Биоразнообразие верхнеордовикского рифа Большая Косью (Северный Урал)
")
Автор: Шмелва Л.А.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 11 (287), 2018 года.
Бесплатный доступ
В строении рифа Большая Косью выделены 4 интервала, соответствующие фазам/стадиям развития. Самое высокое биоразнообразие рифовой палеоэкосистемы характерно для III интервала, тогда как для завершающей развитие рифа IV стадии - наиболее низкое. Среди каркасостроителей преобладают строматопороидеи, табуляты и сфинктозоа. Широкое распространение и таксономическое разнообразие сфинктозоа впервые установлено в верхнеордовикских рифах Урала. Это были преимущественно криптобионты, они жили в средах, благоприятных для других метазоа, и не имели, таким образом, собственной экологической ниши. Однако при ухудшении условий обитания они довольно быстро адаптировались к изменяющимся условиям среды и стали одной из доминантных групп среди каркасостроящих организмов.
Биоразнообразие, риф, верхний ордовик, р. илыч, северный урал
Короткий адрес: https://sciup.org/149129287
IDR: 149129287 | УДК: 551.733.13 | DOI: 10.19110/2221-1381-2018-11-38-42
Biodiversity of the upper Ordovician Bol'shaya Kos'yu reef (Northern Urals)
In the structure of the Bol'shaya Kos'yu reef, four intervals corresponding to the phases/ stages of growth are established. The highest biodiversity of the reef ecosystem is typical for interval III, while for the final stage IV - it is the lowest. The frame-building organisms are dominated by stromatoporoids, tabulates and sphinctozoan. For the first time, a wide distribution and taxonomic diversity of sphinctozoan in the Upper Ordovician reefs of the Urals is recognized. They were mostly cryptobionts, lived in environments conducive to other Metazoa, and had, thus, their own ecological niche. However, with the deterioration of living conditions, they quickly adapted to changing environmental conditions and became one of the dominant groups among frame-building organisms.
Текст научной статьи Биоразнообразие верхнеордовикского рифа Большая Косью (Северный Урал)
Рифы как важнейшие источники информации о былых состояниях биосферы являются уникальными объектами исследования. На западном склоне севера Урала в настоящее время известно три верхнеордовикских рифа (Лек-Елец, Бадъя и Большая Косью). Лишь один из них — риф Большая Косью на Северном Урале — практически не затронут процессами доломитизации, что делает его уникальным геологическим объектом палеоэкологического изучения с позиций развития рифовой экосистемы в палеозое Урала. Первое упоминание о биогермных известняках верхнего ордовика в бассейне р. Илыч (рис. 1) принадлежит А. Г. Кондиайн [4]. Позднее разрез изучался А. И. Антошкиной, впервые проследившей в структуре рифа смену стадий развития палеосообществ [1]. В настоящее время выявлен сложный па-
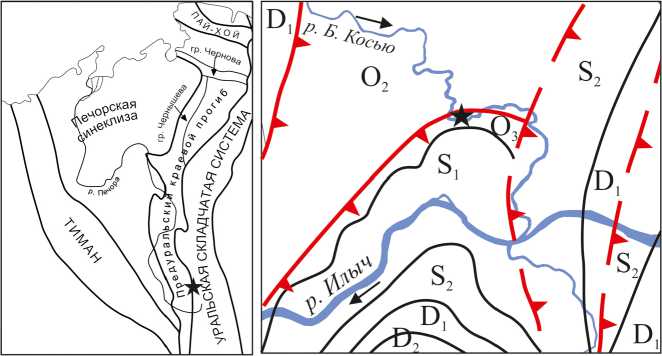
Рис. 1. Местонахождение рифа Большая Косью
Fig. 1. The location of the reef Bol'shaya Kos'yu леоландшафтный профиль рифа [6], получены новые данные о разнообразии биоты, в том числе установлено присутствие в рифовом сообществе многочисленных сфинктозоа. Так как зарождение и развитие рифа напрямую зависит от каркасообразующего потенциала организмов, способных создавать каркас, и структуры экосистемы в целом, определение их ассоциаций в рифовом сообществе имеет принципиальное значение. Цель статьи — охарактеризовать палеоэкосистемы на различных этапах формирования рифа, показать его структуру и особенности функционирования.
