Ca2 + -зависимая аденилатциклаза растительных клеток в бактериальном патогенезе

Автор: Романенко А.С., Филинова Н.В., Ломовацкая Л.А.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 2 т.15, 2019 года.
Бесплатный доступ
Это первое исследование, в котором обнаружено влияние ионов кальция на активность трансмембранной аденилатциклазы (tmAC), ключевого фермента сигнальной системы аденилатциклазы, в нормальных условиях и после кратковременного воздействия экзополисахаридов (EPS) бактериальный кольцевой гниль, возбудитель Clavibacter michiganensis ssp. сепедоник (см). После обработки корней растений ЭПС Cms реакция на Ca2 + изменилась: активность tmAC растений устойчивого сорта значительно увеличилась, тогда как в клетках восприимчивого сорта она осталась неизменной.
Короткий адрес: https://sciup.org/143166907
IDR: 143166907
Ca2+-dependent adenylate cyclase of plant cells in bacterial pathogenesis
This is the first study to detect the effect of calcium ions on the activity of transmembrane adenylate cyclase (tmAC), the key enzyme of the adenylate cyclase signaling system, under normal conditions and after a short-term exposure to exopolysaccharides (EPS) of the bacterial ring rot pathogen Clavibacter michiganensis ssp. sepedonicus (Cms). After the treatment of the roots of plants with the Cms EPS, the response to Ca2+ changed: the activity of the tmAC of plants of the resistant cultivar significantly increased, whereas in the cells of the susceptible cultivar it remained unchanged.
Текст научной статьи Ca2 + -зависимая аденилатциклаза растительных клеток в бактериальном патогенезе
В настоящее время ионы кальция признаны универсальными посредниками в клеточных реакциях организмов растений и животных. Именно ионы кальция в цитозоле служат связующим звеном для многих сигнальных путей, обеспечивая формирование сигнальной сети растительной клетки (Guo et al. , 2018). Передача стрессовых сигналов с участием Ca2+ происходит благодаря его способности дифференцированно взаимодействовать с клеточными белками, в том числе с ферментами, которые являются ключевыми звеньями в других сигнальных системах растений (Hepler, 2005).
На клетках животных показано, что аденилатциклазная и кальциевая сигнальные системы активно реагируют на стрессоры разной природы (Cao et al. , 2017), при этом тесно взаимодействуя между собой. У растений исследования по взаимному влиянию сигнальных систем менее многочисленны, чем таковые у животных. Пока в клетках растений выявлены только неселективные, регулируемые нуклеотидами кальциевые каналы, активация которых происходит при присоединении цАМФ (Chin et al. , 2009). У животных давно выявлены изоформы аденилатциклазы, фермента, синтезирующего цАМФ, отличающиеся по чувствительности к ионам кальция (Halls, Cooper, 2011). Для растений такие сведения единичны (Chatukuta et al. , 2018), что значительно осложняет понимание путей трансдукции сигналов, происходящих в растительных клетках на ранних этапах воздействия стрессоров. Цель настоящей работы – исследовать изменение чувствительности к экзогенным ионам Ca2+ трансмембранной аденилатциклазы (тмАЦ) из клеток растений двух сортов картофеля, контрастных по устойчивости к бактериальному возбудителю кольцевой гнили Clavibacter michiganensis ssp. s epedonicus ( Cms ), при действии экзополисахаридов (ЭПС) Cms.
MATERIALS AND METHODS
В работе использовали растения картофеля ( Solanum tuberosum L . ) сорта Луговской
(резистентный к Cms), сорта Лукьяновский (чувствительный к Cms) и ЭПС Cms вирулентного, мукоидного штамма 5369. Экзополисахариды Cms добавляли в жидкую среду роста растений и инкубировали в течение 1 мин. Пробоподготовку для выделения тмАЦ проводили по стандартной схеме, описанной в (Ломоватская и др., 2015). Кальций добавляли к выделенному ферменту в виде CaCl2 (ч.д.а., “Реахим”, Россия) в следующих конечных концентрациях: 500 нМ, 1мкМ, 500 мкМ, 1 и 10 мМ, рассчитанных по содержанию Са2+. Об активности фермента судили по концентрации цАМФ, которую определяли методом иммуноферментного анализа. Активность выражали в мг белка/мин. Белок в пробе определяли по методу Бредфорда. Контролем служили образцы без добавления кальция. Эксперименты проводили в двукратной биологической и четырёхкратной аналитической повторностях. Результаты обрабатывали статистически с помощью программы SigmaPlot v. 12.3 (“SYSTAT Software”, США). По литературным данным свободные ионы Са2+ в растительных клетках присутствуют в микромолярных концентрациях. Эти концентрации могут резко и кратковременно повышаться (до миллимолей в течение нескольких секунд) под влиянием стрессоров разной природы (White, Broadley, 2003).
