Биоморфология гвоздики борбаша Dianthus borbasii Vandas

Автор: Савиных Наталья Павловна, Шишкина Наталья Ивановна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 2, 2016 года.
Бесплатный доступ
С позиции онтогенетического и биоморфологического подходов изучены и описаны побегообразование, основная жизненная форма, онтогенез и онтоморфогенез охраняемой в Кировской области гвоздики Борбаша Dianthus borbasii Vandas на северо-востоке его распространения. Отмечены морфологическая и динамическая поливариантность растения. Предложены меры по сохранению популяций вида.
Редкие виды, онтогенез, онтоморфогенез, биоморфология, экология, сохранение биоразнообразия
Короткий адрес: https://sciup.org/146116645
IDR: 146116645 | УДК: 581.44
Biomorphology of Dianthus borbasii Vandas
Shoot formation, the primary life form, ontogeny and ontomorphogeny of the protected in the Kirov region Dianthus borbasii Vandas in the north- east of its distribution are studied and described from the standpoint of ontogenetic and biomorphological approaches. Morphological and dynamic polyalternativeness of the plants are determined. Measures to conserve populations of the species are suggested.
Текст научной статьи Биоморфология гвоздики борбаша Dianthus borbasii Vandas
Материал и методика. Мы изучали D. borbasii с 2012 по 2015 г. на особо охраняемой природной территории регионального значения «Медведский бор» в Нолинском р-не Кировской обл. (Шишкина, Рябова, 2015). Основные сообщества здесь - разные типы сосновых лесов на дюнах материкового происхождения боровой террасы р. Вятки (Исупова, 1997), где сосредоточена большая часть охраняемых степных видов -53- региона. D. borbasii произрастает в Медведском бору на песчаной слабо задернованной почве опушек сухих сосновых лесов, вдоль дорог, большей частью - на антропогенно нарушенных территориях.
Исследование проводили под линией электропередач по берегу р. Талицы. Поскольку растение охраняемое, структурную организацию типичных особей изучали весной, летом и осенью без изъятия их из состава сообщества. Собранные и изготовленные образцы растений, фотографии хранятся в гербарной коллекции кафедры биологии и методики обучения биологии Вятского государственного университета. Дополнительно изучены образцы этой коллекции, а также гербария LE. При проведении исследования особое внимание обращали на длину и тип подземных органов, число побегов цветоносных и с неполным циклом развития, особенности цветорасположения, соотношение длины и ширины пластинки листа срединной формации, целостность особей. Особенности растений фиксировали в фотографиях, рисунках, схемах.
Биоморфология растений описана на основе собственных наблюдений с использованием современной терминологии (Современные подходы.., 2008; Актуальные проблемы.., 2012; Фундаментальная и прикладная.., 2014). Жизненную форму характеризовали согласно синтетической множественной классификации (Мейен, 1978; Шорина, 1994). Онтогенетические состояния выделяли в соответствии с дискретным описанием онтогенеза (Работнов, 1950; Уранов, 1975; Жукова, 1995), фазы онтоморфогенеза - по О.В. Смирновой и др. (Ценопопуляции.., 1976).
Результаты и их обсуждение
Побегообразование. Побеговые системы D. borbasii формируются по симподиальной полурозеточной модели (терм.: Серебрякова, 1977). Монокарпический побег (МК) имеет типичное строение для такого рода структур (рис. 1) в понимании И.Г. Серебрякова (1952), W. Troll (1964), И.Г. Борисовой и Г.А. Поповой (1990). Зона возобновления представлена базальным участком розеточного побега с отмершими листьями и пазушными почками возобновления. Ко времени цветения исходного МК из них формируются вегетативные розеточные побеги замещения с 5-7 листьями длиной 3-5 см и шириной 1-2 мм (здесь и далее указаны крайние значения). Это одна из фаз (вегетативного ассимилирующего побега) в развитии МК замещения. Листья розеточного участка, сформировавшиеся в текущем году, 3-7 см длиной и 1,5-2 мм шириной, узколинейные. Структурно и функционально верхний участок розеточного побега представляет среднюю зону торможения. В состав этой же зоны входят вышерасположенные метамеры с длинными междоузлиями и узколинейными листьями, уменьшающимися к верхушке побега до щетиновидных, длиной 1,5-4 см и шириной 1-2 мм. В зоне обогащения и соцветии выражена морфологическая поливариантность. Она проявляется в формировании разных по степени ветвления и развитию цветков структур: из неразвившегося или сформировавшегося цветка с одним вегетативным метамером (рис. 1Б, I-11); с неразвившимся или сформированным терминальным и пазушным цветками и одним вегетативным метамером (рис. 1Б, III-IV); с разной степенью развития боковых и терминального цветков (рис. 1Б, V-VI); с заложенным терминальным цветком и без него и разным числом пазушных бутонов, ни один из которых не реализуется в цветок (рис. 1Б, Vil-VIII). Терминальное соцветие представлено сложным симподием, в составе которого до 9 цветков с несущими их короткими междоузлиями.