Стратиграфическое положение
Риф Большая Косью с тектоническим контактом залегает на глинистых сланцах, аргиллитах и петельчатых известняках большекосьюнской свиты, возраст которой соответствует верхней части сандбийского и нижней части катийского ярусов. Рифовая толща перекрывается комковатыми известняками с прослоями брекчий яптикшорской свиты верхнего катия, являющегося стратиграфическим аналогом кырьинско-го горизонта Урала [5]. Таким образом, риф Большая Косью соответствует верхнемалотавротинской подсвите Приполярного Урала, а его фациальным аналогом является сульфатно-доломитовая хорейверская свита, распространенная на западном склоне севера Урала.
Характерные представители рифового сообщества
Среди организмов, участвовавших в формировании рифового каркаса, кораллы (ругозы и табуляты) отмечаются довольно часто. Комплекс табулят на-
Й rD ^

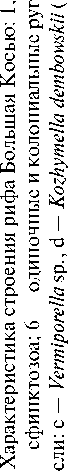
считывает 8 видов, которые принадлежат к 6 родам (рис. 2). Преобладают виды подотряда Heliolitina из массивных колоний куполовидной и пластинчатой форм размером от 30 х 5 до 15 х 20 см. Реже распространены виды подотряда Halysitina, которые встречаются в полостях рифа и характеризуются небольшими размерами. Комплекс ругоз, как одиночных, так и колониальных, насчитывает 5 видов, принадлежащих к 4 родам (рис. 2). Ругозы встречаются по всему разрезу рифовой толщи. Колониальные формы достигают в отдельных случаях довольно крупных размеров (60 х 40 см). Гидроиды Fistulella sp. чаще всего встречаются в виде изолированных колоний на склонах небольших биогермов, построенных в основном пластинчатыми и массивными табулятоморфными кораллами. Пространство между полипами гидроидов заполнено либо литокластовым и биокластовым материалом, либо инкрустационными спаритовыми цементами. Губки являются одной из основных групп каркасостроителей и представлены строматопорои-деями, сфинктозоа и соленопорами. Термин «сфинктозоа» (Sphinctozoa, синоним Thalamida) традиционно используется для камерных губок с жестким известковым скелетом. Соленопоры, традиционно относимые к красным водорослям, теперь также отнесены к губкам, так как, по существующему мнению [12], их внутренняя микроморфология наиболее близка к классу губок, так же, как и у строматопоро-идей [14]. Из-за того, что обломки строматопорои-дей и соленопор чаще всего изменены по краям сверлящими организмами, их таксономическая идентификация затруднена. В строении рифа они обычно встречаются в биокластовых разностях фланкирующих слоев биогермов. Сфинктозоа представлены видами Corymbospongia sp. и Amblysiphonelloides reticulata (Rigby & Potter), которые, согласно современным представлениям, относятся к классу обыкновенных губок Demospongiae Sollas, 1875. Corymbospongia sp. имеют сферические или субсферические камеры размером 1.0—1.8 см, которые часто располагаются одна над другой в виде ветви, либо формируют скопления неправильной конфигурации. Межкамерное пространство в колониях заселено обильными цианобактериями Renalcis . Губки A. reticulata (Rigby & Potter), которые имеют ветвистые колонии из кольцевидных (в сечении) камер с максимальной длиной ветви 120 мм и шириной до 60 мм. Камеры округлой формы, с четкими пережимами, наращиваются с небольшим обхватом. В структуре рифа сфин-ктозойные губки были широко распространены, но предпочитали скрытые полости. Рецептакулиты , по современным понятиям, представляют отдельную группу организмов, систематическое положение которых продолжает обсуждаться палеонтологами [11]. Их тела конической формы размером до 20 см в высоту и 14 см в поперечнике, хотя и достигали значительных размеров, не имели каркасообразующей роли, так как лишь спорадически отмечались на склонах небольших биогермов вместе с сфинктозоа A. reticulata (Rigby & Potter) и зелеными водорослями. Известковые водоросли представлены зелеными Vermiporella sp. и красными Kozhymella dembowskii (Sh.). Vermiporella sp. являются самыми распространенными не только в биогермах, но и в биокластовых 40
