Chamaecyparis belgica - хвойное из палеоценовой Волжско-Южноуральской провинции Гелинденской палеофлористической области

Автор: Викулин С.В., Моров В.П.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 1 т.10, 2016 года.
Бесплатный доступ
Исследованы образцы с отпечатками хвойного - Chamaecyparis belgica Saporta et Marion, 1873 (элемент ‘гелинденской флоры’ Бельгии) из палеоценовых отложений Поволжья (Вольск, Шиханы, верховья реки Свияги, Ульяновско-Саратовский прогиб) и Южного Урала (Романколь) по новым сборам из Поволжья и по классическим палеоботаническим коллекциям начала и середины XX в., хранящимся в БИН РАН.
Палеоцен северный перитетис cupressaceae chamaecyparis побеги
Короткий адрес: https://sciup.org/148314562
IDR: 148314562 | УДК: 561.47:
Текст научной статьи Chamaecyparis belgica - хвойное из палеоценовой Волжско-Южноуральской провинции Гелинденской палеофлористической области
В России местонахождения палеоценовых “тетисовых флор” гелинденского экологического типа1 известны из верхнесызранских, нижнесаратовских и верхнесаратовских (= камышинских) отложений Поволжья [Вольск, Шиханы, верховья реки Свияги в пределах Ульяновско-Саратовского прогиба] и на правобережье нижней Волги [гора Уши близ Ка-мышина2] (Леонов, 1936, 1956, 1961, 1967; Тахтаджян, 1966; Геология СССР, 1969, 1970; Легенда…, 1998, 1999; Александрова, 2013; Ахметьев, Запорожец, 2014), рис. 1. Указанные свиты палеоцена сопоставляются с концом зеландия – танетом.
На Южном Урале и в Западном Казахстане также имеются местонахождения флор ге-линденского типа: Романколь, Тыкбутак (рис. 2 A, Б). Доминирующими компонентами ге-линденско-камышинско-южноуральской палеоценовой флоры являются: 1) вечнозеленые буковые, близкий Nothofagusвымерший род Ushia,древний каштанодуб Dryophyllum; 2) загадочная Dewalquea, возможно относящаяся к сем. Araliaceae; 3) разнообразные лавровые, а также пальмы и другие растения тропического и субтропического родства3.
