Близкородственные виды Allium erubescens C. Koch. и Allium rotundum L. (Allium, Alliaceae) с точки зрения формирования жизненной стратегии
 с точки зрения формирования жизненной стратегии")
Автор: Чадаева Виктория Александровна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 4, 2016 года.
Бесплатный доступ
Выявлены особенности формирования жизненной стратегии морфологически схожих видов A. rotundum и A. erubescens в природе. Показано, что организменные и популяционно-онтогенетические механизмы устойчивости данных видов, в комплексе определяющие тип их стратегии выживания, в схожих условиях произрастания аналогичны и не могут быть использованы в качестве диагностических таксономических признаков.
Ценопопуляция, механизмы устойчивости, стратегия жизни видов
Короткий адрес: https://sciup.org/146116671
IDR: 146116671 | УДК: 581.5;
Closely related species, Allium erubescens C. Koch. and Allium rotundum L. (Allium, Alliaceae), and their life strategies
We describe life strategies of two closely related wild species, A. rotundum и A. erubescens. Their life strategies are similar under the same conditions. Thus they can not be used to diagnose species.
Текст научной статьи Близкородственные виды Allium erubescens C. Koch. и Allium rotundum L. (Allium, Alliaceae) с точки зрения формирования жизненной стратегии
А.И. Введенский (1935) во «Флоре СССР» разграничивает A. rotundum и A. erubescens по признакам окраски и формы листочков околоцветника. При этом описание видов изобилует уточнениями «чаще», «реже», «обычно», «иногда» и т. п. По признанию самого автора, в работе была произведена попытка до известной степени насильственного разделения видов внутри группы A. rotundum s. 1, так как другие признаки по имеющемуся, часто плохо собранному гербарному материалу, использовать не удалось. Для A. rotundum указываются темнопурпурные с более темной жилкой, тупые или туповатые наружные листочки околоцветника и более светлые, чаще почти белые, с пурпурной жилкой, очень тупые внутренние листочки, для И. erubescens - розовые с пурпурной жилкой, острые, иногда на верхушке слегка отогнутые листочки.
А.И. Галушко (1978) при составлении сводки «Флора Северного Кавказа» в качестве диагностических признаков, кроме указанных, использует высоту растений - 30-60 см у A. rotundum и 1530 см у A. erubescens. У второго вида автором отмечается рыхлое соцветие.
Более подробно диагностическое значение морфологических признаков данных видов изучено А.П. Серегиным (2007). В качестве морфологических маркеров для дифференциации A. rotundum и A. erubescens автором использовались: форма соцветия (соответственно густое, шаровидное и довольно рыхлое, полушаровидное), цвет листочков околоцветника (варьируется от интенсивно темно-пурпурного до почти розового, жилка менее выражена и обычно розоватый, иногда темно-пурпурный с более темной жилкой), форма верхушки листочков (менее острая и более острая), параметры листьев (до 7-8 мм шириной и обычно узколинейные, менее 2 мм шириной), высота растения (выше 30 см и до 30(40) см).
В то же время, многочисленными исследователями отмечена значительная изменчивость морфологических признаков A. rotundum, в том числе и окраски долей околоцветника (Чешмеджиев, 1979; Кудряшова, 1990; Погосян, 1992). В качестве причины фенотипического многообразия вида в статье Е.С. Немировой и И.В. Троцкой (2008) указывается наличие в популяциях как диплоидных, так и полиплоидных форм, появление которых в пределах всего ареала, возможно, объясняется действием климатических и эдафических факторов. Поэтому диагностическое значение окраски листочков околоцветника A. rotundum можно считать небольшим.
Соответственно существуют разногласия относительно ареала распространения A. erubescens. По А.А. Гроссгейму (1940), этот вид широко распространен на Кавказе. А.И. Введенский (1935) указывает на редкость A. erubescens в Крыму, А.П. Серегин (2005) полностью исключает его из флоры полуострова. Республикой Дагестан ограничивает распространение A. erubescens на Кавказе Б. Мэтью (Mathew, 1996). А.П. Серегин (2007) отмечает наличие вида на территории Дагестана и Чеченской Республики. В предварительном обзоре видов рода Allium L. для издания «Конспекта флоры Кавказа» Г.Л. Кудряшовой (1992) A. erubescens указан для Восточного Кавказа, Восточного и Южного Закавказья. По мнению автора, упоминания данного вида И.С. Косенко (1970), А.И. Галушко (1978), ВТ.
Танфильевым и В.Н. Кононовым (1987) на территории Западного Предкавказья и Западного Кавказа относятся к одной из изменчивых форм Л. rotundum. Согласно мнению А.С. Зернова, A. erubescens также характерен для Восточного Кавказа, во флоре Западного Кавказа данный вид отсутствует (Зернов, 2010).
