песков, на которых практически отсутствует травянистая растительность.

Рис. 2. Сообщество с ценопопуляцией Quercus robur
Fig. 2. Plant community with coenopopulation of Quercus robur
Исследовали организацию побеговых систем кроны у 24 имматурных, 54 виргинильных и 28 молодых генеративных особей. Всего изучено 106 деревьев. Онтогенетические состояния устанавливали согласно принятой в популяционной биологии растений методике (Diagnozy…, 1989). Две особи, переходящие в квазисенильное состояние (Smirnova et al., 1984), рассматривали совместно с имматурными особями. У деревьев измеряли высоту, диаметр ствола на уровне почвы или на высоте 1.3 м, радиус кроны по двум проекциям, выборочно – длины годичных побегов верхней части ствола и ветвей различных порядков. Описывали особенности архитектуры побеговых систем (направление роста, нарастание, интенсивность ветвления), а у двух имматурных особей – также и области корневой шейки и основания ствола. Типизировали конструктивную организацию побеговых систем особей с учетом особенностей строения ствола и ветвей от ствола в средней и верхней зонах кроны. Отдельно описывали строение осей в нижней части кроны вблизи субстрата.
Результаты
Влияние почвенно-геоморфологических условий на строение кроны и особенности размещения деревьев в пространстве . У виргинильных и молодых генеративных особей Q. robur какие-либо зависимости между проективным покрытием травостоя, с одной стороны, и пространственной структурой, морфометрическими показателями и строением кроны, с другой стороны, не обнаружены. В то же время имматурные особи произрастают в основном локусами в верхней части склона вблизи опушки широколиственного леса. В средней и нижней частях склона они отсутствуют. Кроме того, две имматурные особи закрепились в стенке небольшого карьера на южной окраине района исследований.
Морфометрические параметры кроны представлены в табл. 1. Видно, что переход в генеративный период онтогенеза не всегда сопровождается существенным увеличением высоты дерева, в отличие от перехода от имматурного к виргинильному онтогенетическому состоянию.
Таблица 1. Морфометрические параметры особей Quercus robur
Table 1. Morphometric parameters of Quercus robur specimens
|
Морфометрические параметры Morphometric parameters |
Онтогенетические состояния Ontogenetic stages |
||
|
Имматурное ( im ) Immature |
Виргинильное ( v ) Virginile |
Молодое генеративное ( g 1 ) Young generative |
|
|
Высота дерева, м Tree height, m |
0.7–1.5 |
2.5–4.5 |
4–6 |
|
Диаметр ствола, см Stem diameter, cm |
1.8–2.5 |
2.5–4 |
6–12 (до 20) |
|
Радиус проекции кроны, м Crown radius, m |
0.4–0.8 |
0.9–1.8 |
1–2.5 |
|
Календарный возраст, лет Calendar age, years |
10–12 |
15–25 |
20–30 |
|
Длины элементарных побегов ствола, см Length of growth units of the trunk, cm |
10–30 |
10–50 |
20–50 |
|
Длины элементарных побегов крупных ветвей от ствола, см Length of growth units of the skeletal branches, cm |
7–15 |
10–30 |
15–35 |
Примечание. Для имматурных особей приведены значения диаметра ствола на уровне субстрата, для виргинильных и молодых генеративных особей – на высоте груди. Радиус проекции кроны устанавливали по длине проекции наиболее крупных ветвей, отходящих от ствола.
Note. For immature individuals, the stem diameter values are given at the substrate level, for virginile and young generative individuals – at breast height. The crown radius was determined by the projection length of the largest branches extending from the trunk.
Жизненность. Виргинильные и молодые генеративные особи обладают преимущественно нормальной жизненностью. Имматурные особи в составе локуса в верхней части склона также в основном имеют нормальную жизненность, а в остальных частях склона – пониженную и низкую.
Конструкция побегового тела у имматурных особей Q. robur представлена тремя типами:
-
1. Ортотропный неустойчиво-моноподиально нарастающий ствол с восходящими ветвями разной силы развития (рис. 3.1) (79% особей).
-
2. Ортотропный симподиально нарастающий ствол с редкими и слабыми ветвями (рис. 3.2) (13% особей).
-
3. Г-образный ствол. Направление роста главной оси особи меняется с ортотропного на близкое к плагиотропному. Ветви отходят от ствола в горизонтальном направлении. Ствол нарастает в целом моноподиально, а ветви – симподиально (рис. 3.3) (8% особей).
У виргинильных и молодых генеративных особей Q. robur выявлено три из четырех ранее описанных типов организации скелетных осей в кроне (Stamenov, 2024):
-
1. Ортотропный ствол и преимущественно восходящие ветви от ствола (68% особей). Базальная часть ветви может расти плагиотропно, но впоследствии она «поднимается» под углом до 30° к стволу (рис. 4. 1). На стволе регулярно формируются ложные мутовки либо ярусы, состоящие из 2–4 ветвей.
