Биоморфологические особенности и онтогенез Lathyrus niger в связи с вопросами охраны

Автор: Владыкина Наталья Сергеевна, Казакова Марина Васильевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 22, 2011 года.
Бесплатный доступ
На примере Lathyrus niger (L.) Bernh. описан подход, используемый авторами при изучении редких видов сосудистых растений, занесенных в региональные Красные книги. При анализе охраняемых видов выявляют хорологические, эколого-фитоценотические и популяционно-онтогенетические особенности в естественных местообитаниях и в условиях культуры. Выяснен диапазон жизненных форм L. niger, прослежены особенности прохождения стадий онтогенеза на восточной окраине города Рязани и в условиях Биостанции РГУ, уточнены диагнозы возрастных состояний, определены параметры экологической валентности вида. Узкая экологическая амплитуда по отношению к увлажнению почвы определяется спецификой возобновления вида. Ограничения по солевому режиму почвы и уровню освещенности связаны с низкой конкурентоспособностью растений прегенеративного периода. Светолюбивость L. niger обусловлена довольно поздними сроками цветения. Характерна патиентная жизненная стратегия.
Региональные красные книги, рязанская область, биоморфология, онтогенез, жизненная стратегия, экологическая валентность.
Короткий адрес: https://sciup.org/146116321
IDR: 146116321 | УДК: 581.4+502.7
Biomorphological features and ontogenesis Lathyrus niger in connection with protection questions
On example Lathyrus niger (L.) Bernh. the approach used by authors at studying of rare species of vascular plants, brought in regional Red books is described. At the analysis of protected kinds reveal chronological, ekologo-fitotsenoticheskie and populjatsionno-ontogeneticheskie features in natural habitats and in the conditions of culture. The range of vital forms L. niger is found, features of passage of stages onthogenesis on east suburb of a city of Ryazan and in the conditions of Biological research station RSU are tracked, diagnoses of age conditions are specified, parameters of ecological valency of a kind are defined. The narrow ecological amplitude in relation to soil humidifying is defined by specificity of renewal of a kind. Restrictions on a salt mode of soil and light exposure level are connected with low competitiveness of plants virgil the period. L. niger is heliophyt that is caused by late terms of flowering. Vital strategy is patient.
Текст научной статьи Биоморфологические особенности и онтогенез Lathyrus niger в связи с вопросами охраны
Около 25–30% видов сосудистых растений природной флоры в разных областях Средней России относятся к числу редких или уязвимых, в том числе и в Рязанской обл. [9]. Столь высокий процент редких и уязвимых видов служит основанием для более глубокого анализа этого явления, поскольку на основании составленных списков в регионах разрабатываются программы по охране видов, принимаются административные решения, ведется широкая эколого-просветительская деятельность. До сих пор создание списков редких и охраняемых видов растений относилось к задачам, решаемым в основном флористами. Сведения о присутствии видов, локальности или исключительной редкости их распространения и малочисленности находок во время полевых наблюдений лежат в основе сделанных выводов относительно уязвимости и необходимости принятия мер охраны, а также в основе различных документов, принимаемых на уровне региональной администрации. Обоснованность выводов, как по спискам видов, так и по их характеристикам, повышает комплексный анализ. Актуальна разработка методики оценки статуса вида в определенной части ареала.
Редкие во флоре Рязанской обл. виды сосудистых растений обладают эколого-биологическими особенностями, которые определяют их стенотопность. С этой точки зрения нами проанализированы виды Красной книги Рязанской обл. [15]. Многие из них взяты под охрану и в других, в том числе сопредельных регионах. Исследования проводятся в основном на территории Рязанской обл., как в местах естественного произрастания, так и в условиях культуры на биостанции РГУ им. С.А. Есенина. Для выяснения причин экологической стенотопности проводится сопряженный анализ хорологических, эколого-фитоценотических и популяционно-онтогенетических характеристик, позволяющих понять реакцию видов на антропогенные воздействия.
Заслуживает специального изучения биоморфология Lathyrus niger (L.) Bernh., характеризующийся европейским типом ареала. Восточная граница ареала в Средней России проходит по территории Владимирской, Рязанской, Липецкой, Курской и Белгородской областей. Вид встречается также на Кавказе. Он редок в областях Окского бассейна и занесен в Красные книги Рязанской [15], Липецкой [13], Московской [14], Калужской [12], Тамбовской [16] областей. В соседней Тульской обл. он нередок в засечных лесах [23].