Нами проведен сравнительный анализ стратегии выживания A. erubescens и A. rotundum с целью установления возможности использования особенностей жизненной стратегии в качестве дополнительного диагностического признака при дифференциации двух видов. При этом под стратегией выживания вида понимаем интегральную генетически обусловленную характеристику, комплексный адаптивный ответ вида на внешние воздействия, формирующийся благодаря тесной взаимосвязи организменных и популяционно-онтогенетических механизмов устойчивости (Шхагапсоев, Чадаева, 2015). Организменные механизмы устойчивости обеспечивают гетерогенность ценопопуляций (ЦП): биоморфологическая, возрастная и размерная дифференциация особей, семенное размножение, обусловливающее генетическое разнообразие растений. Популяционно-онтогенетические механизмы устойчивости обеспечивают лабильность ценопопуляций в неоднородных условиях среды: онтогенетические тактики, онтогенетические и репродуктивные стратегии, адаптивная изменчивость возрастной, виталитетной, пространственной и биоморфологической структур, плотности и численности, жизненного состояния ценопопуляций, ритмов фенологического развития.
Методика. Исследования проводились нами в период с 2008 по 2015 гг. На территории Кабардино-Балкарской и Карачаево-Черкесской Республик, Республик Северной Осетии-Алании обследовано 14 ценопопуляций (ЦП) A. rotundum, на территории Республики Дагестан - 11 ЦП A. erubescens - многолетних, луковичных, короткостолонообразующих поликарпических видов семейства Alliaceae J. Agardh., выделенного как самостоятельное из обширного семейства Liliaceae L. (Тахтаджян, 1966) (табл. 1).
В качестве счетной единицы использовали морфологическую особь. Возрастные состояния растений выделены на основании комплекса качественных и количественных морфологических признаков согласно методике Т.А. Работнова (1992) и А.А. Уранова (1975). Онтогенетический спектр ЦП изучали методом учетных площадок (не менее 20 квадратов размером 1м2 для каждой ЦП). Онтогенетическую структуру ЦП анализировали по критерию «А-со» Л.А. Животовского (2001), а также с использованием некоторых демографических показателей: индексы замещения 1з и восстановления 1в (Жукова, 1987), индекс старения 1с (Глотов, 1998), эффективная плотность Ме (Животовский, 2001), коэффициент генеративности (g/g+v) (Трулевич, 1991). Оценка виталитетной структуры и жизненности ЦП дана с опорой на морфометрические параметры с использованием метода двумерной ранжировки особей на три класса виталитета (Злобин, 1989) и метода средне-взвешенного на основе 17 морфологических параметров с вычислением коэффициента IVC (Ишбирдин, Ишмуратова, 2004). Далее проводилась координация ценопопуляций по градиенту комплексного фактора благоприятности условий (установление экоклина). При изучении семенной продуктивности определяли число цветков и плодов в соцветии, потенциальную (ПСП) и реальную (РСП) семенную продуктивность согласно методикам Т.А. Работного (1992), И.В. Вайнагия (1974) и др.
Характеристика ЦП исследуемых видов
Таблица 1
|
№ ЦП |
Фитоценоз, высота над ур.м., м |
Экспозиция, крутизна склона,0 |
S ЦП, м2 |
N, тыс осб |
м, осб/м2 |
Антропогенное давление |
Уровень межвидовой конкуренции |
IVC |
|
Allium erubescens |
||||||||
|
5 |
Ф,450 |
ю, 30-40 |
15000 |
512,55 |
34,17 |
Слабое |
Низкий |
1,18 |
|
6 |
Ф, 100 |
в, 25-30 |
10200 |
381,68 |
37,42 |
Слабое |
Низкий |
1,16 |
|
И |
Ф,200 |
с-в, 30-40 |
20000 |
654,80 |
32,74 |
Умеренное |
Низкий |
1,15 |
|
9 |
ЩОЛ, 360 |
в, 45 |
12000 |
561,48 |
46,79 |
Слабое |
Средний |
1,06 |
|
2 |
ЩОЛ, 350 |
ю, 30 |
80000 |
4020,00 |
50,52 |
Слабое |
Средний |
1,04 |
|
1 |
ОпЛ, 350 |
- |
80000 |
780,00 |
9,75 |
Слабое |
Высокий |
1,03 |
|
3 |
ОЛ, 530 |
в, 30 |
90000 |
126,00 |
1,40 |
Слабое |
Высокий |
0,93 |
|
8 |
О Л, 120 |
- |
220000 |
1716,00 |
7,80 |
Умеренное |
Высокий |
0,90 |
|
10 |
ОЛ, 400 |
с-в, 30 |
50000 |