По своему составу гелинденско-камышинско-романкольские флоры довольно близки, но примечательна особая крупноли-стность многих видов флоры Романкульсая на Южном Урале. Еще А.П. Павлов (1896, 1897), анализируя флору камышинского горизонта, пришел к выводу, что к ней наиболее близка флора бельгийского типа. И, тем не менее, по его мнению, данные флористические комплексы не могут рассматриваться как одновозрастные, так как камышинская флора принадлежит более высокому стратиграфическому горизонту, близкому к серии лигнитов Франции или к более древним слоям Англии: outcrops of the Reading Formation and lower part of the London Clay Formation on the Isle of Wight
|
23 х 4 |
су X X X н н ад о о 1 |
х СУ Е S ад го |
СУ ад н О О го |
Я Я СУ |
cy DS |
W 3 § |
ад о о № О ■ri |
||||||
|
S В я s E 8 я a g Я я д к s g к a | a щ |
|||||||||||||
|
ЕХИНЭ КЕМЭЯОХ -вс1вээнх<1?д |
В1И8Э квхэнвскнэ |
||||||||||||
|
5S x rid Q - О ^ X О СУ §2 5 ~ co Ы |
ё Ж у 3 и 5 и а 'В Й 8 ” 2 g£ о го с |
Ж Ё X ° и 2 |
ж Ё X си X g |
я Ё го 2 |
§ 1 II к 2 |
я ™ я 5 о ™ g 5 | * ri I S ° |
Я? P s ад Он m x w U |
А § ад й о ад$ н и х н s 5 s 5 S о. § ад з ад Q |
)S |
||||
|
ъмияо КЕЯЭН1ЧЙИ<1в^ |
втиао кeлoи9uodц |
В1ИЯЭ квяонишнн^ |
oXdK HHMOHEdcHQ |
||||||||||
|
>s s rid -Q 2 00 x n ri < |
о и и G S § к g ж $ го X )Q | и * о. I а С |
X U а я я ^33 Си |
у о w X х X S 3 о X — Q |
3 >я '_> ГО 1 <И О У я ® § ГО а |
я 9 я X § о u S ° Й СА ад о |
5 5 ГО S СТО щ |
(D , 6 x 2 к g S 2 2 й о К 9 й о |
ад pj 5 5^5 К |
о • |
о о о 5 |
|||
|
элак иияэннйи(1б]^ |
oXdb hi |
im; |
)8 |
OiadEJ ИИ |
эЛйи xoHEdcH^ |
||||||||
|
PQ О G oo ri ^ < |
ё ж 5 «ХЮ 3 g « § ^ и ■го-------- 5^ |
ж а в ад Е 9 я 9 я ^ Е 2 X го |
th m t |
| ?/ islslggs| V О § g = / х " & 1 ” ^ |
в ад и 3 о ад х ад ° ад ° К |
ад ад У Е |
§ 5 |
||||||
|
оЛс!к иимоао±вс1вэ ку |
вхияэ MOHedm0 |
||||||||||||
|
1 >x О X 23 5 g 2 8 v у X X О ^5 |
< V < X У ^ ■ X • X < >С ' X . х: х < X < К |
■V. |
<’■;.'< |
Ws 0# |
11^ |
4<4< , х^ с с : х(с(с< УхС с С с ( У// |
|||||||
|
c- ГО ri |
и S X Cl |
tD X X s и з С Го я |
Q Я Я Я S з g s Я 3 ГО й я |
W О X DX Н |
es X X X 3 “ |
о X о X X Я О V О о |
и я у х 5 я х ад о |
D S 8 § :Q О Оо W |
су S ^ О о о 2 |
||||
|
У1ИЯЭ KBMOHHliHd^ |
X |
В1ИЯ0 кемэнвсклчэ |
|||||||||||
|
Н м го |
ИИНЖИЦ |
HHHxdag |
HHHS9d3 |
S S |
|||||||||
|
H9h0£ |
НЭПООКЕЦ |
||||||||||||

