Биотопические группировки булавоусых чешуекрылых (Lepidoptera, Diuma) в экосистемах северной тайги Карелии
 в экосистемах северной тайги Карелии")
Автор: Резниченко Елена Сергеевна, Горбач Вячеслав Васильевич
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 2 (123), 2012 года.
Бесплатный доступ
Видовой состав и встречаемость дневных бабочек изучали в окрестностях г. Костомукши в течение 4 сезонов (2007-2010 годы). Всего найдены 46 видов. 587 особей 42 видов зарегистрированы в ходе 518 учетов на трансектах в 74 биотопах. Преобладали малинница, крапивница и репейница. Наиболее богатые группировки бабочек формируются на лугах, самые бедные - в сосняках. Отмечена слабая биотопическая дифференциация населения в условиях северной тайги.
Северная тайга карелии, булавоусые чешуекрылые, биотопическое распределение видов, состав и структура группировок
Короткий адрес: https://sciup.org/14750096
IDR: 14750096 | УДК: 574.472:595.789
Butterfly assemblages (Lepidoptera, Diurna) in ecosystems of northern taiga of Karelia
The species composition and occurrence of butterflies were studied in the area near Kostomuksha during the four years of research (2007-2010). A total of 46 species were found in the study area. 587 butterflies of 42 species were observed during 518 counts at 74 sites. Callophrys rubi, Nymphalis urticae and Vanessa cardu were prevailing species. The most numerous butterfly assemblages were formed in meadows and numerically insignificant butterfly assemblages - in pine forests. Poor differentiation of butterfly population in northern taiga is indicated.
Текст научной статьи Биотопические группировки булавоусых чешуекрылых (Lepidoptera, Diuma) в экосистемах северной тайги Карелии
Булавоусые, или дневные чешуекрылые, являются одной из хорошо исследованных групп насекомых. Вместе с тем изучение видового состава, распространения и особенностей экологии бабочек на конкретных территориях остается актуальной задачей. Большинство работ о булавоусых в Карелии посвящены южным районам, тогда как по территориям, расположенным к северу от Онежского озера, информации немного: опубликованная в середине прошлого века сводка Пелтонена [27] и по сей день остается основным источником для познания фауны бабочек северной тайги. Немногочисленные работы позднего периода [1], [8], [10], [11], [12], [13], [14], [17], [19] посвящены локальным фаунам Прибеломорья и международного российско-финляндского заповедника «Дружба». Всего из северотаежных провинций Карелии известно 59 видов булавоусых чешуекрылых, и данный список, по-видимому, близок к завершению [3]. Однако имеющиеся сведения о распространении и обилии видов фрагментарны, мало известно о закономерностях формирования биотопических группировок дневных бабочек в условиях северной тайги, их составе и структуре. Настоящая работа призвана восполнить недостаток знаний в этой области.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в окрестностях г. Костомукши, в том числе на территории государственного природного заповедника «Костомукшский» (64°35’ с. ш., 30°35’ в. д.). Природно-климатические условия данного района типичны для Западно-Карельской возвышенности. Преобладающий тип рельефа – денудационно-тектонический холмисто-грядовый. Озера занимают 8 % территории, болота – 18 %. Леса представлены сосняками, ельниками и мелколиственными насаждениями, покрывающими 49, 12 и 2 % всех земель соответственно. Около 9 % приходится на долю вырубок, пустырей, дорог и просек. Лугов и полей немного, менее 2 % в структуре ландшафта [2]. Среднегодовая температура воздуха составляет +0,5 °С при среднемесячных показателях от –15 °С в феврале до +14 °С в июле. Количество осадков достигает 550 мм в год [9].
На исследуемой территории выбрали 74 местообитания, представляющих все основные типы биотопов, заселяемых дневными бабочками в условиях северной тайги Карелии. Сфагновые болота включали массивы с доминированием в растительном покрове сосны, олиготрофных кустарничков (карликовой березы, багульника, мирта болотного и др.), осок и пушиц. В лесных биоценозах бабочки встречаются лишь на открытых участках – полянах и просеках. Подобные местообитания в различных типах сосновых лесов не отличались существенным образом по составу растительности, их характерная особенность – бедность разнотравья и присутствие багульника, голубики и других болотных кустарничков. Лиственные леса , сформированные березовыми и осиновыми насаждениями, напротив, имели хорошо развитую, разнообразную травянистую растительность по открытым местам. Исследованные луга представлены комплексами разнотравнозлаковых и влажноразнотравных группировок. Постоянными компонентами растительных ассоциаций выступали низовые злаки (полевица, белоус, душистый колосок, овсяница красная и др.), мелкое и среднее мезофильное разнотравье (манжетки, лютики, горошек мышиный,
чина луговая и др.). Растительность придорожных биотопов сочетала признаки как лесных, так и луговых фитоценозов. Отличительная черта таких местообитаний – обилие сорных и рудеральных растений (погремка, одуванчика, осота, бодяка, крапивы и др.).