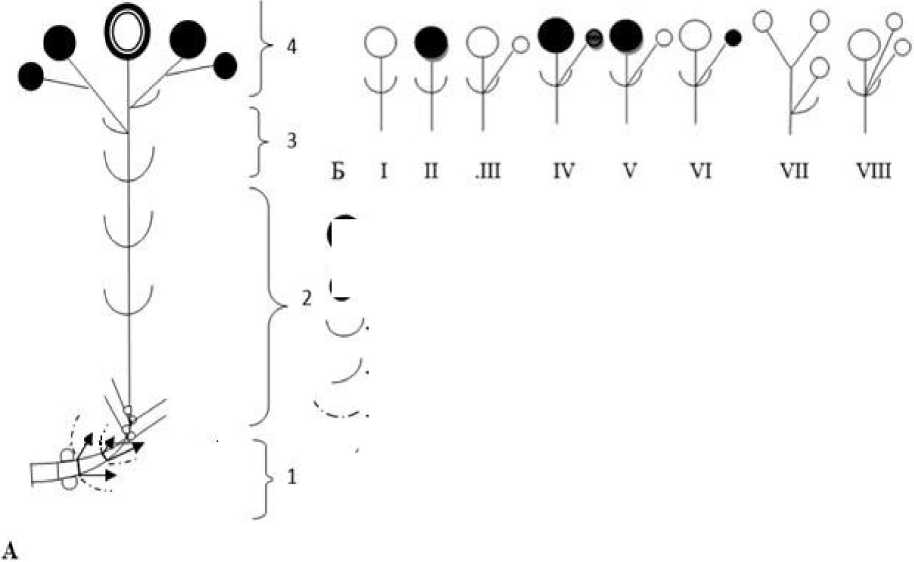
- листы в составе пяракладнев,
- отмершие .листья.
/ - листы текущего года в составе розеточного участка монокарпического побега.
/ - побеги замещения,
D - почки возобновления терминтьное соцветие монокарпического побега, ф - цветки, О - недоразвитые цветки,
- листья удлиненной части побега,
Рис. 1 . Строение монокарпического побега D. borbasii: X. Структурно-функциональные зоны: 1 - возобновления, 2 - средняя торможения, 3 - обогащения, 4 - главное соцветие; Б. Структурные элементы соцветия (I-VIII)
Из-за раннего развития побегов замещения, ко времени цветения МК, на его основе формируется побеговая система двух порядков ветвления: с паракладиями на верхушке и побегами замещения в зоне возобновления. Это - система зрелого моноподиального побега (терм.: Савиных, 1979), построенная по типу дизъюнктивной (терм.: Мамедова, 1985): участки с боковыми побегами (побегами замещения и паракладиями) разделены средней зоной торможения. Из таких систем сложено побеговое тело растения. Целостными они существуют до отмирания МК с верхушки до зоны возобновления осенью. Побеги ' -55- замещения на первых этапах развития, с одной стороны, обеспечивают успешную репродукцию исходным МК и самоподдержание ценопопуляции, с другой - возобновление (самоподдержание) особи. Такое развитие побегов свойственно и другим степным растениям (Савиных, 1997), что обеспечивает им успешную семенную репродукцию до наступления засухи.
МК D. borbasii развиваются с прохождением основных фаз по И.Г. Серебрякову (1959) и Т.И. Серебряковой (1971). В фазе почки побег находится один год. В фазе вегетативного ассимилирующего побега - с июня текущего года до начала июня следующего. Фаза бутанизации, цветения и плодоношения продолжается с июня по август. В фазе вторичной деятельности побег существует после отмирания удлиненной части с верхушки до зоны возобновления. Образующийся резид сохраняется до конца жизни особи. МК без учета фазы вторичной деятельности обычно дициклические. Как у большинства степных трав с симподиальными моделями побегообразования, они развиваются с зимним перерывом.
