Биопродукционные характеристики живого напочвенного покрова старовозрастных пихто-ельников северо-востока Костромской области

Автор: Грозовская Ирина Сергеевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология растительных сообществ
Статья в выпуске: 1-6 т.14, 2012 года.
Бесплатный доступ
Приведены результаты измерения биомассы живого напочвенного покрова трех типов пихто-ельников востока Костромской области. В живом напочвенном покрове пихто-ельников выделены микрогруппировки по доминирующей эколого-ценотической группе (ЭЦГ) видов. Непрямая ординация показала сходство однотипных микрогруппировок по надземной биомассе в трех типах фитоценозов. Наибольшую биомассу имеют крупнопапоротниковые микрогруппировки, наименьшую – бореально-мелкотравные. Высокие значения биомассы имеет черничная микрогруппировка. По результатам расчетов биомассы живого напочвенного покрова на учетных площадях максимальную надземную биомассу имеет ельник черничный, максимальную подземную – пихто-ельник бореально-мелкотравно-чернично-крупнопапоротниковый.
Южная тайга, разнообразие растительности, биомасса живого напочвенного покрова, микрогруппировка, эколого-ценотическая группа
Короткий адрес: https://sciup.org/148200845
IDR: 148200845 | УДК: 634.0.581.526.42
Biomass parameters of ground vegetation in old-growth spruce-fir forest in the north-east of the Kostroma region
Biomass of ground vegetation was measured in three forest types with domination of spruce and/or fir in the overstorey. According to the plant functional group domination, there were identified several microgroups in the ground vegetation for the each forest type. DCA analysis showed a similarity of the above-ground biomass of the same microgroups in different forest types. The large-fern microgroups have the maximal biomass values and the boreal small herb microgroups have the minimal ones. Vaccinium myrtillus microgroup has quite high values of the biomass. The maximal total above-ground biomass of ground vegetation was in the Spruce forest with domination of Vaccinium myrtillus. The maximal total below ground biomass was in Spruce-fir forest with co-domination of boreal small herbs, Vaccinium myrtillus and large ferns.
Текст научной статьи Биопродукционные характеристики живого напочвенного покрова старовозрастных пихто-ельников северо-востока Костромской области
В современном мире остро встает проблема сохранения биологического разнообразия лесных экосистем при одновременно эффективном использовании лесных ресурсов. Оптимальные способы ведения лесного хозяйства должны сохранить не только продуктивную способность лесной экосистемы, но и ее биологическое разнообразие [4]. Для оптимизации хозяйства при сохранении экологических функций лесного фитоценоза необходимо знать продуктивность всех его элементов. В том числе важен учет продуктивности живого напочвенного покрова (травяно-кустарничкового и мохово-лишайникового ярусов), поскольку его органическое вещество вносит большой вклад в общий круговорот углерода и азота в лесной экосистеме [6].
В качестве объектов исследования были выбраны лесные экосистемы южной тайги Европейской части России. Цель работы заключалась в проведении сравнительной оценки разнообразия и биомассы живого напочвенного покрова ельников и пих-то-ельников северо-востока Костромской области. В качестве полевых объектов исследования нами были выбраны пихто-ельники Костромской области, находящиеся в подзоне южной тайги. Исследования проводили в июле 2011 года в Октябрьском районе Костромской области (Октябрьское лесничество). По лесорастительному районированию А.В. Письмерова [5] исследованные участки относят к району пихтово-еловых лесов южных отрогов Северных Увалов. Характерной особенностью таких лесов является примесь в еловых древостоях Abies sibirica и Tilia cordata [5]. Отличительной особенностью исследуемых пихто-ельников является также практически равное участие в напочвенном покрове видов бореальной и неморальной эколого-ценотических групп (ЭЦГ) [9].