Встречаемость бабочек в выбранных местообитаниях исследовали в 2007–2010 годах. Использовали метод учета имаго на трансектах [28]. Длина трансект в зависимости от размера местообитания варьировала от 50 до 150 м при ширине 5 м. Учеты проводили с мая по сентябрь раз в 7–10 дней в солнечную погоду. При необходимости бабочек отлавливали, определяли, а затем отпускали или собирали как эталонные экземпляры. В качестве биотопической группировки мы рассматривали совокупность видов, зарегистрированных в одном местообитании в течение всего периода исследований. Данные по встречаемости бабочек унифицировали посредством пересчета числа особей на единицу площади (экз./га). По полученным выборкам изучали видовой состав, плотность населения, видовое разнообразие и общность группировок, формирующихся в сходных условиях. Видовое разнообразие измеряли с помощью показателей энтропии Шеннона и доминирования Бергера – Паркера, общность – евклидовым расстоянием [5], [7]. Значимость отличий параметров оценивали посредством однофакторного дисперсионного анализа [6]. Видовое богатство группировок, относительную биотопическую приуроченность видов и тренды изменчивости населения бабочек северной тайги исследовали после объединения выборок, полученных в местообитаниях, отнесенных к одному типу биотопов. Видовое богатство описывали кривыми насыщения [16]. Приуроченность оценивали по частоте встреч вида в каждом типе биотопов. Критерий χ2 использовали для сравнения наблюдаемого и равномерного распределений частот. Статистический вывод о значимости различий между распределениями предполагает существование избирательности по отношению к местообитаниям. В таком случае в качестве излюбленного рассматривали местообитание с наиболее высокой встречаемостью особей. Сравнение частоты встреч вида в первом в иерархии местообитании с каждым следующим позволило выявить круг предпочитаемых местообитаний – группу, отличия между членами которой статистически незначимы. Поскольку данный метод нечувствителен к малым величинам, характеризующим встречаемость бабочек в наших выборках (экз./га), сохраняя принцип равенства площадей, мы использовали в расчетах действительные частоты встреч бабочек. Направления изменчивости населения изучали с помощью компонентного анализа [6]. В качестве значимых рассматривали главные компоненты с дисперсией больше единицы.
РЕЗУЛЬТАТЫ
В результате 518 учетов на 74 трансектах были зарегистрированы 587 особей 42 видов булавоусых чешуекрылых (табл. 1). Еще 4 вида – голубянка артаксеркс ( Aricia artaxerxes (Fabricius, 1793)), чернушка болотная ( Erebia embla (Thunberg, 1791)) и новые для карельской фауны червонцы непарный ( Lycaena dispar (Haworth, 1809)) и бурый ( L. tityrus (Poda, 1761)) найдены за пределами исследованных местообитаний. Доля трех наиболее многочисленных видов – малин-ницы, крапивницы и репейницы составила 41 % всех особей.
Биотопические группировки значимо отличались по числу видов (табл. 2: F = 10,91, df = 4,69, η 2 = 0,39, p < 0,001). Хорошей иллюстрацией изменчивости видового богатства населения служат кривые насыщения (рис. 1). Распадаясь на два кластера, они демонстрируют явное преимущество луговых и придорожных группировок чешуекрылых над лесными и болотными. Средняя плотность населения варьировала от 5,4 экз./га в сосняках до 27,2 экз./га на лугах ( F = 11,47, df = 4,69, η 2 = 0,40, p < 0,001). Сравнение структуры группировок чешуекрылых, формирующихся в биотопах одного типа, с помощью евклидовой меры показало высокую степень однородности населения сосняков и значительную изменчивость луговых группировок. Отличия между выборками по данному параметру значимы ( F = 138,37, df = 4,522, η 2 = 0,51, p < 0,001). Видовое разнообразие группировок, согласно индексу Шеннона, возрастало в направлении от сосняков и болот к лиственным лесам, придорожным биотопам и лугам ( F = 11,61, df = 4,69, η 2 = 0,40, p < 0,001). Самым многочисленным видом в болотных, лесных и придорожных местообитаниях является малинница, на лугах отмечено доминирование репейницы. Заметное участие в формировании луговых группировок, кроме того, принимали зорька, брюквенница, малин-ница и дневной павлиний глаз, а в придорожных биотопах чаще других встречались крапивница и репейница. Оценка видового разнообразия по индексу доминирования Бергера – Паркера показала, что наименее выравненная структура населения бабочек свойственна сосновым лесам, где около 40 % особей приходится на долю малинницы. В других местообитаниях происходит расширение круга видов с относительно высокой встречаемостью и значения показателя значимо уменьшаются, достигая минимума в луговых группировках чешуекрылых ( F = 9,26, df = 4,69, η 2 = 0,35, p < 0,001).