Онтоморфогенез D. borbasii
Латентный период. Плод — лизикарпная многосемянная вскрывающаяся коробочка. Семена мелкие, сплюснутые с обеих сторон, вогнутые, округлой формы, закругленные на узком конце. Поверхность семенной кожуры ребристая. Семена в раскрытых коробочках черные.
Предгенеративный период.
Проростки. Прорастание надземное. При высокой освещенности и отсутствии травяного покрова прорастание семян, по нашим данным, почти 100%. Семядольные листья длиной 0,2-0,3 см, шириной 0,1-0,12 см рано высыхают и отмирают. Растение стержнекорневое, моноподиально-розеточное, однопобеговое. Фаза онтоморфогенеза -первичный вегетативный розеточный побег.
Ювенильные растения высотой от 0,8 до 1,5 см. Здесь и далее высоту растений определяли по размерам самого длинного вертикально расположенного листа. Вслед за высохшими бурыми семядолями располагаются 3-4 пары супротивно расположенных первых настоящих продолговато-ланцетных листьев длиной 0,2-3 см и шириной 0,1-0,2 см. Листовые пластинки цельнокрайние, с острой верхушкой отклонены от оси стебля и почти параллельны поверхности. Гипокотиль короткий. Корневая система стержневая. Главный корень ветвится и уходит вглубь почвы на 5-6,2 см. Особи, по-прежнему, стержнекорневые моноподиально-розеточные однопобеговые растения в фазе первичного розеточного побега. Критерии этого онтогенетического состояния: наличие главного побега, отсутствие живых семядолей, цельнокрайние листья.
У имматурных растений в числе от 2 до 5-6 ортотропных побегов высотой до 4 см, с 3-4 парами продолговато-ланцетных листьев. Они серовато-зеленые длиной 1,5-7 см и шириной 0,08-0,12 см, листовые пластинки по краям с небольшими зубчиками. Главный корень от 6,5 до 7 см длиной, ветвится до 111 порядка. Особи стержнекорневые моноподиально-розеточные ветвящиеся. Фаза онтоморфогенеза -первичный куст. Критерий выделения этого онтогенетического состояния: ветвление главного побега.
Признаки виргинилъных растений в основном те же, что у имматурных. Это стержнекорневые моноподиально-розеточные ветвящиеся растения. Изменяется длина (от 6,5 до 7,0 см) и ширина (от 0,1 до 0,2 см) листа. Край листовой пластинки зубчатый. Главный корень длиной до 15 см, ветвится до 111 порядка. Фаза онтоморфогенеза - по-прежнему первичный куст. Поэтому основной критерий этого онтогенетического состояния - размеры листовой пластинки. Прегенеративный период не длителен, продолжается в течение двух, обычно трёх лет.
Генеративный период.
Раннегенеративные растения в зависимости от субстрата отличаются длиной междоузлий зоны возобновления МК, что определяет габитус первичного куста. На подвижном субстрате (при засыпании песком) междоузлия длиннее, из-за чего куст имеет вид «рыхлого». На неподвижном (плотном) субстрате междоузлия короче, куст компактный («плотный»), растение подобно «подушке».
Первичный куст состоит из побегов: 1) ортотропных дициклических полурозеточных МК высотой от 15 до 60 см; 2) ди- и моноциклических вегетативных полурозеточных с неполным циклом развития; 3) вегетативных розеточных побегов замещения в фазе вегетативного ассимилирующего побега. На главном корне длиной 7-10 см в верхнем слое почвы формируются тонкие многочисленные эфемерные придаточные (терм. Шалыт, I960) корни, отмирающие в конце вегетационного периода, и один - два боковых. Поэтому признаки для выделения раннегенеративных особей: «плотный» или «рыхлый» первичный куст с полурозеточными МК, небольшим числом вегетативных полурозеточных побегов и побегов с неполным циклом развития, с двумя типами корней.
Среднегенеративные растения существуют в виде рыхлых кустов больших диаметров. У этих растений наиболее выражена морфологическая поливариантность, описанная ранее. МК высотой от 16 до 60 см у особи от 3 до 49. На одном растении может быть от 19 до 182 цветков и бутонов и от 1 до 182 плодов. Вегетативных розеточных побегов в составе куста от 3 до 15-17. Побегов с неполным циклом развития от одного до 5-7. Боковые оси формируются в ходе базисимподиального нарастания. Главный корень длиной более 20 см. Растения на выровненной поверхности мало ветвящиеся, с эфемерными корнями в верхнем слое грунта. На склонах с подвижным субстратом главный корень имеет несколько боковых. Эти особи - стержнекорневые растения с боковыми симподиальными системами из резидов полурозеточных олиго-/дициклических МК. Фаза онтоморфогенеза -рыхлый куст. Критерии выделения этого онтогенетического состояния у D. borbasii: крупные размеры растения, поливариантность и множественность МК, небольшое число побегов с неполным циклом развития. В этом онтогенетическом состоянии часто заканчивается жизненный цикл растения, особенно на открытых местах.