-
2. Ортотропный ствол и преимущественно плагиотропные ветви от ствола (26% особей). Дистальная часть ветви может загибаться вниз к поверхности субстрата или выпрямляться. 1–3 ветви в кроне могут отходить от ствола под острым углом. Кроме того, в верхней трети или четверти кроны на стволе могут образовываться восходящие ветви (рис. 4. 2).
-
3. Ствол в форме полудуги или буквы Г (6% особей). В средней части направление роста меняется с ортотропного на наклонное или реже плагиотропное. Ниже области перегиба ствола ветви отходят под прямым углом. На внешней стороне диагонального отрезка ствола направление роста ветвей меняется на восходящее или даже ортотропное. На внутренней стороне ветви сохраняют плагиотропное направление роста (рис. 4.3).
У особей 2-го и 3-го типов ложные мутовки на стволе образуются реже и включают не более трех ветвей.

Рис. 3. Конструкция побегового тела у имматурных особей Quercus robur . 1, 2 и 3 – типы конструкции кроны (см. текст). Высота особей типов 1, 2 и 3 – 1.6 м, 1.1 м и 1.2 м соответственно
Fig. 3. Construction of the shoot body in immature individuals of Quercus robur . 1, 2 and 3 are types of crown construction (see text). The height of individuals of types 1, 2 and 3 is 1.6 m, 1.1 m and 1.2 m, respectively

Рис. 4. Строение кроны у виргинильных и молодых генеративных особей Quercus robur . 1, 2 и 3 – типы конструкции кроны (см. текст). В – вильчатая структура. Высота особей типов 1, 2 и 3 – 4 м, 2.5 м и 2.2 м соответственно
Fig. 4. Crown structure in virginal and young generative individuals of Quercus robur . 1, 2 and 3 – types of crown structure (see text). B – forked structure. The height of individuals of types 1, 2 and 3 is 4 m, 2.5 m and 2.2 m, respectively
У большинства особей ствол несколько раз раздваивается посредством вильчатых структур (рис. 4.1). Дочерние оси могут либо продолжать расти ортотропно, либо дифференцироваться по силе развития. Более сильная ось принимает на себя роль ствола, в то время как более слабая, растущая косо вверх, функционально соответствует мощной ветви. Оси, составляющие основной и дочерние стволы, нарастают неустойчиво-моноподиально (Serebryakov, 1962): после 2–4 лет моноподиального нарастания ось перевершинивается. Ветви от ствола также нарастают главным образом неустойчиво-моноподиально.
Характерной особенностью большинства описанных виргинильных и молодых генеративных деревьев (97%) и всех имматурных особей Q. robur является образование нескольких вариантов побеговых комплексов в нижней части ствола. Зона, в которой формируются данные структуры, составляет 10–15% от высоты всего дерева и простирается вверх по стволу на высоту до 50–70 см:
-
1. Плагиотропные ветви из почек возобновления одного-двух самых нижних годичных побегов ствола (67% особей). Ветви могут отходить от ствола поодиночке либо образовывать своеобразный «диск» из 2–3 «слоев». Ветви обычно расположены выше уровня субстрата не менее, чем на 10–20 см, но могут соприкасаться с ним при перевершиниваниях осей, а также в дистальной части. В то же время у ряда ветвей дистальная часть оси может и выпрямляться. У 77% особей диаметр «диска» не превышает половины высоты дерева. В этом случае ветви включают последовательности побегов с симподиальным и неусточиво-моноподиальным нарастанием (рис. 5.1).
-
2. Стелющиеся по субстрату оси, отходящие из спящих почек в области корневой шейки (20% особей). Обычно образуется 1–3 слабоветвящихся оси с неустойчиво-моноподиальным нарастанием. Длина таких осей редко превышает 50% от высоты дерева (рис. 5.2).
-
3. Восходящие ветви (7% особей). Образуются из почек возобновления самых нижних побегов ствола (рис. 5.3). Базальная часть может расти плагиотропно. У единичных особей могут достигать средней части ствола. Более короткие ветви нарастают преимущественно симподиально, а наиболее длинные – неустойчиво-моноподиально.
-
4. Дополнительные стволы, растущие ортотропно или косо вверх (35%). В первом случае формируется только один ствол. Он нарастает неустойчиво-моноподиально и может достигать 30–40% от общей высоты особи. Во втором случае формируется до 5 – 7 стволов (чаще 1–3), которые редко превышают 25% от высоты дерева и нарастают как неустойчиво-моноподиально, так и симподиально (рис. 5.4).