Сбор материала проведен в 3-х типах лесных сообществ, расположенных в пределах Веденьёв-ского участкового лесничества. Пихто-ельник бо-реально-мелкотравно-чернично-крупнопапоротниковый (ПЕБ) (45 кв.) описан на водоразделе, на относительно выровненном участке поверхности. Пихто-ельник разнотравный (ПЕР) (49 кв.) расположен на правом высоком берегу реки Ветлуги на 1-ой надпойменной террасе (в пределах водоохранной зоны). Ельник черничный (ЕЧ) (45 кв.) описан на водораздельном участке с небольшим уклоном.
В каждом из фитоценозов выполнены геоботанические описания растительности. На площадках 10 м × 10 м описаны проективное покрытие древесного, кустарникового, травяно-кустарничкового и мохово-лишайникового ярусов; для всех видов по ярусам зафиксированы покрытие-обилие по шкале Браун-Бланке. Виды мохообразных определены до рода.
Для сбора материала по биомассе напочвенного покрова в каждом из изучаемых сообществ заложены учетные площади размером 40 м × 40 м. Сбор данных по биомассе напочвенного покрова производили с учетом неоднородного распределения видов сосудистых растений в сообществе. В пределах учетных площадей были выделены микрогруппировки по доминирующей в живом напочвенном покрове ЭЦГ. Выделены микрогруппировки: черничная, крупнопапоротниковая, бореально-мелкотравная, неморальная, бореально-неморальная, высокотравная. В ПЕБ и ЕЧ выделено по 3 микрогруппировки, в ПЕР – 5 микрогруппировок. В пределах микрогруппировок с помощью рамки закладывали площадки 25 см × 25 см. Учет биомассы напочвенного покрова производили методом монолитов [7]. Для каждой из заложенных площадок составлен список видов сосудистых растений и мохообразных с учетом их обилия. Растения срезали на уровне почвы и разделяли по видам. Мхи сни- мали с поверхности почвы без разделения на виды. Выкапывали почвенный монолит со сторонами 25 см и разбирали подземные части растений с разделением их по видовой принадлежности. Растения были высушены до воздушно-сухого состояния, затем в лабораторных условиях образцы дополнительно высушили в термостате до сухого состояния и взвешены. Всего было заложено 60 пробных площадок: по 20 площадок в каждом типе сообщества. На всех 20-ти отбирали образцы по надземной биомассе, а на 10-ти из них дополнительно брали образцы по подземной биомассе. В результате собраны данные по биомассе 30-ти видов сосудистых растений напочвенного покрова в исследуемых типах леса. Дополнительно, на 17-ти пробных площадках были собраны данные по общей биомассе мохообразных без деления их на виды.
Средние значения надземной и подземной биомассы микрогруппировок живого напочвенного покрова пихто-ельников представлены в Таблице 1. Наименьшая биомасса растений отмечена в бореально-мелкотравных микрогруппировках. Во всех исследуемых фитоценозах они имеют сходные значения надземной биомассы. Подземная биомасса бореально-мелкотравных микрогруппировок значительно выше в ельнике черничном, чем в других сообществах. Такой результат объясняется тем, что при подсчете подземной биомассы микрогруппировки была учтена биомасса корневищ черники, присутствующих на площадке. Неморальная мик- рогруппировка в ПЕР отличается более высокими продукционными показателями. Среди черничных микрогруппировок, которые были выделены в напочвенном покрове 2-х типов сообществ, наибольшую надземную биомассу имеет микрогруппировка в ельнике черничном, а наибольшую подземную – ПЕБ. Высоким варьированием отличаются показатели биомассы крупнопапоротниковых микрогруппировок. Самые большое значение надземной биомассы имеют микрогруппировки в ПЕР. Наиболее вариабельной оказалась подземная биомасса крупнопапоротниковых микрогруппировок. В ПЕР она почти в два раза превышает подземную биомассу этих группировок в ПЕБ.
Рассчитана биомасса живого напочвенного покрова на учетных площадях (40×40 м2) с учетом доли, занимаемой микрогруппировками, от его общего проективного покрытия в трех типах фитоценозов (Рис. 1). В ПЕБ наибольшую долю составляют крупнопапоротниковая (40 %) и черничная микрогруппировки (35 %), наименьшую – бореальномелкотравная (25 %). Для ПЕР характерны высокое участие в формировании напочвенного покрова бореально-мелкотравной (35 %) и неморальной (30 %) микрогруппировок и невысокий процент участия бореально-неморальной (15 %), высокотравной (10 %) и крупнопапоротниковой (10%) микрогруппировок. В ЕЧ 90 % занимает черничная микрогруппировка, 9 % – бореально-мелкотравная, 1% – крупнопапоротниковая.