В Восточной Европе вид тяготеет к южной части зоны широколиственных лесов, встречается в лесостепи в бассейне Дона (ассоциации Fruticeto-Quercetum caricosum tanaiticum и Fruticeto-Quercetum vallense donetzicum), на левобережье Днестра в Молдавии (ассоциация Fruticeto-Quercetum vallense moldavicum), на южной окраине Правобережной лесостепи Украины (Spinoso Fruticeto-Quercetum herbosum thyraico-borysthenicum), в южной части Бугско-Днестровского водораздела (ассоциация Quercetum petraeae caricosum moldavicum), в Тульских засеках (ассоциация Tilieto-nemoretum okense), в лесостепи бассейна Дона с Северским Донцом в ассоциации Mixto-nemoretum tanaiticum s.l., в правобережной части Украинского Полесья (ассоциация Carpineto-nemoretum polessicum), в лесостепи между Днестром и Ворсклой (в Carpineto-nemoretum ucrainicum s. l.) [11].
Являясь представителем неморального флористического

комплекса, L. niger входит в эколого-ценотическую группу опушечнолесных растений и в областях средней полосы европейской части России обитает преимущественно в осветленных широколиственных лесах–дубравах паркового типа, по опушкам антропогенно нарушенных дубо-липняков, реже – в сосновых и смешанных лесах, на легких почвах, обычно формирующихся на карбонатной основе. Тяготеет к опушкам, полянам, где наблюдается его обильное цветение [8].
По нашим данным, L. niger является стержне-кистекорневым многолетником, хотя в литературе встречаются разные характеристики жизненной формы: кистекорневой многолетник [1], стержнекорневой каудексовый многолетник [9], короткокорневищный многолетник [21]. Главный стержневой корень сохраняется у растений до средневозрастного генеративного состояния, наряду с ним имеется также несколько хорошо развитых придаточных корней, по своим размерам приближающихся к главному. Разные генеративные особи L. niger могут представлять как стержнекорневую, так и стержне-кистекорневую или кистекорневую [7] жизненную форму. По типу пространственной структуры и степени автономности структурных частей жизненная форма моноцентрическая.
Вид размножается семенами. Партикуляция в генеративном периоде не имеет заметного значения для самоподдержания численности ценопопуляций. В лабораторных условиях семена через несколько месяцев сухого хранения, а также стратифицированные в течение 10–40 суток не прорастали. При стратификации в течение 50–140 суток прорастало 1–5% семян. При весеннем посеве на делянках и при посеве в конце лета нестратифицированных семян всходы появляются только весной следующего года. Всхожесть выше у семян, хранившихся в течение года, чем у свежесобранных. Всходы при самосеве появляются также только весной, наиболее массово – в первой
декаде мая. В последние годы всходы
многочисленные, что связано, видимо, с формирован банка семян и более высокой всхожестью семян, проле или на ее поверхности год или более. Для прора
на биостанции в
РГУ ного
скарификация [20], хотя и скарифицированны

ших в почве необходима ена имеют
растянутый период прорастания.
Семена прорастают ещё осенью, но проросток остается в почве до следующей весны, период подземного роста длится 200–240 дней. Подземное прорастание известно также у Lathyrus vernus (L.) Bernh. [2]. Вероятно, часть семян может прорастать и весной. У проростков (рис. 1, 1–4) сначала появляется главный корень и вслед за ним первичный побег с несколькими (обычно 2–3) редуцированными чешуевидными листьями. В это время на главном корне уже могут быть заметны зачатки боковых корней. Семядоли на поверхность не выносятся. Побег этиолирован, весной он выходит на поверхность и зеленеет. Переход в ювенильное состояние (рис. 1, 5–9) связан с появлением первых настоящих листьев с парой развитых листочков 11–16 мм длиной и 3–4 мм шириной. Листочки следующих листьев до 20 мм длиной, 5–6 мм шириной; стержневой корень до 1 мм в диаметре. В единичных случаях уже у ювенильного растения появляются придаточные корни (рис. 1, 9).