-
5. Дополнительные стволы в форме арки или буквы Г (13% особей). Образуется обычно 2–3 сводчатых ствола или один Г-образный, до 20–25% от общей высоты особи. Стволы в форме арки чаще нарастают неустойчиво-моноподиально, а в форме буквы Г – симподиально (рис. 5.5).

Рис. 5. Варианты побеговых комплексов в приземной части кроны у виргинильных и молодых генеративных особей Quercus robur. Номера 1–5 соответствуют вариантам, перечисленным в тексте. 6 – сочетание нескольких вариантов побеговых комплексов (плагиотропные ветви и дополнительные стволы в форме буквы Г). С – дополнительный слабый ствол, Вт – плагиотропная ветвь. Длина наиболее протяженных побеговых систем в вариантах 1–6 – 1.4 м, 1.2 м, 0.8 м, 0.7 м, 0.9 м и 1.5 м соответственно
Fig. 5. Variants of shoot complexes in the ground part of the crown in virginile and young generative individuals of Quercus robur . Numbers 1–5 correspond to the variants listed in the text. 6 – a combination of several variants of shoot complexes (plagiotropic branches and additional trunks in the shape of the letter Г). C – additional weak trunk, Bт – plagiotropic branch. The length of the longest shoot systems in variants 1–6 is 1.4 m, 1.2 m, 0.8 m, 0.7 m, 0.9 m and 1.5 m, respectively
У 40% особей комплекс побеговых систем в нижней части ствола состоит только из ветвей, у 2% – только из стелющихся осей вторичного происхождения, у 22% – из добавочных стволов различной конфигурации. У 36% виргинильных и молодых генеративных деревьев и у всех имматурных особей побеговый комплекс состоит из структур различного происхождения (рис. 5.6).
Возможно, что столь широкое распространение сложно организованных побеговых комплексов в нижней части кроны вызвано в том числе функционированием специального органа – ксилоподия. Он представляет собой утолщенный подземный орган, состоящий из верхней части главного корня, гипокотиля и эпикотиля (Smirnova et al., 1984). Мы обнаружили ксилоподий у двух имматурных особей, произрастающих в стенке небольшого песчаного карьера (рис. 6).

Рис. 6. Подземные и надземные органы у имматурных особей Quercus robur . Кс – ксилоподий. Высота надземных частей левой и правой особей – около 0.7 м
Fig. 6. Underground and aboveground organs in immature individuals of Quercus robur . Кс – xylopodium. The height of the aboveground parts of the left and right individuals is about 0.7 m
От верхней части вертикально направленного ксилоподия, имеющего протяженность 7–20 см, отходят две основные оси кроны: ортотропная и плагиотропная. Ортотропная ось нарастает преимущественно моноподиально; от нее отходят либо восходящие, либо плагиотропные ветви. Ветви, растущие косо вверх, перевершиниваются реже, чем горизонтально направленные ветви. Плагиотропная ось часто перевершинивается и образует серию неравновильчатых структур, напоминающих щупальца (рис. 6). От оси отходят мелкие и быстро отмирающие короткие ветви. От ксилоподия также могут отходить более слабые косые оси с редким ветвлением и частыми перевершиниваниями.
Обсуждение
Анализ строения надземной части особей Q. robur показал, что, несмотря на крайне бедный питательными веществами субстрат, крона у молодых деревьев формируется примерно такими же способами, как и в популяциях, произрастающих в более благоприятных почвенногидрологических условиях. Более того, в исследованной ЦП типы конструкции кроны распределены сходным образом с ЦП, описанных в других природных зонах и физикогеографических регионах. Так, преобладающий в рассматриваемой ЦП тип 1 также является основным и в популяциях вида в южной тайге на подзолистых почвах (Stamenov, 2024), и в луговых степях на черноземных почвах (Stamenov, 2023). Таким образом, даже на песках с редкой травянистой растительностью особи Q. robur достаточно полно реализуют потенциал архитектурной единицы вида. В то же время в описываемой ЦП некоторая часть особей относится к типам 2 и 3, которые характеризуются значительной ролью плагиотропных осей в кроне. Горизонтально растущие ветви, как правило, рассматриваются как одна из адаптаций к дефициту света (Millet et al., 1998; Antonova, Nikolaeva, 2002; Kazakova, Antonova, 2015), однако в исследованной нами ЦП все особи произрастают в условиях полного освещения верхней и средней частей кроны. Вероятно, переход части особей к горизонтальному росту ветвей и частично ствола вполне можно трактовать как механизм ухода от абиотического стресса, вызванного комплексом факторов (Ivanova, Mazurenko, 2013).