Относительная приуроченность к одному и более типам биотопов выявлена у 20 видов, их число варьирует от 1 в сосняках до 16 на лугах (табл. 1). Распространение многих видов по исследуемой территории сильно ограничено. Наиболее высокая локализация популяций отмечена для толстоголовки палаемон, червонца огненного, голубянки лесной, ленточника тополевого и углокрыльницы с-белое. Только на сфагновых болотах встречались перламутровки северная и эвномия, сенница болотная и сатир болотный, а голубянка эвмедон найдена лишь на лугах.
Метод главных компонент выявил одну значимую компоненту, учитывающую 75 % изменчивости исходной выборки (табл. 3). Все биотопические группировки имеют здесь примерно равные факторные нагрузки и составляют единственную плеяду.
Таблица 1
|
Вид |
Сфагновые болота, n = 11 |
Сосняки, n = 19 |
Лиственные леса, n = 15 |
Луга, n = 15 |
Придорожные биотопы, n = 14 |
||||||||||
|
M |
S |
Ps |
M |
S |
Ps |
M |
S |
Ps |
M |
S |
Ps |
M |
S |
Ps |
|
|
Pyrgus malvae (Linnaeus, 1758) – Толстоголовка мальвовая |
– |
– |
– |
– |
– |
– |
0,2 |
0,7 |
13 |
0,5 |
1,3 |
13 |
– |
– |
– |
|
Carterocephalus silvicola (Meigen, 1829) – Т. лесная желтая* |
0,4 |
1,2 |
10 |
– |
– |
– |
0,2 |
0,7 |
13 |
– |
– |
– |
– |
– |
– |
|
C. palaemon (Pallas, 1771) – Т. палемон* |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
– |
– |
– |
|
Thimelicus lineola (Ochenheimer, [1808]) – Т.-тире |
– |
– |
– |
0,2 |
0,6 |
11 |
– |
– |
– |
0,8 |
1,4 |
27 |
– |
– |
– |
|
Papilio machaon Linnaeus, 1758 – Махаон* |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,2 |
0,7 |
13 |
0,1 |
0,5 |
7 |
|
Leptidea sinapis (Linnaeus, 1758) – Беляночка горошковая |
– |
– |
– |
– |
– |
– |
0,2 |
0,9 |
7 |
0,8 |
1,2 |
33 |
0,3 |
0,7 |
14 |
|
Anthocharis cardamines – Зорька |
– |
– |
– |
– |
– |
– |
0,6 |
1,2 |
27 |
2,4 |
2,7 |
53 |
1,8 |
2,6 |
36 |
|
Aporia crataegi (Linnaeus, 1758) – Боярышница* |
– |
– |
– |
– |
– |
– |
0,2 |
0,7 |
13 |
1,0 |
1,7 |
33 |
0,5 |
1,2 |
21 |
|
Pieris brassicae (Linnaeus, 1758) – Капустница* |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
0,1 |
0,5 |
7 |
0,3 |
0,7 |
14 |
|
P. rapae (Linnaeus, 1758) – Репница* |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,2 |
1,0 |
7 |
0,1 |
0,5 |
7 |
|
P. napi (Linnaeus, 1758) – Брюквенница |
– |
– |
– |
– |
– |
– |
0,6 |
1,2 |
27 |
1,8 |
3,3 |
33 |
1,1 |
1,9 |
29 |
|
Colias palaeno (Linnaeus, 1761) – Желтушка торфяниковая* |
0,4 |
0,8 |
20 |
0,5 |
1,1 |
21 |
0,5 |
1,1 |
20 |
0,2 |
1,0 |
7 |
– |
– |
– |
|
Gonepteryx rhamni (Linnaeus, 1758) – Крушинница |
– |
– |
– |
– |
– |
– |
1,0 |
1,4 |
40 |
1,0 |
2,1 |
27 |
0,7 |
1,6 |
21 |
|
Callophrys rubi (Linnaeus, 1758) – Малинница |
1,9 |
1,7 |
70 |
2,2 |
2,0 |
68 |
3,4 |
3,3 |
80 |
3,0 |
3,2 |
67 |
4,2 |
3,0 |
86 |
|
Lycaena virgaurea (Linnaeus, 1758) – Червонец огненный* |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
– |
– |
– |
|
Celastrina argyolus (Linnaeus, 1758) – Голубянка весенняя |
0,5 |
1,2 |
20 |
0,1 |
0,4 |
5 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Plebejus argus – Г. аргус* |