В ювенильном состоянии может начинаться ветвление (через 15– 20 дней или более после появления всходов, в фазе 4–6 листьев). Боковые побеги появляются в подземной или надземной части вичного побега, иногда на уровне почвы. Продолжительность нильного состояния в культуре от 1,5 до 4 месяцев. В естественном тообитании (урочище Карцевский лес, восточная окраина г. Рязани)
июле отмечены ювенильные особи второго года жизни.

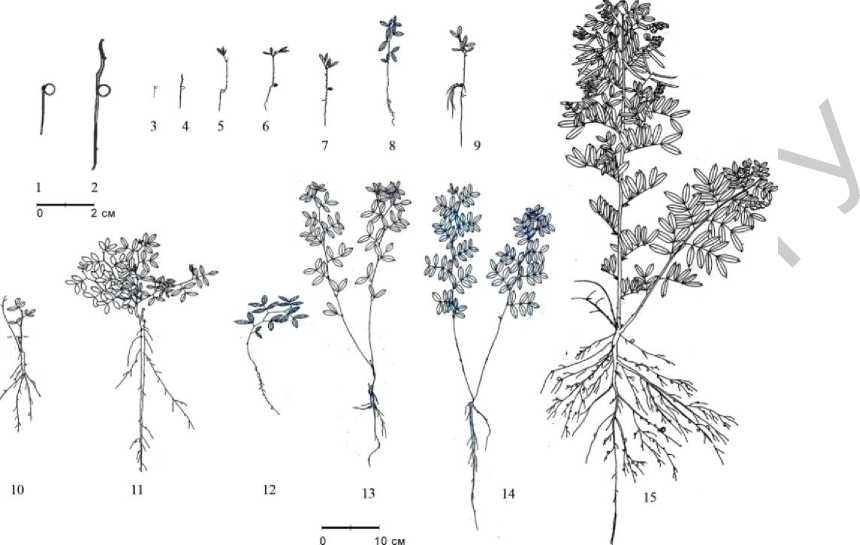
Р и с . 1 . Возрастные состояния Lathyrus niger :
1–4 – проросток; 5–9 – ювенильные растения (последовательные стадии и варианты развития особей); 10–13 – имматурные растения (варианты развития);
14 – виргинильное растение; 15 – молодое генеративное растение;
масштабы для 1–2 и 3–15 различаются;
растения 1–7, 9–11, 15 собраны в культуре, 8, 12–14 – в естественном местообитании
Скорость развития молодых растений, очевидно, зависит от напряженности конкурентных отношений и от уровня освещенности. В условиях культуры на открытых местах растения развиваются быстро. При затенении взрослыми особями испытывают угнетение и к концу вегетационного периода нередко остаются в ювенильном состоянии, не ветвятся, а достигшие имматурного состояния растения заметно менее развиты. Молодые растения угнетаются также при остром дефиците влаги. В естественной ценопопуляции бóльшая часть проростков оказывается в условиях затенения и развивается медленно, к концу первого года жизни остается в ювенильном состоянии.
После развития на побеге 2–5 листьев с 1 парой листочков растения переходят в имматурное возрастное состояние (рис. 1, 10–13). Число листочков сложного листа увеличивается до 2 пар, причем довольно часто сначала появляются листья с непарным третьим листочком. Далее число листочков в составе сложного листа достигает 3 пар. Имматурные растения имеют от 1 до 3 основных побегов. Иногда побеги, развившиеся наиболее рано, уже в середине вегетационного периода первого года жизни отмирают (рис. 1, 10). Может продолжаться ветвление, достигая II порядка, изредка – III порядка. Высота растений от 6 до 30 см. В корневой системе четко выражен главный стержневой корень с хорошо развитыми боковыми корнями. Корень проникает на глубину 20–25 см, возможно и более. Его диаметр 2,5–13 мм (утолщается к концу вегетационного сезона). В условиях культуры главный корень обычно утолщен равномерно, а в естественных ценопопуляциях более утолшенная часть как правило находится в пределах верхнего участка длиной 1,2–3 см. Иногда главный корень на глубине нескольких сантиметров ветвится. У некоторых растений в этот период появляются придаточные корни (обычно один), которые остаются менее мощными, чем главный. В подземной части побегов или на уровне почвы уже в середине вегетационного периода бывают заметны почки возобновления, а иногда и начальные стадии развития побегов возобновления до 1 см длиной (к концу вегетационного периода их количество 1–4). Побеги возобновления остаются этиолированными и в текущем вегетационном сезоне на поверхность не выходят.