218,00 |
4,36 |
Слабое |
Высокий |
0,89 |
|
7 |
ОЛ, 20 |
- |
5680 |
29,31 |
5,16 |
Высокое |
Средний |
0,78 |
|
4 |
РД, 380 |
- |
6000 |
12,30 |
2,05 |
Сильное |
Низкий |
0,75 |
|
Allium rotundum |
||||||||
|
12 |
ПК, 500 |
3, - |
1500 |
1’025 |
30,68 |
Сильное |
Низкий |
1,47 |
|
И |
Ф, 790 |
ю-в, 35 |
1'728 |
53,60 |
31 |
Умеренное |
Низкий |
1,35 |
|
7 |
ОЛ, 400 |
ю-в, 20-30 |
5000000 |
5’402,76 |
45,23 |
Слабое |
Средний |
1,26 |
|
1 |
ОЛ, 410 |
- |
574,10 |
52,10 |
90,80 |
Умеренное |
Средний |
1,23 |
|
2 |
ОЛД00 |
- |
1'056 |
58,90 |
55,80 |
Умеренное |
Средний |
U9 |
|
14 |
ОЛ, 500 |
з, 30-40 |
6600 |
1’610,40 |
24,40 |
Сильное |
Средний |
1,13 |
|
8 |
ОЛ, 1220 |
ю, 20-25 |
750000 |
13’500 |
18 |
Умеренное |
Средний |
1,10 |
|
10 |
ПР Л, 840 |
ю-в, 35-40 |
20'160 |
1’500 |
76,85 |
Умеренное |
Средний |
1,03 |
|
9 |
СОЛ, 1400 |
ю, 0-10 |
3000000 |
30’000 |
6,60 |
Слабое |
Высокий |
0,94 |
|
13 |
О Л, 430 |
ю-в, 15 |
45000 |
173,25 |
3,85 |
Слабое |
Высокий |
0,91 |
|
3 |
ОЛ, 320 |
- |
768 |
69,043 |
89,90 |
Сильное |
Высокий |
0,88 |
|
4 |
ПР Л, 570 |
ю-в, 30 |
8'704 |
107,60 |
17,70 |
Сильное |
Средний |
0,79 |
|
5 |
СОЛ, 1150 |
с, 30 |
2хЮ6 |
93’000 |
46,50 |
Слабое |
Высокий |
0,73 |
|
6 |
СОЛ, 1250 |
ю, 30-40 |
600 |
8,86 |
8,09 |
Слабое |
Высокий |
0,72 |
Примечание. Ф - фриганоидное сообщество, ОпЛ - опушка леса, ОЛ - остепененный луг, ЩОЛ - щебнистый остепененный луг, СОЛ - субальпийский остепненый луг, РД -рудеральная растительность, ПК - песчаный карьер, ПР Л - приречный луг; S - площадь, N -численность, М - плотность особей ЦП; IVC - индекс виталитета ЦП, определяющий эколого-ценотический градиент ЦПшах—>ЦПтш.
В каждом пункте проводили сборы соцветий не менее 30 средневозрастных генеративных растений. Урожай оценивали по числу семян на 1м2, реализацию урожая (рождаемость) — по числу всходов на 1м2, реализацию семенной продуктивности - по отношению числа проростков к числу жизнеспособных семян (Левина, 1981; Жукова, 1995). Для оценки изменчивости анализировали 17 морфологических признаков 30 средневозрастных генеративных особей каждой ЦП: диаметр и высота луковиц, длина корневой системы, длина и ширина нижнего и верхнего листьев, высота побега и диаметр его основания, диаметр цветоноса, число цветков в соцветии, длина и ширина долей околоцветника, длина цветоножки, высота и диаметр соцветия. Для определения онтогенетических тактик вида (Злобин, 1989) средние значения коэффициента вариации признаков особей ценопопуляций CV,% располагали на экоклине. Определение типа онтогенетической стратегии проводилось на основе оценки характера изменения коэффициента детерминации R2m на экоклине (Ишбирдин, Ишмуратова, 2004). Названия жизненных форм видов даны в соответствии с классификацией В.А. Черемушкиной (2001). Стратегии выживания рассмотрены в рамках концепции Л.Г. Раменского (1935) и Дж. Грайма (Grime, 1979). Латинские названия таксонов даны по С.К. Черепанову (1995). Первичный материал обработан с использованием пакетов программ Statistica 6, MS Excel.
Результаты и обсуждение. Исследования показали, что наиболее типичными местами произрастания A. erubescens и A. rotundum являются луговые фитоценозы со средне и сильно задернованными почвами. Поэтому особый интерес для выявления адаптивных реакций видов представляют случаи развития растений на песчаных и песчано-каменистых склонах с природно и/или антропогенно эродированными почвами (ЦП5, ЦПб, ЦП 11 А. erubescens и ЦП11, ЦП12 A. rotundum). В подобных условиях широкое распространение в пределах местообитания получают растения слабопартикулирующей моноцентрической жизненной формы, что определяет переход ЦП обоих видов на преимущественно семенной, отчасти вегетативный способ самоподдержания (II тип по Л.А. Жуковой (1995). При этом наблюдается уменьшение доли ювенильных растений (дочерних рамет) в партикулах (рис. 1), особенно значительное для A. rotundum.