Характерной особенностью особей в исследованной ЦП Q. robur , не отмеченной ранее в ЦП иных почвенно-климатических условий, является формирование целого набора осей различного происхождения в нижней части ствола. К ним относятся плагиотропные и полегающие ветви и оси, а также дополнительные более слабые, чем основной, стволы разной формы. Таким образом, в рассматриваемой нами ЦП у особей Q. robur частично реализуется стланиковая форма роста. Полегают только функционально второстепенные оси, а в целом особь развивается по программе «полноценного» дерева. Это свидетельствует о том, что слабозадернованные пески на пологом склоне речной террасы, который в определенной степени защищен от ветров лесным массивом на вершине, более благоприятны для развития Q. robur , чем крутые каменистые склоны Жигулевских гор и возвышенностей Заволжья, в которых описаны именно стланиковые и близкие к стелющимся жизненные формы Q. robur (Belostokov, 1980; Ivanova, Mazurenko, 2013).
В исследованной ЦП преобладают виргинильные и молодые генеративные особи с нормальной жизненностью. Деревья, принадлежащие к указанным онтогенетическим состояниям, произрастают вдоль всего склона на участках с разной степенью задернованности. Какие-либо выраженные зависимости между жизненностью, строением кроны и проективным покрытием травяного яруса мы не обнаружили. В то же время имматурные особи сосредоточены в верхней части склона, вблизи опушки широколиственного леса. В этом локусе значительная часть особей имеет нормальную жизненность. В остальных частях склона имматурные растения встречаются крайне редко. При этом они имеют пониженную и низкую жизненность, а две особи перешли из иммматурного в квазисенильное состояние, или стадию «торчка». Причины отсутствия устойчивого пополнения ЦП новыми особями Q. robur еще предстоит выяснить в ходе дальнейших исследований. На данный момент можно лишь констатировать, что уже после перехода представленных в ЦП особей в виргинильное состояние сообщество не повреждалось сильными низовыми пожарами.
Вероятно, у большинства исследованных особей Q. robur сформирован ксилоподий. Об этом свидетельствуют приземные стелющиеся оси и дополнительные слаборазвитые стволы. Несмотря на наличие одревесневающего многолетнего основания, особи смогли перейти в генеративный период онтогенеза и сформировать жизненную форму дерева. Это свидетельствует об обратимости отклонений от основного пути онтогенеза у данного вида даже в неблагоприятных условиях роста (Smirnova et al., 1984; Ivanova, Mazurenko, 2013). Тем не менее, детальное исследование области корневой шейки и подземных органов – предмет будущих исследований.
В списке индикаторных видов, применяемом в практике лесоводства для определения типов лесорастительных условий (ЛРУ) (Vorobiev, 1953), указывается, что Q. robur произрастает в типах леса В 1–4 – D 1–4 (от сухих суборей до сырых грудов) и не встречается на наименее плодородных почвах, сформированных на песках и на кристаллических породах. Таким образом, полученные нами данные расширяют представления об экологической амплитуде и механизмах биоморфологических адаптаций у Q. robur .
В дальнейшем представляется важным продолжить поиск ЦП Q. robur на выходах песков в лесостепной зоне. Необходимо детально описать организацию ксилоподия и подземных органов. Также важно провести комплексные исследования почв и почвообразующих пород в местообитаниях с ЦП Q. robur .
Заключение
На выходах слабозадернованных песков в долине р. Большой Черемшан Q. robur успешно выходит в генеративный период онтогенеза. Виргинильные и молодые генеративные особи произрастают вдоль всего склона и имеют в основном нормальную жизненность. Локус имматурных особей расположен в верхней части склона вблизи опушки леса. В этих условиях большинство особей имеет нормальную жизненность. В остальных частях склона имматурные растения встречаются редко и обладают только пониженной и низкой жизненностью. В соответствии с направлением роста ствола и самых мощных ветвей от ствола виргинильные и молодые генеративные деревья Q. robur относятся к трем типам. Преобладают особи с ортотропным стволом и восходящими ветвями. Данный тип характерен и для особей Q. robur, произрастающих в сообществах других природных зон и физико-географических районов. В приземной части ствола у большинства особей Q. robur сформирован комплекс, состоящий из ветвей, стелющихся по субстрату осей и/или дополнительных слабых стволов.
Автор благодарит А.В. Иванову (к.б.н., н.с. ИЭВБ РАН – филиала СамНЦ РАН) за логистическое обеспечение исследования. Работа выполнена в рамках в рамках государственного задания Института экологии Волжского бассейна РАН «Комплексная оценка состояния биологических ресурсов и мониторинг природных экосистем Волжского бассейна (FMRW-2025-0047)», № 1024032600230-5-1.6.19.