Позднегенеративные особи - стержнекорневые растения с хорошо различимыми симподиальными боковыми осями. МК высотой 13-30 см в числе от 1 до 27. На МК имеется одновременно от одного до 15(16) цветков и от одного до 30 (35) бутонов, из которых цветки в будущем не развиваются, плодов, соответственно до 15. Вегетативных полурозеточных побегов с неполным циклом развития может быть от одного до 12. Вегетативных розеточных побегов от 3 до 16-17. Главный корень становится более тонким, наружные ткани его легко отслаиваются. В составе корневой системы боковые многолетние и немногочисленные эфемерные придаточные корни. Эти особи -стержнекорневые растения с боковыми симподиальными системами из резидов МК и побегов с неполным циклом развития. Фаза онтоморфогенеза - рыхлый куст. Критерии выделения позднегенеративных особей: много побегов с неполным циклом развития и МК с неразвившимися цветками. Эти особи отмирают.
Растения субсенильного и сенильного онтогенетических состояний не отмечены.
Таким образом, на северо-востоке ареала D. borbasii длительность жизни особей определяется условиями освещенности, а A-тип онтогенеза сочетает признаки обоих подтипов (Ai и А2). Поддержание ценопопуляции семенное, особи - за счёт формирования дициклических МК. Сохранению популяций вида в этой части ареала способствовало наличие антропогенно нарушенных территорий и высокий уровень освещенности в отдельных типах (остепненных) сосняков. Для поддержания устойчивых ценопопуляций этого вида при естественной трансформации сосновых лесов необходимо создание путем регулируемой хозяйственной деятельности ключевых участков сосновых лесов с повышенным уровнем освещенности.
Заключение. D. borbasii - поликарпическое моноцентрическое многолетнее стержнекорневое травянистое растение из олиго- и дициклических монокарпических полурозеточных побегов с
Вестник ТвГУ. Серия "Биология и экология". 2016. № 2 разнообразными вариантами цветорасположения; высоко конкурентный вид на открытых местообитаниях.
Онтогенез D. borbasii незаконченный, обрывающийся без постгенеративного периода и 8 онтогенетических состояний. Сохранение этого вида на северо-востоке европейской России возможно при осветлении сосняков.
Фундаментальная и прикладная биоморфология в ботанических и экологических исследованиях: материалы Всероссийской научной конференции с международным участием (к 50-летию Кировского отделения русского ботанического общества). Киров: Радуга - ПРЕСС. 358 с.
Ценопопуляции растений (основные понятия и структура). 1976. М.: Наука. С. 14-44. ' '
Шалыт М. 1960. Эфемерные корни и их значение / Труды Первой научной конф, препод, биол., с.-х. и хим. дисциплин педагог, ин-тов УССР. Симферополь. С. 49-94.
Шишкина Н.И., Рябова Е.В. 2015. Онтогенез гвоздики Борбаша (Dianthus borbasii Vandas.) / Принципы и способы сохранения биоразнообразия: материалы VI Всероссийской научной конференции с международным участием / ред. Г.О. Османова; Л.А. Жукова Йошкар-Ола: Изд-во МарГУ. С. 133-135. '
Шорина Н.И. 1994. Экологическая морфология и популяционная биология представителей подкласса Polypodiophyta: автореф. дне.... докт. биол. наук М.: МГПИ. 34 с. - ‘ -
Troll W. 1964. Die Infloreszenzen. Bd. 1. Jena. 615 8.
BIOMORPHOLOGY OF DIANTHUS BORBASII VANDAS
N. P. Savinykh, N. I. Shishkina
Vyatka State University, Institute of Biology and Biotechnology, Kirov
Shoot formation, the primary life form, ontogeny and ontomorphogeny of the protected in the Kirov region Dianthus borbasii Vandas in the north- east of its distribution are studied and described from the standpoint of ontogenetic and biomorphological approaches. Morphological and dynamic polyalternativeness of the plants are determined. Measures to conserve populations of the species are suggested.