0,5 |
1,2 |
20 |
0,3 |
0,7 |
16 |
0,1 |
0,5 |
7 |
0,5 |
1,1 |
20 |
0,1 |
0,5 |
7 |
|
P. idas (Linnaeus, 1761) – Г. идас* |
0,2 |
0,6 |
10 |
0,1 |
0,4 |
5 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
P. optilete (Knoch, 1781) – Г. торфянико-вая* |
0,4 |
1,2 |
10 |
0,4 |
1,0 |
16 |
– |
– |
– |
– |
– |
– |
0,3 |
0,7 |
14 |
|
Aricia eumedon (Esper, [1780]) – Г. эвме-дон |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,4 |
1,1 |
13 |
– |
– |
– |
|
Polyommatus icarus (Rottemburg, 1775) – Г. икар* |
0.4 |
0.8 |
20 |
– |
– |
– |
– |
– |
– |
0,2 |
0,7 |
13 |
– |
– |
– |
|
P. semiargus (Rottemburg, 1775) – Г. лесная* |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
– |
– |
– |
– |
– |
– |
|
Limenitis populi (Linnaeus, 1758) – Ленточник тополёвый* |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
– |
– |
– |
– |
– |
– |
|
Nymphalis antiopa (Linnaeus, 1758) – Траурница* |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
0,1 |
0,5 |
7 |
|
N. io (Linnaeus, 1758) – Дневной павлиний глаз |
– |
– |
– |
– |
– |
– |
0,9 |
2,5 |
20 |
2,0 |
3,0 |
40 |
1,2 |
2,2 |
36 |
Видовой состав и плотность населения (экз./га) булавоусых чешуекрылых в местообитаниях северной тайги Кар елии
Примечание. n – число исследованных местообитаний, M – средняя арифметическая, S – стандартное отклонение, Ps – доля местообитаний, где был отмечен вид (встречаемость, %), жирным шрифтом указана относительная биотопическая приуроченность вида, * – вид не обнаружил значимой приуроченности ни к одному из биотопов.
Продолжение табл. 1
|
Виды |
Сфагновые болота, n = 11 |
Сосняки, n = 19 |
Лиственные леса, n = 15 |
Луга, n = 15 |
Придорожные биотопы, n = 14 |
||||||||||
|
M |
S |
Ps |
M |
S |
Ps |
M |
S |
Ps |
M |
S |
Ps |
M |
S |
Ps |
|
|
N. urticae (Linnaeus, 1758) – Крапивница |
– |
– |
– |
0,4 |
1,0 |
16 |
1,1 |
2,6 |
27 |
1,5 |
2,8 |
33 |
2,4 |
3,6 |
43 |
|
Polygonia c-album (Linnaeus, 1758) – Углокрыльница с-белое* |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
|
Vanessa atalanta (Linnaeus, 1758) – Адмирал |
– |
– |
– |
0,1 |
0,4 |
5 |
0,6 |
2,0 |
13 |
1,4 |
3,0 |
20 |
0,7 |
1,8 |
14 |
|
V. cardui (Linnaeus, 1758) – Репейница |
1,0 |
1,3 |
50 |
0,2 |
0,6 |
11 |
1,8 |
2,0 |
60 |
4,4 |
2,8 |
87 |
3.5 |
2,5 |
86 |
|
Arashnia levana (Linnaeus, 1758) – Пестрокрыльница |
– |
– |
– |
– |
– |
– |
0,5 |
1,1 |
20 |
0,5 |
1,1 |
20 |
– |
– |
– |
|
Brenthis ino (Rottemburg, 1775) – Перламутровка-таволжанка* |
– |
– |
– |
0,3 |
1,0 |
11 |
0,6 |
1,4 |
20 |
0,5 |
1,3 |
13 |
0,4 |
1,1 |
14 |
|
Boloria aquilonaris (Stichel, 1908) – П. северная |
0,9 |
1,3 |
40 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Clossiana eunomia (Esper, [1799]) – П. эв-номия |
0,5 |
0,9 |
30 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
C. euphrosyne (Linnaeus, 1758) – П. эфро-сина |
0,2 |
0,6 |
10 |
0,2 |
0,6 |
11 |
– |
– |
– |
0,9 |
1,3 |
27 |
0,3 |
1,0 |
7 |
|
C. selene ([Dennis & Schiffermüller], 1775) – П. селена* |
0,2 |
0,6 |
10 |
0,1 |
0,4 |
5 |
0,1 |
0,5 |
7 |
0,4 |
1,1 |
13 |
0,5 |
1,2 |
21 |
|