В виргинильное возрастное состояние часть растений в условиях культуры переходит в начале вегетационного периода 2 года жизни. Такие растения имеют в средней части побега уже до 4 (редко до 5) пар листочков в составе сложного листа (рис. 1, 14). Чаще развито 2–3, реже 1 побег возобновления. Число порядков ветвления обычно не превышает двух. По количеству остатков отмерших побегов (в среднем 4–5) можно предположить, что возраст виргинильных растений в естественных ценопопуляциях не менее трех лет. В корневой системе обычно выражен стержневой главный корень, который иногда ветвится ближе к основанию, образуя 2–3 боковых корня. У большей части растений также имеется 1, реже 2 придаточных корня, диаметром 2–3 мм. Вероятно, в виргинильном возрастном состоянии начинает формироваться корневище (выражено ещё не у всех растений), длина которого на этом этапе около 1 см. В формировании корневища принимают участие основания надземных побегов и основания придаточных корней. Живые побеги текущего года располагаются близ места отхождения придаточного корня, а на основании главного корня имеются остатки отмерших побегов.

В условиях культуры часть растений уже на 2-м году жизни переходит в генеративное состояние. Молодые генеративные растения 48–56 см высотой (рис. 1, 15), имеют 2–3 генеративных и 1–2 вегетативных побега. Ветвление до II порядка. Могут быть развиты немногочисленные паракладии I порядка, обычно с 1 парциальной кистью, а в главном соцветии – около 7 парциальных кистей. В составе сложного листа в средней части побега 4, реже – 5 пар листочков.
Корневая система представлена стержневым главным корнем, проникающим на глубину 33–35 см и более. Имеются многочисленные боковые корни нескольких порядков, на которых развиты клубеньки. Главный корень покрыт перидермой. Отсутствие у описанных растений придаточных корней и формирующегося корневища связано, вероятно, с меньшим их возрастом по сравнению с растениями из естественных ценопопуляций. У молодого генеративного растения, найденного в природе, имелся придаточный корень и корневище длиной 1,7 см. К концу вегетационного периода в основании побегов хорошо выражены почки и побеги возобновления.
Среднегенеративные растения (рис. 2, 1–2) в условиях культуры имеют в среднем 6–8 (до 22) генеративных побегов, в естественных ценопопуляциях – 1–7. У некоторых растений развивается также по 1–2 вегетативных побега. Побеги возобновления ветвятся на высоте 2–5 см от уровня отхождения от корневища. Высота растений в культуре 95– 140 см, в природе 70–85 (105) см. В условиях культуры, где растения более развиты, обычно имеются паракладии I и II порядков; число парциальных кистей в среднем превышает данный показатель у растений из естественных ценопопуляций. Число листочков в составе сложного листа в средней части побега 5–6 пар, изредка – до 7 пар. В условиях культуры число листочков в среднем больше на 1 пару, чем в естественных ценопопуляциях.
Подземные органы представлены 3–8 утолщенными корнями, среди которых главный корень визуально выделить сложно (рис. 2, 3–4). Сформировано многоглавое укороченное корневище, достигающее в длину 5–9 см. Заметны основания отмерших побегов прошлых лет, до 10 и более. Глубина расположения почек и побегов возобновления (3–8 см) больше у генеративных растений по сравнению с предшествующими возрастными состояниями (так, у имматурных растений – 1–3 см). Более старые растения в условиях культуры весной отрастают заметно позже молодых. В естественной ценопопуляции в Карцевском лесу глубина расположения почек возобновления оказалась в среднем меньше, чем на Биостанции РГУ.