Поэтому в наших экспериментах мы применяли кальций в микро- и миллимолярных концентрациях.
RESULTS
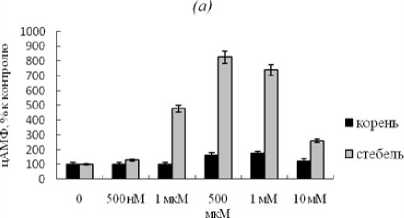
В корнях растений чувствительного сорта в варианте без ЭПС добавление в среду роста растений кальция при всех концентрациях не оказывало влияния на активность тмАЦ, тогда как после добавлении ЭПС кальций при самой высокой концентрации (10 мМ) ингибировал активность фермента на 30% (рис. 1 а , б ). В стеблях тех же растений в варианте без ЭПС 1 мкМ, 500 мкМ и 1 мМ кальция существенно стимулировали активность тмАЦ (рис. 1 а ). После инкубации с ЭПС при высоких концентрациях этого катиона (1 и 10 мМ) наблюдалось значительное ингибирование, особенно в стеблях, активности тмАЦ (рис. 1 б ).
У картофеля, резистентного к Сms сорта, в корнях активность тмАЦ после добавления кальция возросла как в варианте с ЭПС, так и без него. Чем больше была концентрация кальция, тем выше была активность фермента (рис. 2а, б). В варианте с ЭПС активирующий эффект всех концентраций Са2+ был более выраженным. В клетках стеблей тех же растений в варианте без ЭПС активность тмАЦ не изменилась в присутствии всех концентраций кальция, но после добавлении ЭПС значительно возросла, особенно при миллимолярных концентрациях катиона (рис. 2б).
Коицент рати Саг*
в 5
( б )
llllll
■ корень
□ стебель
500 нМ 1 мкМ 500 мкМ 1 мМ 10 мМ
Концентрация Са 2+
Figure-1. Изменение активности тмАЦ из клеток растений картофеля восприимчивого сорта под влиянием разных концентраций ионов кальция. Здесь и на рис. 2 (а) – контрольный вариант (без добавления ЭПС Cms), (б) – опытный вариант (с добавлением ЭПС Cms). M ± m.
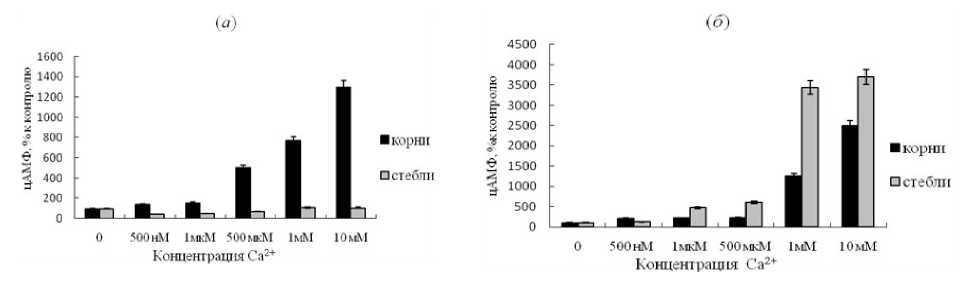
Figure-2. Изменение активности тмАЦ из клеток растений картофеля устойчивого сорта под влиянием различных концентраций ионов кальция.
DISCUSSION
Модуляция активности тмАЦ ионами кальция вероятно, свидетельствует о наличии нескольких изоформ этого фермента, отличающихся по чувствительности к данному иону. Возможно, у растений обоих сортов спектр изоформ тмАЦ различен. На основании полученных данных можно предположить, что эта особенность является одним из механизмов разной устойчивости растений данных сортов к Cms и обусловлена развитием/блокировкой системного ответа растений картофеля обоих сортов на биотический стрессор. Ранее (Ломоватская и др., 2004) мы также установили зависимость системного ответа на биотический стрессор аденилатциклазной сигнальной системы растений картофеля от их сортовой принадлежности.