Pararge maera (Linnaeus, 1758) – Бархатка большая* |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
– |
– |
– |
0,3 |
1,0 |
7 |
|
P. petropolitana (Fabricius, 1787) – Б. петербургская* |
– |
– |
– |
0,1 |
0,4 |
5 |
– |
– |
– |
– |
– |
– |
0,3 |
0,7 |
14 |
|
Coenonympha tullia – Сенница болотная |
0,5 |
0,9 |
30 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
C. pamphilus (Linnaeus, 1758) – С. обыкновенная* |
0,5 |
1,2 |
20 |
– |
– |
– |
0,1 |
0,5 |
7 |
0,4 |
0,8 |
20 |
0,4 |
1,1 |
14 |
|
Aphantopus hyperantus (Linnaeus, 1758) – Глазок цветочный |
– |
– |
– |
– |
– |
– |
0,1 |
0,5 |
7 |
0,6 |
1,6 |
20 |
0,1 |
0,5 |
7 |
|
Erebia ligea (Linnaeus, 1758) – Чернушка кофейная |
0,2 |
0,6 |
10 |
0,2 |
0,6 |
11 |
1,0 |
1,7 |
33 |
0,6 |
1,6 |
20 |
1,1 |
3,0 |
21 |
|
Oeneis jutta (Hübner, [1806]) – Сатир болотный* |
0,2 |
0,6 |
10 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
Таблица 2
Характеристики биотопических группировок чешуекрылых
|
Показатель |
Сфагновые болота, n = 11, *n = 55 |
Сосняки, n = 19, *n = 171 |
Лиственные леса, n = 15, *n = 105 |
Луга, n = 15, *n = 105 |
Придорожные биотопы, n = 14, *n = 91 |
|
|
Число видов |
M S |
3,5 |
2,2 |
4,8 |
7,1 |
5,6 |
|
2,4 |
1,4 |
2,4 |
2,5 |
2,8 |
||
|
Плотность населения, |
M |
8,7 |
5,4 |
14,7 |
27,2 |
21,1 |
|
экз./га |
S |
6,6 |
4,1 |
10,4 |
14,5 |
13,1 |
|
Общность группировок, |
M |
0,137 |
0,104 |
0,209 |
0,313 |
0,262 |
|
евклидово расстояние |
S |
0,040 |
0,043 |
0,103 |
0,093 |
0,100 |
|
Видовое разнообразие, |
M |
0,55 |
0,37 |
0,71 |
0,89 |
0,77 |
|
индекс Шеннона |
S |
0,32 |
0,26 |
0,24 |
0,16 |
0,22 |
|
Видовое разнообразие, |
M |
0,53 |
0,72 |
0,40 |
0,27 |
0,37 |
|
индекс Бергера – Паркера |
S |
0.33 |
0,31 |
0,20 |
0,07 |
0,17 |
Примечание. n – число группировок, *n – число сравниваемых пар для оценки общности группировок, M – средняя арифметическая, S – стандартное отклонение.
Ординация объектов показывает, что виды расположились вдоль оси рассматриваемой компоненты согласно частоте их встречаемости (рис. 2). Наименьшие значения получили редкие виды, такие как толстоголовка палаемон и махаон, а наибольшие значения остались за доминантами – малинницей, репейницей и крапивницей. Информативность второй компоненты оценена в 16 %. И хотя формально она незначима (дисперсия меньше 1), без учета содержащейся в ней информации анализ нельзя признать полным. Именно здесь проявилась тенденция к противопоставлению группировок бабочек, формирующихся на болотах и в сосняках, луговым группировкам. Таким образом, первая ось описывает встречаемость видов в условиях северной тайги, общую структуру фауны, а вторая намечает направления биотопической дифференциации населения.
Таблица 3
Факторные нагрузки и дисперсии главных компонент
|
Биотопические группировки |
ГК-1 |
ГК-2 |
|
Сфагновые болота |
0,389 |
-0,587 |
|
Сосняки |
0,430 |
-0,480 |
|
Лиственные леса |
0,495 |
0,096 |
|
Луга |
0,426 |
0,575 |
|
Придорожные биотопы |
0,488 |
0,292 |
|
Дисперсия |
3,75 |
0,81 |
|
Дисперсия, % |
75,0 |
16,2 |
Примечание. ГК – главная компонента.