Признаком перехода в стареющее генеративное состояние является преобладание в подземной сфере процессов отмирания над процессами нарастания. Было найдено лишь несколько растений, предположительно отнесенных к субсенильному или сенильному возрастному состоянию. Такие растения не цветут, побеги могут иметь относительно небольшие размеры. Число листочков в составе сложного листа в среднем 4–5 пар, иногда до 6 пар. Число утолщенных корней от 1 до 3, они имеют меньшие размеры по сравнению с генеративными растениями (более тонкие и иногда более короткие), с верхушки иногда происходит отмирание. Кроме того, наблюдаются некротические участки по длине корня, обычно ближе к основанию. Имеются следы отмерших корней. Главный корень, по крайней мере у части растений, отмирает. По-видимому, в ряде случаев происходит партикуляция из-за разрушения участков корневища. У исследованных растений имелись почки возобновления. Найти четкие отличия в надземной сфере сенильных особей от взрослых нецветущих растений пока не удалось.
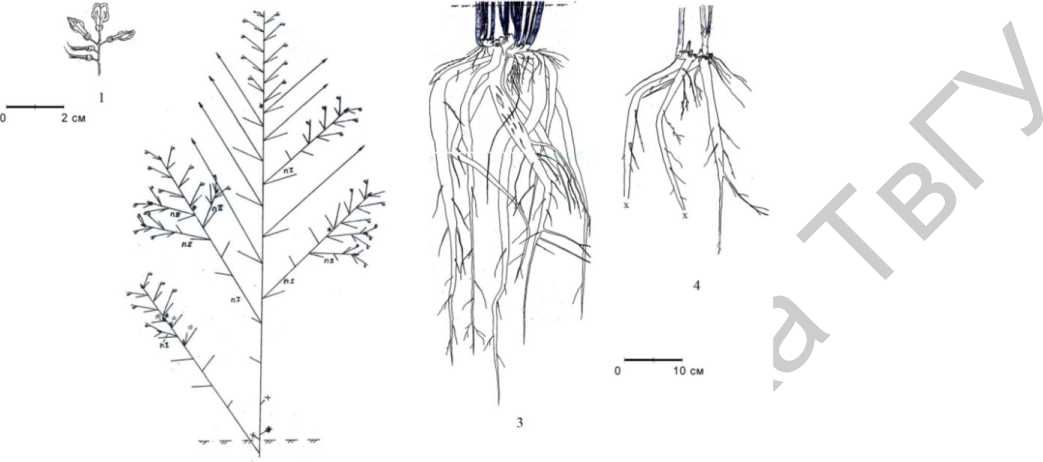
О 20 см
Рис. 2. Средневозрастные генеративные растения Lathyrus niger :
1 – парциальная кисть; 2 – схема побега I порядка;
3 – подземные органы растения с опытной делянки,
4 – подземные органы растения из естественной ценопопуляции;
--
-
- апекс, - лист, - листовой рубец, - парциальная кисть,
* - основное междоузлие, пI, пII - паракладии I и II порядка соответственно
На основании наблюдений 2006–2009 гг. выявлена возрастная структура крупной ценопопуляции L. niger на восточной окраине г. Рязани (численность не менее 1000 особей). Популяция полночленная. В возрастном спектре в 2008 г. преобладали растения прегенеративного возрастного периода (ювенильные – 37%, имматурные – 29%, виргинильные – 10%, генеративные – 17% и взрослые нецветущие – 7%). На пробных площадках не отмечены субсенильные и сенильные растения. Однако вследствие того, что анализ подземных органов всех особей на площадках не проводили, растения этих возрастных состояний могли могли оказаться в составе групп взрослых нецветущих или виргинильных растений.
Расчеты экологической валентности L. niger показали, что вид стеновалентен по двум факторам: увлажнение почв (ЭВ=0,09, от сухолесолугового до влажнолесолугового режима) и солевой режим (ЭВ=0,11, от небогатых до довольно богатых почв). Он эвривалентен по фактору кислотности почвы (ЭВ=0,83). По остальным факторам, для которых имеются данные по шкалам Д.Н. Цыганова [22], вид мезовалентен. Отметим, что условия освещенности ограничены режимами от полуоткрытых пространств до светлых лесов. Индекс толерантности L. niger равен 0,43, т.е. вид относится к гемистенобионтной группе толерантности. А.Д. Булохов и А.И. Соломещ [3] указывают для L. niger мезофитную экобиоморфу.