Благодаря снижению межвидовой и внутривидовой конкуренции, экономии организмом энергетических и пластических материалов, затрачиваемых на партикуляцию, относительно низкой изменчивости генеративных признаков растений, возрастанию в виталитетных спектрах доли вегетативно развитых особей класса жизненности «а» (рис. 2), обладающих, по определению (Злобин, 1989; и др.), большим репродуктивным потенциалом, в ЦП обоих видов наблюдаются максимальные показатели виталитета IVC и семенной продуктивности, размеров и всхожести семян (рис. 3, табл. 2).
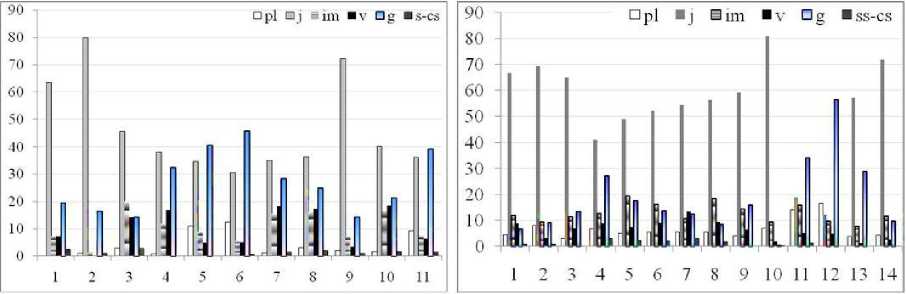
Рис. 1. Возрастные спектры ЦП A. erubescens (сверху) и A. rotundum (снизу): pl-cs — онтогенетические состояния; по оси абсцисс - номера ценопопуляций, по оси ординат - процент представленности возрастных групп в ЦП.
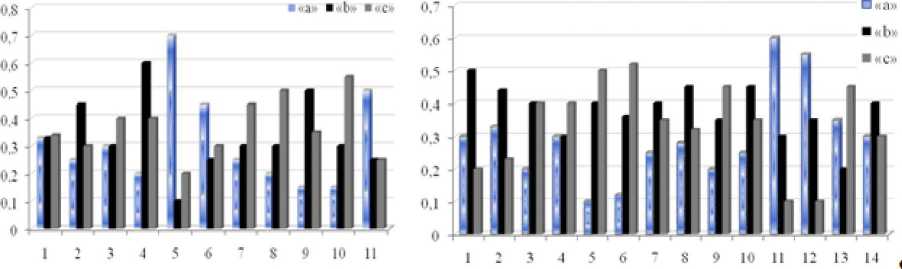
Рис. 2 . Виталитетные спектры ЦП A. erubescens (слева) и А. rotundum (справа), «а», «Ь», «с» - классы виталитета; по оси абсцисс -номера ценопопуляций, по оси ординат - доля каждого класса в спектре
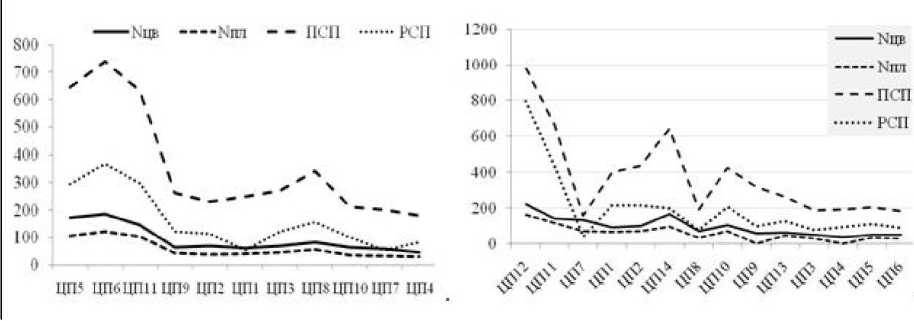
Рис. 3 . Динамика показателей семенной продуктивности А. erubescens (слева) и A. rotundum (справа) на эколого-ценотическом градиенте.