Савиных Н.П. Биоморфология гвоздики Борбаша Dianthus borbasii Vandas / Н.П. Савиных, Н.П. Шишкина // Вести. ТвГУ. Сер.: Биология и экология. 2016. № 2. С. 53-61.
Список литературы Биоморфология гвоздики борбаша Dianthus borbasii Vandas
- Актуальные проблемы современной биоморфологии. 2012/ред. Н.П. Савиных Киров: Радуга -ПРЕСС. 610 с.
- Борисова И.Г., Попова Г.А. 1990. Разнообразие функционально-зональной структуры побегов многолетних трав//Бот. журнал. Т. 75. № 10. С. 1420-1426.
- Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. 2003. Иллюстрированный определитель растений Средней России: Покрытосеменные (двудольные: раздельнолепестные). М.: Т-во научных изданий КМК. Ин-т технологических исследований. Т. 2. 665 с.
- Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола: РИИК «Ланар». 224 с.
- Исупова Е.М. 1997. Рельеф//Энциклопедия земли Вятской. Т.7. Природа. Киров: Вятка. С. 112-137.
- Красная книга Кировской области: Животные, растения, грибы. 2014. Изд. 2-е/ред. О.Г. Баранова, Е.П. Лачоха, В.М. Рябова, В.Н. Сотников, Е.М. Тарасова, Л.Г. Целищева. Киров: «Кировская областная типография». 336 с.
- Мамедова Э.Т. 1985. Структура цветоносных побегов в сем. Геснериевых (Gesneriaceae): автореф. дис. …канд. биол. наук. М.:16 с.
- Мейен С.В. 1978. Основные аспекты типологии организмов//Журн. общ. биол. Т. 39. № 4. С. 495-508.
- Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах//Тр. БИН АН СССР. Сер. 3. Геоботаника. Вып. 6. М.: Наука. С. 77-204.
- Савиных Н.П. 1979. Побегообразование и взаимоотношения жизненных форм в секции Veronica рода Veronica L.//Бюл. МОИП. Отд. биол. Т. 84. Вып. 3. С. 92-105.
- Савиных Н.П. 1997. Вероники секции Veronica//Биологическая флора Московской области. М.: Изд-во «Гриф и К0». Вып. 14. С. 160-180.
- Савиных Н.П. 2013. Сохранение биоразнообразия с позиций биоморфологии//Вестн. ТвГУ. Сер. Биология и экология. Вып. 31. С. 155-165.
- Серебряков И.Г. 1952. Морфология вегетативных органов высших растений. М.: Сов. наука. 391 с.
- Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования//Учен. зап. МГПИ им. Потемкина. Т. 100. Вып. 5. С. 3-37.
- Серебрякова Т.И. 1971. Морфология растений и эволюция жизненных форм злаков. М.: Наука. 360 с.
- Серебрякова Т.И. 1977. Об основных «архитектурных моделях» травянистых многолетников и модусах их преобразований//Бюл. МОИП. Отд. биол. Т. 82. Вып. 2. С. 112-128.
- Современные подходы к описанию структуры растений 2008/ред. Н.П. Савиных, Ю.А. Бобров. Киров: ООО «Лобань». 355 с.
- Уранов А.А. 1975. Возрастной спектр фитоценопопуляции как функции времени и энергетических волновых процессов//Науч. докл. высш. шк. Биол. науки. № 2. С. 7-34.
- Фундаментальная и прикладная биоморфология в ботанических и экологических исследованиях: материалы Всероссийской научной конференции с международным участием (к 50-летию Кировского отделения русского ботанического общества). Киров: Радуга -ПРЕСС. 358 с.
- Ценопопуляции растений (основные понятия и структура). 1976. М.: Наука. С. 14-44.
- Шалыт М. 1960. Эфемерные корни и их значение/Труды Первой научной конф. препод. биол., с.-х. и хим. дисциплин педагог. ин-тов УССР. Симферополь. С. 49-94.
- Шишкина Н.И., Рябова Е.В. 2015. Онтогенез гвоздики Борбаша (Dianthus borbasii Vandas.)/Принципы и способы сохранения биоразнообразия: материалы VI Всероссийской научной конференции с международным участием/ред. Г.О. Османова; Л.А. Жукова Йошкар-Ола: Изд-во МарГУ. С. 133-135.
- Шорина Н.И. 1994. Экологическая морфология и популяционная биология представителей подкласса Polypodiophyta: автореф. дис…. докт. биол. наук М.: МГПИ. 34 с.
- Troll W. 1964. Die Infloreszenzen. Bd. 1. Jena. 615 S.