Опираясь на ранее полученные результаты (Романенко, Ломоватская, 2017) о наличии в структуре плазмалеммы и мембран клеточных органелл растений картофеля этих сортов цАМФ-зависимых Са2+-каналов, локализация которых количественно изменялась в ответ на воздействие
ЭПС Cms , можно предположить существование возможного механизма взаимовлияния аденилатциклазной и кальциевой сигнальных систем при биотическом стрессе. В цитируемой работе мы показали, что в клетках корней растений тех же сортов картофеля ЭПС Cms интенсивно модулировали активность тмАЦ. По крайней мере, в клеточной стенке и плазмалемме клеток растений картофеля присутствуют рецепторы к ЭПС Cms , имеющие количественные и качественные сортовые различия. У растений восприимчивого сорта эти рецепторы обладали сродством к супрессорным компонентам ЭПС, у растений устойчивого сорта – к элиситорным (Романенко и др , 1998). Очевидно, что системная реакция индуцируется в корнях, непосредственно контактирующих со стрессовым фактором. При этом под воздействием стрессора в плазмалемме клеток корней растений возникает электрохимический потенциал, системно распространяющийся до верхушки стебля (Новикова и др , 2017), а активность тмАЦ меняется под его воздействием (Соореr et al., 1998). Модуляция активности тмАЦ может приводить к локальному изменению концентрации цАМФ, что способствует активации потенциалзависимых Са2+-каналов, повышая кратковременно и локально концентрацию ионов кальция в составе плазмалеммы за счёт их выхода из свободного пространства клеточной стенки (Новикова и др , 2017). Дополнительный вклад в супрессию развития системного сигнала в клетках растений чувствительного сорта вносит ингибирование тмАЦ высокими концентрациями кальция. Очевидно, что для растений каждого сорта картофеля существует тонкая внутренняя регуляция этих процессов, зависящая от первичных триггеров сигналов, в частности от рецепторов, а также от активности тмАЦ.
ACKNOWLEDGMENT
Работа выполнена на оборудовании ЦКП “Биоаналитика” с использованием коллекций ЦКП “Биоресурсный центр” Сибирского института физиологии и биохимии растений СО РАН (г. Иркутск).
Список литературы Ca2 + -зависимая аденилатциклаза растительных клеток в бактериальном патогенезе
- Cao X.-Q., Juang Z.-H., Yi Y.-Y., Yang Y., Ke L.-P., Pei Z.-M., Zhu S. (2017) Biotic and abiotic stresses activate different Ca2+ permeable channels in Arabidopsis. Frontiers in Plant Sci., 8, Article 83 DOI: 10.3389/fpls.2017.00083
- Chatukuta P., Dikobe T.B., Kawadza D.T., Sehlabane K.S., Takundwa M.M., Wong A., Gehring C., Ruzvidzo O. (2018) Biomolecules, 8(15) DOI: 10.3390/biom8020015
- Chin K., Moeder W., Yoshioka K. (2009) Biological roles of cyclic ucleotide-gated ion channels in plants: What we know and don’t know about this 20 member ion channel family. Botany, 87, 668-677
- Cоореr D.M.F., Shell M.J., Thorn P., Irvine R.F. (1998) Regulation of adenylyl cyclase by membrane potential. J. Biol. Chem., 273(2), 27703-27707
- Guo J., Islam M.A., Lin H., Ji C., DuanY., Peng L., Zeng Q., Day B., Kang Z., Guo J. (2018) Genome-wide identification of cyclic nucleotide-gated ion channel gene family in wheat and functional analyses. Frontiers in Plant Sci., 9, Article 18 DOI: 10.3389/fpls.2018.00018
- Halls M.L., Cooper D.M.F. (2011) Regulation by Ca2+ -signaling pathways of adenylyl cyclases. Bootman M.D., Berridge M.J., Putney J.W., Roderick H.L. (eds). Cold Spring Harbor Perspectives in Biology, 3, Cold Spring Harbor Laboratory Prerss, Cambridge, UK, 1-22 a004143
- Hepler P.K. (2005) Calcium: a central regulator of plant growth and development. Plant Cell, 17, 2142-2155
- White P.J., Broadley M.R. (2003) Calcium in plants. Annals Bot., 92, 487-511
- Ломоватская Л.А, Романенко А.С., Рыкун О.В. (2015) Трансмембранная аденилатциклаза контролирует факторы вирулентности фитопатогена Pseudomonas siringae и мутуалиста Rhizobium leguminosarum. Микробиология, 84(4), 404-410
- Ломоватская Л.А., Романенко А.С., Криволапова Н.В., Копытчук В.Н., Саляев Р.К. (2004) Возможная роль эндогенного цАМФ картофеля в развитии системного сигнала при патонегезе кольцевой гнили. ДАН, 394(5), 715-117
- Новикова Е.М., Воденеева В.А., Сухов В.С. (2017) Разработка математической модели потенциала действия высших растений, учитывающей роль вакуоли в генерации электрического сигнала. Биологические мембраны, 34(2), 109-125
- Романенко А.С., Ломоватская Л.А. (2017) Влияние экзополисахаридов бактериального возбудителя кольцевой гнили на субклеточную локализацию регулируемых циклическими нуклеотидами ионных каналов (CGNC) в клетках корней картофеля. Биологические мембраны, 34(2), 1-8
- Романенко А.С., Рымарева Е.В., Шафикова Т.Н. (1998) Компоненты оболочек клеток картофеля, обладающие сродством к токсину возбудителя кольцевой гнили. ДАН, 358(2), 277-279