Мцв и Nnn - число цветков и плодов на побеге, ПСП и РСП - потенциальная и реальная продуктивность семян
Таблица 2
Качество свежесобранных семян и семенное возобновление ЦП
|
ЦП |
Всхожесть семян |
LceMXBceM |
Ng |
У |
рУ |
рСП |
|
Allium erubescens |
||||||
|
ЦП1 |
56 |
0,24x0,13 |
1,83 |
97,72 |
0 |
0 |
|
ЦП2 |
58 |
0,24x0,15 |
0,81 |
92,86 |
0,49 |
0,53 |
|
ЦПЗ |
45 |
0,23x0,13 |
0,19 |
23,03 |
0,04 |
0,17 |
|
ЦП4 |
32 |
0,25x0,13 |
0,65 |
54,39 |
0,01 |
0,02 |
|
ЦП5 |
78 |
0,26x0,14 |
13,60 |
4003,84 |
3,12 |
0,08 |
|
ЦП6 |
85 |
0,26x0,15 |
15,87 |
5862,85 |
4,53 |
0,08 |
|
ЦП? |
20 |
0,24x0,13 |
1,42 |
76,02 |
0,05 |
0,06 |
|
ЦП8 |
46 |
0,23x0,13 |
1,75 |
273,91 |
0,22 |
0,08 |
|
ЦП9 |
33 |
0,24x0,15 |
6,44 |
789,16 |
0,92 |
0,12 |
|
ЦП10 |
48 |
0,24x0,15 |
0,84 |
85,97 |
0,07 |
0,08 |
|
цпп |
66 |
0,26x0,15 |
12,48 |
3715,79 |
2,87 |
0,08 |
|
Allium rotundum |
||||||
|
ЦП1 |
39 |
0,26x0,13 |
6,11 |
1303,26 |
4,22 |
0,31 |
|
ЦП2 |
45 |
0,25x0,13 |
6,23 |
1323,08 |
5,00 |
0,38 |
|
ЦПЗ |
32 |
0,23x0,13 |
13,33 |
1035,34 |
25,51 |
2,46 |
|
ЦП4 |
30 |
0,23x0,12 |
3,86 |
365,66 |
1,71 |
0,47 |
|
ЦП5 |
24 |
0,25x0,14 |
8,11 |
881,56 |
5,44 |
0,62 |
|
ЦП6 |
26 |
0,23x0,12 |
4,40 |
390,02 |
0,80 |
0,20 |
|
ЦП? |
47 |
0,28x0,16 |
4,67 |
356,46 |
1,33 |
0,37 |
|
ЦП8 |
36 |
0,23x0,13 |
3,50 |
267,15 |
1,50 |
0,56 |
|
ЦП9 |
36 |
0,24x0,14 |
4,50 |
444,01 |
5,00 |
1,13 |
|
ЦП10 |
34 |
0,30x0,15 |
1,54 |
318,47 |
0,09 |
0,03 |
|
ЦПП |
52 |
0,31x0,16 |
6,91 |
3040,40 |
8,21 |
0,27 |
|
цпп |
63 |
0,31x0,16 |
5,40 |
4288,40 |
8,60 |
0,20 |
|
ЦП13 |
34 |
0,19x0,10 |
2,30 |
287,96 |
0,30 |
1,04 |
|
ЦП14 |
37 |
0,25x0,14 |
3,60 |
7200 |
0,40 |
0,005 |
Примечание. Всхожесть семян, % - число проросших семян от общего числа заложенных в опыт; ЬсемхВсем, мм - длина и ширина семени; Ng, осб/м2 - число генеративных особей на м2; У, шт/м2 - урожай (число семян на м2); рУ, осб/м2 -реализация урожая (число проростков па м2); рСП, % - реализация семенной продуктивности (процентное отношение реализации урожая к урожаю).
Соответственно, несмотря на относительно низкую плотность генеративных особей, постоянную угрозу смыва семян дождевыми водами и иссушения повышаются показатели семенного возобновления ЦП, однако общая эффективность самоподдержания (1в и 1з - индексы восстановления и замещения), а также плотность ЦП падают (табл. 3): основную роль в возобновлении A. erubescens и А. rotundum в природе играет вегетативное размножение.
Таким образом, захват и удержание территории, поддержание численности, средовлияния ЦП A. erubescens и A. rotundum и их участия в жизнедеятельности фитоценоза в пределах нарушенных, но благоприятных для произрастания местообитаний достигается в основном за счет интенсификации роста и семенного размножения особей. Подобная реакция свидетельствует о наличии в жизненной стратегии видов эксплерентной составляющей.