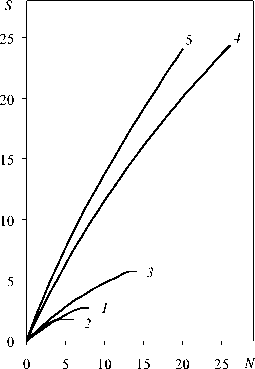
Рис. 1. Насыщенность видами биотопических группировок чешуекрылых:
1 – сфагновые болота, 2 – сосняки, 3 – лиственные леса, 4 – луга, 5 – придорожные биотопы. N – число особей, S – число видов
-
1 .0
0.5
0.0
-0.5
-1.0
-1.5
crat
napi
atal
♦ rham
io
carm card
urt
hvn/ sina hype malv leva
♦ bras ♦ maer ♦ pet silv
line
lige
idas
icar ♦ jut
♦ euph sele
pamp
ino
tul I euno
♦ argy
opt
argu
♦ pala
aqui
rub
-
-1.5 -1.0 -0.5 0.0 0.5 1.0 _ - 1
Рис. 2. Ординация чешуекрылых в осях главных компонент: виды обозначены первыми литерами их латинского названия, для обозначения V. cardui принято сокращение card, для A. cardamines – carm. У видов, помещенных на границы диаграммы, уменьшены значения соответствующих главных компонент. Эллипсом выделена группа видов, включающая C. palaemon , P. machaon , P. rapae , H. virgaurea , A. eumedon , C. semiargus , L. populi , N. antiopa и P. c-album
ОБСУЖДЕНИЕ
46 видов булавоусых чешуекрылых, найденных в окрестностях г. Костомукши, составляют 77 % всех видов, известных из северной тайги Карелии [3]. Лишь 14 видов не отмечены нами на исследуемой территории. Прежде всего это толстоголовка северная ( Pyrgus centaureae (Ram-bur, 1839)), червонец голубоватый ( Lycaena helle ([Dennis&Schiffermüller], 1775)), голубянка ни-кия ( Aricia nicias (Meigen, 1830)), перламутровки фрея ( Clossiana freija (Becklin in Thunberg, 1791)) и фригга ( C. frigga (Becklin in Thunberg, 1791)), типичные виды для местных условий, но имеющие локальное распространение [13]. Остальные виды северотаежной фауны – толстоголовка-запятая ( Hesperia comma (Linnaeus, 1758)), зефир березовый ( Thecla betulae (Linnaeus, 1758)), червонцы пятнистый ( Lycaena phlaeas (Linnaeus, 1761)) и щавелевый ( L. hippothoe (Linnaeus, 1758)), перламутровки аглая ( Argynnis aglaja (Linnaeus, 1758)) и полевая ( Issoria lathonia (Linnaeus, 1758)), шашечница аталия ( Melitaea athalia (Rottemburg, 1775)), сенница луговая ( Coenonympha glycerion (Borkhausen, 1788)) и чернушка диса ( Erebia disa (Becklin in Thunberg, 1791)) отмечены для сопредельных районов по немногим находкам [27]. Впервые на исследуемой территории зарегистрирован 31 вид. Почти все эти бабочки известны для запада северной тайги по опубликованным данным. Находку непарного и бурого червонцев мы склонны рассматривать как результат случайного заноса, поскольку северные границы их ареалов проходят на 300–500 км южнее [22]. Тем не менее нельзя полностью исключать и возможность естественного проникновения этих видов на север в результате экспансии, которую в последние годы наблюдали у многих чешуекрылых [26], [30]. В целом же результаты исследования свидетельствуют о бедности северотаежной фауны дневных бабочек. С одной стороны, это связано с тем, что распространение ряда обычных для Карелии видов в северном направлении ограничено 62–63° с. ш. Те из них, что проникают в северную тайгу, попадаются крайне редко. С другой стороны, нет здесь и арктических бабочек, которые не встречались бы в среднетаежных провинциях. Исключение составляют лишь чернушка диса, известная из района Калевалы, и северная форма толстоголовки-запятой (f. geogr. catena ), указанная для Соловецкого архипелага [27].
Количественные оценки населения булавоусых чешуекрылых исследуемой территории оказались существенно ниже, чем в среднетаежных ландшафтах: например, средняя частота встреч имаго в местообитаниях Прионежья составляла 57 экз./га [15] против 15 экз./га в рассматриваемом случае. Низкое обилие дневных бабочек обусловлено, по-видимому, не только природно-климатическими особенностями рассматриваемого района, но и крайне неблагопри- ятными для развития большинства видов погодными условиями в летние месяцы 2007–2008 годов [4]. Преобладание малинницы и крапивницы характерно и для южных провинций Карелии, другие доминантные виды среднетаежных ландшафтов – брюквенница, крушинница, перламутровка-таволжанка, глазок цветочный и чернушка кофейная – на исследуемой территории попадались гораздо реже. Высокая численность репейницы – явление в наших широтах редкое, обусловленное интенсивной миграцией особей на север в годы с теплым летом. В такие сезоны в таежных местообитаниях чаще встречаются и другие мигранты (адмирал, дневной павлиний глаз и т. д.).
Бабочки очень неравномерно размещены по исследуемой территории, так же как и в средней тайге, населяя лишь более или менее открытые местообитания – болота, луга, поляны, просеки, вырубки и обочины дорог. Однако северотаежные группировки булавоусых чешуекрылых заметно беднее. Число видов, зарегистрированных в местообитании, здесь не превышает 11, тогда как в Северо-Западном Прионежье этот показатель достигает 21 вида [15], в Северном Прила-дожье – 38 [21], [25], а на юге Финляндии – 46 [18]. Наблюдаемое обеднение биотопических группировок происходит вследствие уменьшения встречаемости, а затем и полного выпадения видов из состава локальных фаун. Еще более заметно в северном направлении меняется плотность населения бабочек. В основных типах среднетаежных местообитаний среднее число зарегистрированных на трансектах особей составляло 45–101 [23], [29] и 35–88 экз./га [15], тогда как в северотаежных местообитаниях значение этого показателя не превышало 28 экз./га. Анализ структуры населения показал, что в северотаежных группировках чешуекрылых относительно высокую встречаемость имеют немногие виды, прежде всего те, что указаны в качестве доминирующих для исследуемой фауны. Использованные меры видового разнообразия вполне адекватно регистрируют выявленные градиенты. Значения энтропии Шеннона по сравнению с Прионежьем [15] снижаются с 1,6–2,27 до 0,37–0,88, а значения индекса доминирования Бергера – Паркера возрастают с 0,22–0,42 до 0,27–0,72. Вместе с тем иерархия биотопических группировок бабочек по числу видов, плотности населения и видовому разнообразию остается постоянной независимо от географического положения упомянутых районов: наиболее бедные группировки в условиях таежной зоны формируются в лесных и болотных местообитаниях, а наиболее богатые – на лугах. Лучшей иллюстрацией общей тенденции служат показанные на рис. 1 кривые насыщения группировок видами.