шлейфах. В биогермах встречаются в различных формах роста, обычно небольшие, хотя единичные экземпляры достигают нескольких сантиметров в длину и имеют хорошую сохранность. Красные водоросли K. dembowskii Sh. редки и встречаются в скрытых полостях рифа. Комплекс цианобактерий насчитывает 5 видов. Доминирующими являются Renalcis ex. gr. granosus (Vologd.), встречающиеся в виде корок на сфинктозойных губках Corymbospongia sp. и водорослях Vermiporella sp., K. dembowskii Sh. и в виде разрозненных колоний в биокластовых разностях. Girvanella ex. gr. problematica (Nich. et Ether.) встречаются вместе с зелеными водорослями в био-гермных известняках. Гирванеллоподобные скопления Botominella ex. gr. problematica (Nich. et Ether.) и Proaulopora ex. gr. glabra (Krasnop) редки и обычно присутствуют в биокластовых известняках вместе с фрагментами других водорослей и криноидей. Колониальные нитчатые водоросли Ikella sp. встречаются в виде однослойных пленок вокруг полипов гидроидов Fistulella sp. Широко распространены преимущественно пятиугольные членики криноидей Apertocrinus cf. apertus (Yelt. et Stuk.) и Bystrowicrinus cf. bilobatus (Yelt. et Stuk.).
Строение рифовой толщи
В строении рифа Большая Косью можно выделить 4 интервала разреза, соответствующие фазам/ стадиям развития рифа (рис. 2). И^mepваё I (17 м) характеризуют слои массивных биокластовых известняков, в которых встречаются небольшие линзы бра-хиопод и скопления табулят и ругоз. И^mepвaё II (19 м) представляют изолированные биогермы, достигающие 3—5 м в высоту, сложенные массивными биогермными известняками, а окружающее их пространство заполнено биокластовыми и литокластовыми известняками. Интервал III (75 м) формируют рифовые ядра из тесно расположенных и нарастающих друг на друга биогермов высотой 3—10 м. Биогермные известняки сложены разнообразными массивными каркасными организмами, в верхней части интервала распространены водорослевые и гидроидные разности. На склонах биогермов развиты биолитокластовые разности, редко — линзочки пелоидных известняков. Пелитоморфные известняки редки и встречаются в скрытых полостях биогермов. И^тeрвал IV (26 м) характеризуется увеличением в разрезе биолитокластовых разностей, среди которых отмечаются небольшие линзовидные биогермы мощностью до 3 м из пластинчатых табулят. Разрез рифовой толщи завершается плитчатыми биолитокластовыми известняками.
Характер строения рифовой толщи показывает, что слои интервала I отражают период формирования твердого скелетного известкового субстрата, сложенного преимущественно криноидным песком и цианобактериями. Линзы и скопления табулят, ругоз и брахиопод указывают на благоприятные условия для формирования здесь небольших органогенных банок. Биогермы, характерные для II, III и IV интервалов, имеют ясно выраженные различия в размерах и форме. Анализ таксономического разнообразия биоты позволил выявить следующие су- щественные отличия. Самое высокое биоразнообразие в рифовой палеоэкосистеме присуще III интервалу разреза с наиболее распространенными карка-состроителями — строматопороидеями, табулятами подотряда Heliolitina и сфинктозоа Corymbospongia sp. Для завершающей развитие рифа IV стадии отмечается самое низкое биоразнообразие. В целом по разрезу характерна смена преимущественно строматопо-роидных палеосообществ (интервал II) сфинктозой-но-табулятовыми (интервал IV), которая происходила постепенно в интервале III. Водоросли, одиночные и колониальные ругозы, гидроиды, губки (сфинктозоа A. reticulata, соленопоры) и рецептакулиты выступают в качестве второстепенных рифостроителей во II, III и IV интервалах разреза. Цианобактерии, такие как Renalcis, Girvanella, Botominella и Proaulopora, распространены повсеместно, Ikella — только в ассоциации с гидроидами Fistulella.