Таблица 1 Биомасса микрогруппировок живого напочвенного покрова трех исследованных типов сообществ (сухой вес, г/м2)
|
Микрогруппи-ровка |
Биомасса |
Тип сообщества |
||
|
ПЕБ |
ПЕР |
ЕЧ |
||
|
Черничная |
Надземная |
189,4±85,7 |
— |
252,1±126,9 |
|
Подземная |
435,8±244,5 |
— |
256,4±69,7 |
|
|
Крупнопапоротниковая |
Надземная |
222,1±129,8 |
308,7±135,8 |
154,9±16,5 |
|
Подземная |
1253,2±634,5 |
2268,4±1445,1 |
— |
|
|
Бореальномелкотравная |
Надземная |
31,5±22,9 |
32,2±3,9 |
34,1±13,0 |
|
Подземная |
77,3±20,6 |
46,5±10,0 |
218,0±47,5 |
|
|
Неморальная |
Надземная |
— |
70,5±11,3 |
— |
|
Подземная |
— |
112,3±18,2 |
— |
|
|
Бореальнонеморальная |
Надземная |
— |
56,5±28,5 |
— |
|
Подземная |
— |
— |
— |
|
|
Высокотравная |
Надземная |
— |
145,8±20,8 |
— |
|
Подземная |
— |
— |
— |
|
Максимальную подземную биомассу живого напочвенного покрова имеет ПЕБ, что связано с высокой долей участия крупнопапоротниковой и черничной микрогруппировок. В этих микрогруппировках доминируют и по обилию и по биомассе Dryopteris expansa и Vaccinium myrtillus, для которых характерно наличие одревесневающих корневищ. Максимальные значения по надземной биомассе имеет ЕЧ, что также объясняется доминированием в напочвенном покрове черничной микрогруппировки и наличием у Vaccinium myrtillus над- земных одревесневающих побегов. Самую низкую надземную биомассу имеет ПЕР. В этом фитоценозе в образовании напочвенного покрова преимущественно принимали участие травяные микрогруппировки.
Проведен сравнительный анализ значений надземной биомассы однотипных микрогруппировок в исследуемых типах сообществ методом непрямой ординации (DCA, Detrended Correspondense Analysis). Всего проанализировано 60 навесок надземной биомассы. Расчеты производили в системе PC-ORD for Windows версии 5.
Ординация навесок надземной биомассы показала, что черничная и крупнопапоротниковая микрогруппировки занимают крайние позиции в орди-национном пространстве и образуют обособленные группы. Третья обособленная группа образована неморальной и высокотравной группировками. Бореально-неморальная и бореально-мелкотравная микрогруппировки занимают центральное положение (Рис. 2). Таким образом, результаты ординации подтверждают наличие неоднородности напочвенного покрова (мозаичности), которая отражается не только в видовом составе микрогруппировок, но и в их продукционных характеристиках. Одинаковые микрогруппировки в разных типах сообществ имеют сходные значения по надземной биомассе, наибольшее варьирование значений биомассы отмечено для бореально-мелкотравной микрогруппировки. Полученные результаты хорошо согласуются с данными, полученными О.В. Смирновой с коллегами при изучении продуктивности напочвенного покрова старовозрастных пихто-ельников с кедром высокотравных [1—3, 8—9]. Дальнейшее исследование функциональной значимости биомассы растений живого напочвенного покрова может лечь в основу выделения функциональных типов растений (ФТР) по биомассе.