Биотопические предпочтения видов формируются под влиянием многих факторов, важнейшими из которых являются наличие в достаточном количестве пищевых ресурсов и благоприятные мезо- и микроклиматические условия. Соответствие условий среды экологической специфике каждого вида обусловливает свойственное только ему биотопическое распределение. Одни виды на исследуемой территории сохраняют строгую приуроченность к очень узкому набору местообитаний. Примером могут служить сенница болотная, перламутровки северная и эвномия, которые в условиях лесной зоны облигатно связаны с болотными биоценозами [20]. Другие виды демонстрируют более равномерное биотопическое распределение, чем в среднетаежных ландшафтах. Некоторые из них, такие как желтушка торфянниковая, голубянка аргус, сенница обыкновенная, перламутровки таволжанка и селена, не обнаруживают значимой приуроченности ни к одному из рассматриваемых типов местообитаний, у малин-ницы биотопические предпочтения смещаются от болот к придорожным биотопам. Приуроченность многих бабочек к лугам и обочинам дорог объясняется прежде всего богатством растительности, в которой в достаточном количестве присутствуют кормовые растения гусениц и нектароносные растения для питания имаго. Болота и сосновые леса вследствие специфичности среды наименее благоприятны для развития дневных бабочек, поэтому к этим местообитаниям тяготеют немногие виды.
Обобщающий анализ размещения чешуекрылых не выявил значимых трендов изменчивости населения исследуемой территории, деления их на лесные, болотные и луговые, как ранее было показано для среднетаежных ландшафтов [15], [24]. Метод главных компонент зафиксировал лишь тенденцию пространственной дифференциации населения. При этом многочисленные виды обнаружили тяготение к тем же типам биотопов, что и в средней тайге. Даже малин-ница, анализ размещения которой показал приуроченность к придорожным биотопам, здесь попала в категорию лесных и болотных видов, каким по сути и является. Это связано с тем, что взаиморасположение видов в осях главных компонент, отражая корреляционные связи между биотопическими группировками, определяется не столько абсолютными значениями плотности, сколько участием вида в формировании структуры населения местообитаний. Причины отсутствия явных трендов кроются в особенностях природно-климатических условий района исследований. Во-первых, здесь относительно мало лугов и луговые виды бабочек вынуждены использовать все более или менее подходящие для существования открытые местообитания, в том числе и лесные. Во-вторых, отсутствует четкое разграничение между растительным покровом болотных и лесных местообитаний;
под пологом леса, на полянах и просеках присутствует целый ряд типичных болотных растений – обычное явление для северной тайги. Как следствие жесткость связи тирфобионтов и тир-фофилов с болотами ослабевает. В-третьих, поскольку в северной тайге в силу климатических условий численность дневных бабочек редко бывает высокой даже в наиболее благоприятных для развития местообитаниях, то и пространственное размещение большинства видов отличается большей выравненностью, чем их размещение на юге Карелии.
Список литературы Биотопические группировки булавоусых чешуекрылых (Lepidoptera, Diuma) в экосистемах северной тайги Карелии
- Болотов И. Н., Шутова Е. В. Закономерности формирования островных фаун булавоусых чешуекрылых (Lepidoptera, Diuma) на северном пределе распространения лесов в области плейстоценовых материковых оледенений (на примере островов Белого моря)//Известия РАН. Сер. биологическая. 2006. Вып. 3. С. 327-336.
- Волков А. Д., Громцев А. Н., Еруков Г. В. и др. Экосистемы ландшафтов запада северной тайги (структура, динамика). Петрозаводск, 1995. 194 с.
- Горбач В. В., Резниченко Е. С. Видовой состав и распространение дневных бабочек (Lepidoptera, Diurna) в ЮгоВосточной Фенноскандии//Ученые записки Петрозаводского государственного университета. Сер. «Естественные и технические науки». 2009. № 7 (101). С. 31-39.
- Горбач В. В., Сааринен К., Резниченко Е. С. К экологии тополевого ленточника Limenitispopuli (Lepidoptera, Nymphalidae) Восточной Фенноскандии//Зоологический журнал. 2010. Т. 89. № 11. С. 1340-1349.