Можно предположить, что узкая экологическая амплитуда по отношению к увлажнению почвы связана с особенностями возобновления вида. По-видимому, для сохранения побегов возобновления, закладывающихся со второй половины вегетационного периода, необходима достаточная степень увлажнения почвы при хорошей аэрации. Ограничение распространения в естественных условиях по солевому режиму почвы и по освещенности вероятнее всего связано с неспособностью конкурировать в более благоприятных условиях с растениями, обладающими высокой конкурентной способностью. В первую очередь это касается растений прегенеративного возрастного периода. В то же время в условиях культуры на открытых участках L. niger формирует более мощно развитые экземпляры по сравнению с особями естественных ценопопуляций. Это свидетельствует о несовпадении потенциального (аутэкологического) и фактического (синэкологического) оптимумов и соответственно о преобладании жизненной стратегии патиента [17]. О.В. Смирнова [18] также относит L. niger к патиентам. К особенностями биоморфы, определяющим невысокую виолентность L. niger, можно отнести образование небольшого числа корней, невысокие темпы роста. Значительная светолюбивость L. niger связана с довольно поздними сроками ее цветения, приходящимися на период полностью раскрывшихся листьев у широколиственных древесных пород (конец мая–начало июня). Интересно отметить, что широко распространенная в широколиственных и смешанных лесах Средней России L. vernus способна произрастать и в более затененных условиях, поскольку ее цветение приходится на конец апреля – начало мая, то есть на фенофазу «голой кроны» деревьев.
Распространение L. niger в восточном напрвлении может ограничиваться климатическими факторами, в первую очередь континентальностью климата и суровыми зимами. Хотя по шкалам континентальности климата и криоклиматической шкале вид является мезовалентным, его распространение ограничено субматериковым режимом и режимом умеренных зим. Вероятно, такие особенности L. niger связаны с историей формирования вида в Южной Европе. В то же время L. vernus, имеющая евросибирский ареал, по шкале криофильности является эвривалентной и может произрастать в районах с режимом суровых зим, а по шкале континентальности климата хотя и мезовалентна, но приурочена к районам с более континентальным климатом (до промежуточного между континентальным и ультраконтинентальным режимами). На широту распространения вида отрицательное влияние оказало сокращение облесенных площадей в результате хозяйственной деятельности человека. Крупные семена с трудом могут распространяться на значительные расстояния и преодолевать открытые участки, поэтому регулярное внедрение L. niger в изолированные лесные урочища, на наш взгляд, маловероятно.
П.Л. Горчаковский [6] отмечает, что на границах ареалов наблюдается изменение экологическо-фитоценотических особенностей видов. Виды, образующие устойчивые сообщества внутри ареалов, занимают различные местообитания, уже не ассоциируясь друг с другом. Такое явление отмечено и для травянистых растений широколиственных лесов, которые близ границы ареала нередко обитают в иных растительных сообществах. Возможно, у L. niger также происходит расширение фитоценотической амплитуды: ближе к восточной границе ареала она встречается не только в широколиственных лесах, но и в смешанных и сложных сосняках.
Полученные материалы дополнили методику оценки степени и факторов экологической стенотопности редких и уязвимых видов в рамках популяционно-онтогенетического и эколого-методов. Данная методика была нами уже апроби нескольких видов растений [4; 5; 10], занесенных Рязанской и сопредельных областей. Среди них C Lupinaster pentaphyllus Moench s. l., Serratula coronata ценотического а на примере расную книгу a lutetiana L.,
Таким образом, проведенные исследования позволили также уточнить жизненную форму L. niger и установить, что партикуляция не имеет значения для самоподдержания численности ценопопуляции, осуществляется только семенное размножение; выявлены особенности онтоморфогенеза и даны диагнозы возрастных состояний. Растения прегенеративного периода L. niger более уязвимы по сравнению с генеративными. Преобладает жизненная стратегия патиента, Уязвимость особей всех возрастных состояний связана с развитием побегов возобновления и их перезимовкой в верхнем слое почвы. L. niger относится к гемистенобионтной группе толерантности. Основной причиной его экологической стенотопности может быть узкая экологическая амплитуда по факторам увлажнения и освещенности, что необходимо учитывать при разработке рекомендаций по охране вида.

Esenin Ryazan State University, Ryazan