Таблица 3 Демографические показатели ценопопуляций
|
№ ЦП |
A |
co |
Me |
IB |
13 |
Ic |
g/(v+g) |
Тип ЦП |
|
Allium erubescens |
||||||||
|
1 |
0,15 |
0,27 |
2,63 |
4,00 |
3,55 |
0,03 |
0,64 |
Молодая |
|
2 |
0,09 |
0,21 |
10,61 |
5,03 |
4,74 |
0,02 |
0,98 |
Молодая |
|
3 |
0,13 |
0,24 |
0,34 |
5,80 |
4,83 |
0,04 |
0,50 |
Молодая |
|
4 |
0,24 |
0,38 |
0,78 |
0,89 |
0,89 |
- |
0,52 |
Молодая |
|
5 |
0,26 |
0,45 |
15,38 |
1,12 |
1,12 |
- |
0,89 |
Молодая |
|
6 |
0,30 |
0,47 |
17,59 |
0,42 |
0,26 |
0,01 |
0,92 |
Молодая |
|
7 |
0,23 |
0,34 |
1,75 |
1,25 |
1,06 |
0,02 |
0,40 |
Молодая |
|
8 |
0,20 |
0,32 |
2,50 |
3,12 |
2,26 |
0,03 |
0,36 |
Молодая |
|
9 |
0,11 |
0,24 |
11,23 |
5,48 |
4,76 |
0,02 |
0,68 |
Молодая |
|
10 |
0,17 |
0,29 |
1,26 |
3,45 |
2,86 |
0,02 |
0,35 |
Молодая |
|
11 |
0,24 |
0,43 |
14,08 |
0,72 |
0,43 |
0,02 |
0,75 |
Молодая |
|
Allium rotundum |
||||||||
|
1 |
0,15 |
0,18 |
16,43 |
13,69 |
12,14 |
0,009 |
0,43 |
Молодая |
|
2 |
0,07 |
0,18 |
10,43 |
10,93 |
9,45 |
0,007 |
0,70 |
Молодая |
|
3 |
0,08 |
0,21 |
18,88 |
6,96 |
6,96 |
- |
0,64 |
Молодая |
|
4 |
0,12 |
0,25 |
4,74 |
2,96 |
2,32 |
0,03 |
0,79 |
Молодая |
|
5 |
0,12 |
0,22 |
10,23 |
5,16 |
4,23 |
0,02 |
0,65 |
Молодая |
|
6 |
0,08 |
0,19 |
1,54 |
6,18 |
4,30 |
0,02 |
0,49 |
Молодая |
|
7 |
0,11 |
0,24 |
10,85 |
8,70 |
6,37 |
0,03 |
0,48 |
Молодая |
|
8 |
0,04 |
0,13 |
2,34 |
11,14 |
7,43 |
0,02 |
0,50 |
Молодая |
|
9 |
0,07 |
0,17 |
1,12 |
5,47 |
5,57 |
- |
0,74 |
Молодая |
|
10 |
0,03 |
0,09 |
6,92 |
189,18 |
134,01 |
0,002 |
0,22 |
Молодая |
|
11 |
0,21 |
0,31 |
9,61 |
2,44 |
1,64 |
0,015 |
0,84 |
Молодая |
|
12 |
0,27 |
0,58 |
17,79 |
1,71 |
1,71 |
- |
0,92 |
Молодая |
|
13 |
0,18 |
0,30 |
1,15 |
2,94 |
2,94 |
- |
0,81 |
Молодая |
|
14 |
0,06 |
0,15 |
3,66 |
10,08 |
10,08 |
- |
0,72 |
Молодая |
Примечание. Примечание: А и со - индексы возрастности и эффективности, Ме, осб/м2 - эффективная плотность; 1в, 1з, 1с - индексы восстановления, замещения, старения; g/(v+g) - коэффициент генеративности.
При произрастании на более-менее задернованных почвах оба вида сохраняют типичную партикулирующую неявнополицентрическую короткостолонообразующую жизненную форму и характерный IV тип самоподдержания (вегетативное, отчасти семенное), хотя с существенным усилением межвидовой конкуренции в фитоценозе происходит снижение порядка ветвления в партикулах и, соответственно, уменьшение роли вегетативного размножения в возобновлении ЦП.
Активизация партикуляции при произрастании в фитоценозах со средне задернованными почвами (ЦП2, ЦП9 Л. erubescens и ЦП1, ЦП2, ЦП7, ЦП8, ЦП10, ЦП14 Л. rotundum), возможная только при значительном увеличении энергетических и пластических затрат на образование дочерних луковиц, с одной стороны, сопровождается возрастанием в виталитетных спектрах доли наиболее вегетативно подвижных растений класса виталитета «Ь», с другой стороны -снижением виталитета ЦП, семенной продуктивности, качества семян (размеров, всхожести), а во многих случаях - и параметров семенного возобновления. В то же время возрастание долевого участия в возрастных спектрах ювенильных особей вегетативного происхождения определяет значительное увеличение индексов восстановления и замещения, плотности ЦП.
Таким образом, при произрастании на почвах со средним уровнем задернения почвы для Л. erubescens и Л. rotundum характерно повышение эффективности вегетативного размножения, специфика которого (образование одним материнским растением до 100 (Л. erubescens), 150 (Л. rotundum) и более коллатеральных луковичек) позволяет значительно увеличить плотность особей в ЦП. Наряду с проявлением защитной компоненты в онтогенетической стратегии видов (рис. 4) подобная реакция подтверждает выраженность R-составляющей их эколого-ценотической стратегии.