- Коросов А. В. Специальные методы биометрии. Петрозаводск: Изд-во ПетрГУ, 2007. 364 с.
- Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных. Петрозаводск: Изд-во ПетрГУ, 2010. 84 с.
- Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 181 с.
- Полевой А. В., Хумала А. Э. Насекомые//Материалы инвентаризации природных комплексов и научное обоснование ландшафтного заказника «Сыроватка». Петрозаводск: Карельский научный центр РАН, 2003. С. 67-72.
- Романов А. А. О климате Карелии. Петрозаводск: Госиздат КАССР, 1961. 140 с.
- Свиридов А. В. Чешуекрылые (Lepidoptera, Macrolepidoptera) окрестностей Беломорской биологической станции Московского государственного университета и их стациальное распределение//Энтомологическое обозрение. 1970. Т. 49. Вып. 3. С. 365-372.
- Хумала А. Э. Изучение энтомофауны островных экосистем Онежской губы Белого моря//Природное и культурное наследие Северной Фенноскандии. Петрозаводск: Карельский научный центр РАН, 2003. С. 83-89.
- Хумала А. Э., Полевой А. В. К фауне насекомых Карельского побережья и островов Белого моря//Инвентаризация и изучение биологического разнообразия на Карельском побережье Белого моря. Петрозаводск, 1999. С. 106-113.
- Шутова Е. В., Антонова Е. М., Свиридов А. В., Кутенкова Н. Н. Чешуекрылые Кандалакшского заповедника//Флора и фауна заповедников СССР. Вып. 80. М., 1999. 48 с.
- Яковлев Е. Б., Хумала А. Э., Полевой А. В. Насекомые//Инвентаризация и изучение биологического разнообразия на территории центральной Карелии. Петрозаводск: Карельский научный центр РАН, 2001. С. 149-158.
- Gorbach V. V., Saarinen K. The butterfly assemblages of Onega Lake Area in Karelia, middle taiga of NW Russia (Hes-perioidea, Papilionoidea)//Nota lepid. 2003. Vol. 25. P. 267-279.
- Gotelli N., Colwell R. Quantifying biodiversity: procedures and pitfalls in measurement and comparison of species richness//Ecology Letters. 2001. Vol. 4. P. 379-391.
- Heikkilä R., Lindholm T. Biodiversity and conservation of boreal nature. Kainuu, 2003. P. 187-192.
- Kuussaari M., Heliölä J., Salminen J., Niininen I. Maatalousympäristön päiväperhosseurannan vuoden 2000 tulokset//Baptria. 2001. Vol. 26. P. 69-80.
- Leinonen R., Itamies J., Kutenkova N. Lepidoptera of the Nature Reserve Friendship//Ecosystems, fauna and flora of the Finnish-Russian Nature Reserve Friendship. Helsinki: The Finnish Environment, 1997. P. 235-256.
- Mikkola K., Spitzer K. Lepidoptera associated with peatlands in central and northern Europe: a synthesis//Nota lepid. 1983. Vol. 6. P. 216-229.
- Saarinen K. A comparison of butterfly communities along field margins under traditional and intensive management in SE Finland//Agriculture, Ecosystems and Environment. 2002. Vol. 90. P. 59-65.
- Saarinen K., Gorbach V. V. Erikoisia kultasiipiä itärajan tuntumassa//Baptria. 2007. Vol. 32. P. 15.
- Saarinen K., Jantunen J. A comparison of butterfly fauna of agricultural habitats under different management history in Finnish and Russian Karelia//Ann. Zool. Fenn. 2002. Vol. 39. P 173-181.
- Saarinen K., Marttila O., Jantunen J. Species richness and distribution of butterflies (Lepidoptera: Hesperioidea, Papilionoidea) in agricultural environment in SE Finland//Entomol. Fenn. 1998. Vol. 9. P. 9-18.
- Saarinen K., Valtonen A., Jantunen J., Saarnio S. Butterflies and diurnal moths along road verges: Does road type affect diversity and abundance?//Biol. Conserv. 2005. Vol. 123. P 403-412.
- Parmesan C., Ryrholm N., Stefanescu C. et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming//Nature. 1999. Vol. 399. P. 579-583.
- Peltonen O. Vienan perhosfaunasta//Ann. Entomol. Fenn. 1947. Vol. 13. S. 131-144.
- Pollard E., Yates T. J. Monitoring butterflies for ecology and conservation. The British butterfly monitoring scheme. London: Chapman & Hall, 1993. 274 p.
- Väisänen R. Distribution and abundance of diurnal Lepidoptera on a raised bog in southern Finland//Ann. Zool. Fenn. 1992. Vol. 29. P. 75-92.
- Warren M. S., Hill J. K., Thomas J. A. et al. Rapid responses of British butterflies to opposing forces of climate and habitat change//Nature. 2001. Vol. 414. P. 65-69.