Обсуждение результатов и выводы
К началу среднего катия на северо-восточной континентальной окраине Европейской платформы оформился типичный карбонатный шельф, на окраине которого карбонатный осадок на песчаной отмели формировался в основном криноидеями (интервал I). Первыми способными формировать каркасную структуру были ругозы, табуляты, строматопорои-деи и сфинктозоа (интервал II). Небольшие мощности биогермных построек и присутствие в них цианобактерий и обильных известковых водорослей указывают на мелководные условия, в которых вертикальный рост построек зависел от колебания уровня моря, что приводило к ограничению пространства и низкому разнообразию организмов-каркасострои-телей. При устойчивом поднятии уровня моря пионерное сообщество сменилось таксономически более разнообразным, в количественном соотношении стали доминировать строматопороидеи и цианобактерии (нижняя часть интервала III), которые постепенно сменялись крупными массивными разнообразными кораллами в ассоциации с водорослями и цианобактериями. Колонии строматопороидей приобретают здесь черты угнетения, утончаясь в размерах до 5—10 см (верхняя часть интервала III). Это может быть объяснено тем, что при трансгрессии рост «вдогонку» (благоприятный для роста кораллов) становится преобладающим [3]. При достижении рифом уровня моря биогермы строились преимущественно угнетенными пластинчатыми колониями табулятоморф-ных кораллов. Во внутренних полостях этого каркаса селились сфинктозоа с цианобактериями и выступали в роли связывателей и укрепителей. При отсутствии внешних неблагоприятных факторов экосистема на этой стадии развития могла находиться достаточно долго. Причиной прекращения её развития была регрессия, которая фиксируется присутствием в кровле рифа плитчатых биолитокластовых отложений с присутствием вадозного цемента, характеризующего субаэральную обстановку [9].
Анализ разнообразия рифовых палеоценозов показывает, что структура палеоэкосистемы рифа Большая Косью является наиболее разнообразной по сравнению с одновозрастными рифами на
Приполярном и Полярном Урале [1]. Основными каркасостроителями исследуемого рифа являются губки, такие как строматопороидеи и сфинктозоа, и кораллы. В целом для позднего ордовика были характерны рифовые постройки из процветавших в то время строматопороидей. Что касается сфин-ктозойных губок, то ранее считалось, что они были важными рифостроителями в средней-поздней перми и среднем-позднем триасе [2]. В настоящее время эти морские бентосные организмы установлены в теплых низкоэнергетических сублиторальных обстановках островодужных террейнов кембрия Южной Австралии и США [8]. Предполагается, что по крайней мере до начала позднего ордовика эти обстановки были предпочтительной средой обитания сфинктозоа [7, 13]. В позднем ордовике происходит первая эволюционная радиация и географическое расселение сфинктозоа [7]. При этом изменялись и их экологические ниши от уровня дна до рифов. Впервые это было доказано на примере позд-некатийского сфинктозойно-кораллово-микроб-ного рифа юго-восточной части Китая [10]. Риф Большая Косью является вторым подобным примером. Сфинктозоа здесь были преимущественно криптобионтами, жили в средах, благоприятных для других метазоа, и не имели, таким образом, собственной экологической ниши. Однако при изменении условий обитания на неблагоприятные они, как резистентные для многих факторов организмы, становились одной из доминантных групп среди карка-состроителей рифа.
Автор выражает благодарность А. И. Антошкиной и Е. С. Пономаренко за консультации, ценные замечания и полезные рекомендации при подготовке статьи.
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 18-35-00331.
Список литературы Биоразнообразие верхнеордовикского рифа Большая Косью (Северный Урал)
- Антошкина А. И. Рифы в палеозое Печорского Урала. Санкт-Петербург: Наука, 1994. 154 с.
- Бойко Э. В., Беляева Г. В., Журавлева И. Т. Сфинктозоа фанерозоя территории СССР. М.: Наука, 1991. 224 с.
- Ивановский А. Б., Осипова А. И., Кузнецов В. Г. и др. Рифогенные постройки в палеозое России. Москва: Наука, 1997. 157 с.
- Кондиайн А. Г. Силурийские и нижнедевонские отложения Бельско-Елецкой фациальной зоны Печорского Урала // Тр. ВСЕГЕИ. Нов. сер. 1967. Т. 144. 87-122.
- Стратиграфические схемы Урала (докембрий, палеозой). Екатеринбург: Роскомнедра, ИГиГ УрО РАН, 1993.