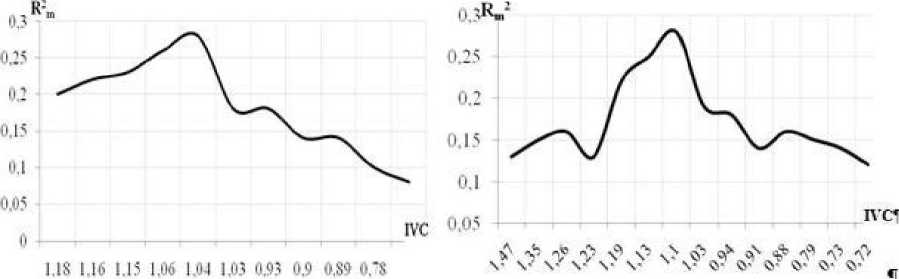
Рис. 4 . Тренд онтогенетической стратегии Л. erubescens (слева) и
A. rotundum (справа). По оси абсцисс - IVC (индекс виталитета ЦП), по оси ординат - R2m (коэффициент детерминации)
Падение интенсивности партикуляции при значительном усилении межвидовой конкуренции (ЦП1, ЦПЗ, ЦП8, ЦП 10 Л. erubescens и ЦПЗ, ЦП5, ЦПб, ЦП9, ЦП13 Л. rotundum) на фоне снижения семенной продуктивности и качества семян негативно сказывается на эффективности самоподдержания ЦП. В то же время, подавленные рост и размножение растений, левосторонность виталитетных спектров, стабилизация вегетативных признаков, морфологическая дезинтеграция особей способствуют экономии и оптимизации перераспределения энергетических и пластических ресурсов на поддержание минимально необходимого для существования ЦП уровня ростовых и репродуктивных процессов, численности, являясь механизмами устойчивости вида в крайне неблагоприятных условиях. Обеспечение базового уровня ростовых и репродуктивных процессов, необходимого для существования и сохранения ЦП своего места в фитоценозе с высоким уровнем межвидовой конкуренции, наряду с подчиненным положением в фитоценозах и способностью избегать фитоценотической конкуренции через формирование листьев летне-осенней генерации являются явными признаками проявления фитоценотической патиентности A. erubescens иА. rotundum.
Подобное снижение активности ростовых и репродуктивных процессов характерно также для ЦП4, ЦП7 A. erubescens и ЦП4 A. rotundum, произрастающих на средне задернованных каменистых почвах, но в условиях сильного антропогенного давления в форме вытаптывания и перевыпаса скота.
Выводы. В итоге, исследование формирования жизненной стратегии обоих видов в неоднородных условиях среды выявило наличие рудерально-патиентной (эксплерентно-стресс-толерантной, RS) стратегии жизни, обеспечивающей им возможность быстро захватывать эродированные и более-менее свободные участки территории, а также длительно выживать под прессом у виолентов (фитоценотическая патиентность) и в неблагоприятных экологических условиях антропогенного пресса (экологическая патиентность). В последнем случае к основным признакам экологической стресс-толерантности можно отнести также типичное обитание видов в засушливых местах с высокой степенью инсоляции, жизненную форму (геофиты с аллокацией запасных питательных веществ в луковице), эндогенный покой семян, а для A. rotundum - суккулентность надземных органов растений в среднегорье (1200-1400 м над уровнем моря).
В схожих условиях среды устойчивость ЦП A. erubescens и А. rotundum достигается за счет аналогичных организменных и популяционно-онтогенетических механизмов, что наряду с данными морфологии и фитоценологии свидетельствует о тесном родстве двух видов. Следовательно, особенности формирования жизненной стратегии не могут быть использованы как диагностические таксономические признаки при разграничении данных видов.
Выражаем искреннюю благодарность профессору МГУ Владимиру Гертрудовичу Онипченко и сотрудникам Горного ботанического сада ДНЦ РАН в лице директора Загирбека Магомедовича Асадулаева и и.о. зав. лаб. фитохимии и медицинской ботаники Абдулахида Магомедовича Мусаева за помогць, оказанную в организации полевых исследований и сбора материала на территории Карачаево- Черкесской Республики и Республики Дагестан.
Чадаева В.А. Близкородственные виды Allium erubescens С. Koch и Allium rotundum (Allium, Alliaceae) с точки зрения формирования жизненной стратегии / В.А. Чадаева // Вести. ТвГУ. Сер.: Биология и экология. 2016. № 4. С. 206-218.
Список литературы Близкородственные виды Allium erubescens C. Koch. и Allium rotundum L. (Allium, Alliaceae) с точки зрения формирования жизненной стратегии
- Вайнагий И.В. 1974. К методике изучения семенной продуктивности растений//Бот. журн. Т. 59. № 6. С. 826-831.