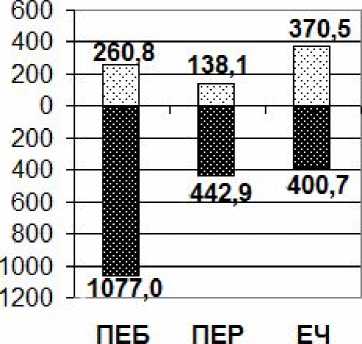
Типы сообществ а Надземная биомасса и Подземная биомасса
Рис. 1. Надземная и подземная биомасса живого напочвенного покрова на учетных площадях трех исследованных типов сообществ (сухой вес, кг/1600 м2).
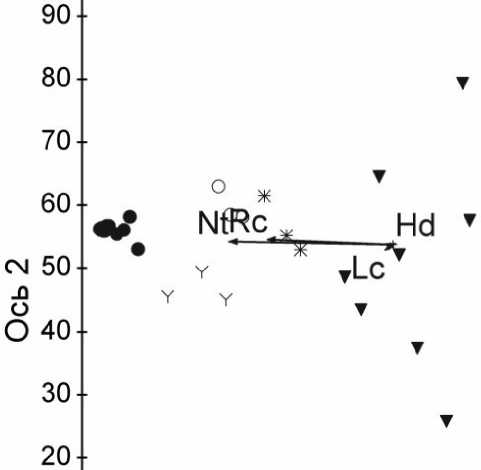
Микрогруппировки
-
■ Черничная
-
• Крупнопапоротниковая
-
▼ Бореально-мелкотравная
-
y Неморальная
* Бореально-неморальная о Высокотравная

ч------------------1------------------1------------------1------------------1------------------►—
О 20 40 60 80 100
Ось 1
Рис. 2. Ординация навесок по надземной биомассе живого напочвенного покрова в трех типах исследованных растительных сообществ (DCA).
Обозначения векторов экологических шкал Д.Н. Цыганова (1983): Hd – шкала увлажнения почв, Nt – шкала богатства почв азотом, Rc – шкала кислотности почв, Lc – шкала освещенности-затенения
Список литературы Биопродукционные характеристики живого напочвенного покрова старовозрастных пихто-ельников северо-востока Костромской области
- Восточноевропейские леса: история в голоцене и современность/О.В. Смирнова (ред.). М.: Наука, 2004. Кн. 1. 479 с.
- Запрудина М.В. Фитомасса травяно-кустарничкового и мохового ярусов темнохвойных лесов Печоро-Илычского заповедника//Изв. Самарского НЦ РАН. 2010. Т. 12, № 1 (3). С. 876-879.
- Запрудина М.В., Смирнов В.Э. Микромозаичная организация травяно-кустарничкового и мохового покрова высокотравных пихто-ельников с кедром нижней части бассейна реки Большая Порожняя (приток р. Печора)//Тр. Печоро-Илычского заповедника. Сыктывкар: Изд-во Коми НЦ УрО РАН, 2010. Вып. 16. С. 60-68.
- Основы устойчивого лесоуправления: учеб. пособие для вузов/М.Л. Карпачевский, В.К. Тепляков, Т.О. Яницкая, А.Ю. Ярошенко; Всемирный фонд дикой природы (WWF). М., 2009. 143 c.
- Письмеров А.В. Методические указания по лесорастительному районированию Костромской области (с применением математических методов). М.: ВНИИЛМ, 1977.
- Ремезов Н.П., Быкова Л.Н., Смирнова К.Н. Потребление и круговорот азота и зольных элементов в лесах Европейской части СССР. М.: Изд-во МГУ, 1959. 284 с.
- Родин Л.Е., Ремезов Н.П., Базилевич Н.И. Методические указания к изучению динамики биологического круговорота в фитоценозах. Л.: Наука, 1968. 145 с.
- Смирнова О.В., Алейников А.А., Семиколенных А.А., Бовкунов А.Д., Запрудина М.В., Смирнов Н.С. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике//Лесоведение. 2011. № 6. С. 67-78.
- Смирнова О.В., Бобровский М.В., Ханина Л.Г., Смирнов В.Э. Биоразнообразие и сукцессионный статус старовозрастных темнохвойных лесов Европейской России//Успехи современной биологии. 2006. Т. 126, № 1. С. 27-49.