- Введенский А.И. 1935. Род Аlliит L.//Флора СССР. Т. 4.: сб. труд. М.-Л.: Изд. АН СССР С. 112-280.
- Галушко А.И. 1978. Флора Северного Кавказа. Определитель. Т.1. Ростов-на-Дону: Изд. Ростовского университета. 320 с.
- Глотов Н.В. 1998. Об оценке параметров возрастной структуры популяций растений//Жизнь популяций в гетерогенной среде: материалы Всероссийского популяц. семинара. Йошкар-Ола: Периодика Марий Эл. С. 146-149.
- Гроссгейм A.A. 1940. Флора Кавказа. Т.2. Баку: Изд. АзФАН. С. 110-141.
- Животовский Л.А. 2001. Онтогенетические состояния, эффективная плотность и классификация популяций//Экология. № 1. С. 3-7.
- Жукова Л.А. 1987. Динамика ценопопуляций луговых растений в естественных фитоценозах//Динамика ценопопуляций травянистых растений: сб. тр. Киев: Наукова думка. С. 9-19.
- Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола: РИИК «Ланар». 224 с.
- Зернов А.С. 2010. Растения российского Западного Кавказа. Полевой атлас. М.: Т-во научн. изд. КМК. 449 с.
- Злобин Ю.А. 1989. Теория и практика оценки виталитетного состава ценопопуляций растений//Бот. журн. Т. 74. № 6. С. 769-781.
- Ишбирдин А.Р., Ишмуратова М.М. 2004. Адаптивный морфогенез и эколого-ценотические стратегии выживания травянистых растений//Методы популяционной биологии: сб. материалов VII Всерос. популяц. семинара. Сыктывкар. С. 113-120.
- Косенко И.С. 1970. Определитель высших растений Северо-Западного Кавказа и Предкавказья. М.: Изд. Колос. 613 с.
- Кудряшова Г.Л. 1990. Кариосистематические заметки о кавказских видах рода Allium (Alliaceae).//Бот. журн. Т. 75. № 6. С. 829-832.
- Кудряшова Г.Л. 1992. Конспект видов рода Allium (Alliaceae) Кавказа//Бот. журн. Т. 77. № 4. С. 86-88.
- Левина Р.Е. 1981. Репродуктивная биология семенных растений. М.: Наука. 96 с.
- Немирова E.С., Троцкая И.В. 2008. К экологии видов рода Allium L. флоры Предкавказья//Вестник МГОУ. Серия «Естественные науки». №3. С. 46-51.
- Погосян А.И. 1992. Луковые Кавказа (систематика, карпология, экология, хорология, эволюция): автореф. дис. … канд. биол. наук., Ереван: Изд. Ин-та ботаники. 66 с.
- Работнов Т.А. 1992. Фитоценология: Уч. пос. 3-е изд., перераб. и доп. М.: Изд. МГУ. 352 с.
- Раменский Л.Г. 1935. О принципиальных установках, основных понятиях и терминах производственной типологии земель//Сов. ботан. № 4. С. 25-42.
- Серегин А.П. 2005. Ревизия диких луков (Allium, Alliaceae) флоры Восточной Европы//Изучение флоры Восточной Европы: достижения и перспективы: тезисы докл. международ. конф. СПб. С. 78-79.
- Серегин А.П. 2007. Род Allium L (Alliaceae) во флоре Восточной Европы: автореф. дис. … канд. биол. наук. Москва. 241 с.
- Танфильев В.Г., Кононов В.Н. 1987. Каталог дикорастущих растений Ставропольского края. Ставрополь: СНИИСХ. 116 с.
- Тахтаджян А.Л. 1966. Систематика и филогения цветковых растений. М.-Л.: Наука. 611 с.
- Трулевич Н.В. 1991. Эколого-фитоценотические основы интродукции растений. М.: Наука. 216 с.
- Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов//Научные доклады ВШ. Биол. наук. № 2. С. 7-34.
- Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья. 990 с.
- Чешмеджиев И.В. 1979. Кариологично изследоване на A. rotundum L.//Висш. селкост. инст. «В. Коларов» Научни трудове. Т. 24. Кн. 1, С. 165-168.
- Шхагапсоев С.Х., Чадаева В.А. 2015. Механизмы устойчивости видов растений на примере Allium albidum Fisch. ex Bieb. Центрального Кавказа»//Экология. №2. С. 103-109.
- Mathew B. 1996. A review of Allium section Allium. Kew: Royal Botanic Gardens. 176 p.
- Grime J.P. 1979. Plant strategies and vegetation processes. Chichester-New York-Brisbane-Toronto: John Wiley & Sons. 222 р.