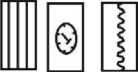

Рис. 1. Сводный стратиграфический разрез палеоценовых и раннеэоценовых отложений Поволжья на основе различных стратиграфических схем (по: Леонов, 1936, 1961, 1967 и Александрова, 2013)
Combined stratigraphic profile of Paleocene and Early Eocene sediments of the Volga region on the basis of different stratigraphic schemes (after: Leonov, 1936, 1961, 1967 and Aleksandrova, 2013)
(Геология СССР…, 1969; Ali, 1994). Зона равномерно влажного на протяжении годового цикла субтропического и паратропиче-ского климата Северного полушария в палеоцене занимала средние широты современного Северного полушария (Mai, 1991, 1995). Осуществлялся свободный океанический и водный перенос тепла из тропического Тети- са посредством открытого океанического обрамления северного Перитетиса в Арктический бассейн. В так называемый Перитетис входили, в том числе и Тургайский пролив, и Западно-Сибирский внутренний бассейн (Ахметьев, Беньямовский, 2006; Беньямов-ский, 2007). Паратропические раннепалеогеновые флоры Северного Перитетиса в Ниж-
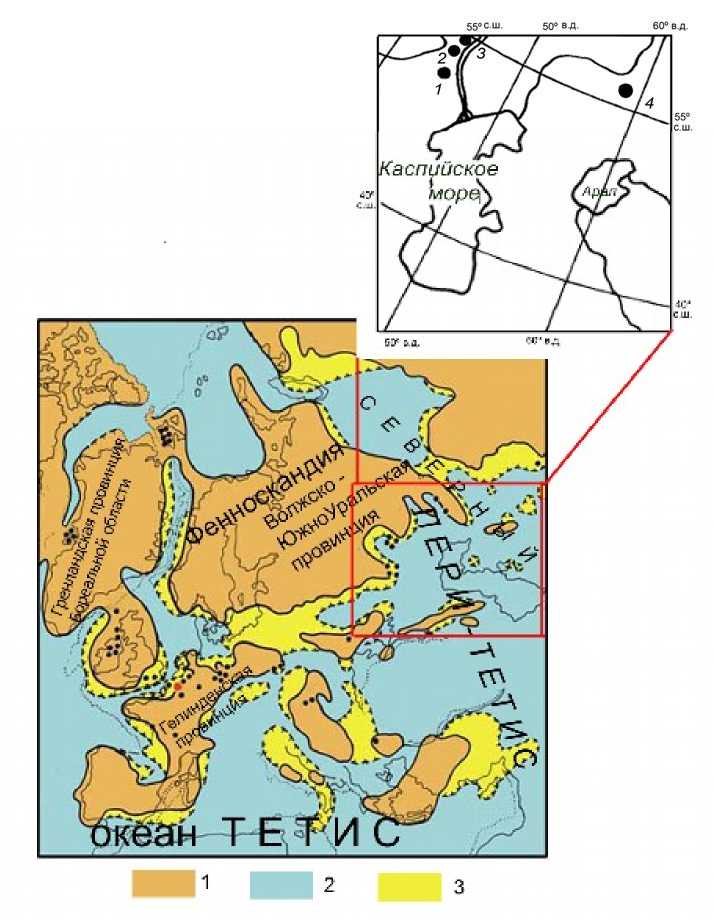
Рис. 2. А. Вверху – местонахождения палеоценовых флор Волжско-Южно-Уральской провинции Тетисовой Гелинденской палеофлористической области: 1 – Камышин, гора Уши; 2 – окрестности г. Вольска, Саратовская обл.; 3 – Ульяновская обл. (Симбирская губ.), 4 – Романкульсай (Южный Урал), рис. С.В. Викулина; Б. Внизу – палеофлористические области и провинции Северо-Западной Евразии по ископаемым флорам высших растений: 1 – суша, 2 – море; 3 – эпиконтинентальные участки мелководного шельфа
А. At thetop : Locations of Paleocene floras of the Volga – Southern Urals province of Tetis Gelinden paleofloristic region (1 – Kamyshin, Mount Ushi; 2 – Volsk neighborhood, Saratov province; 3 – Ulyanovsk (Simbirsk) province; 4 – Romankulsay (Southern Urals), fig. by S.V. Vikulin; Б. At the bottom : Paleofloristic regions and the provinces of North-Western Eurasia by fossil flora of Embryo-phyt: 1 – land, 2 – the sea; 3 – epicontinental shallow shelf areas
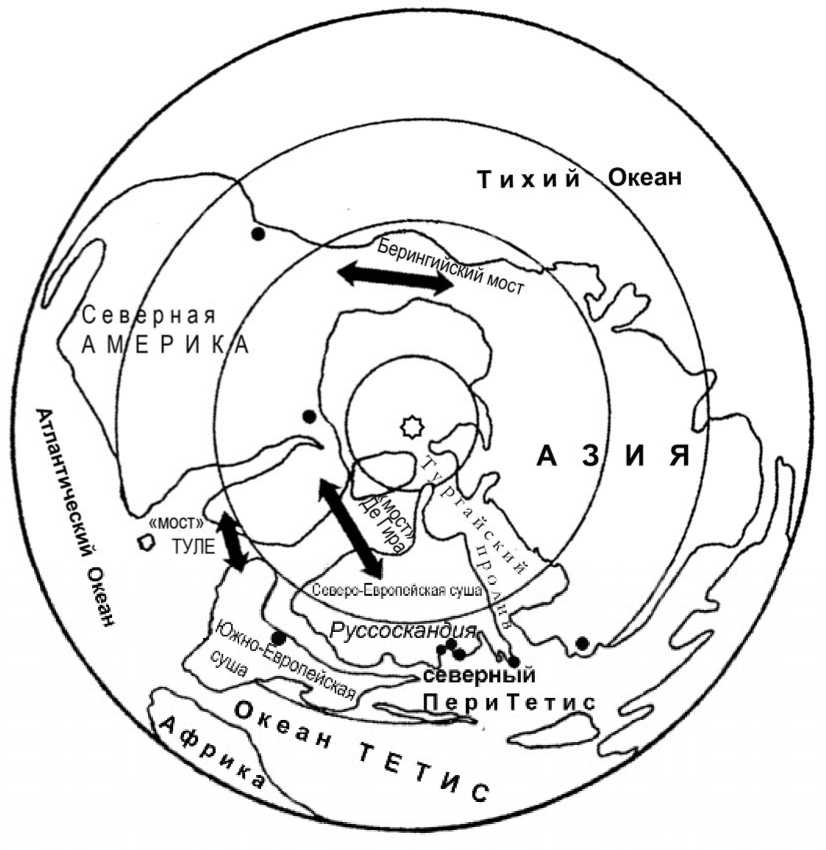
Рис. 3. Палеогеографическая схема очертания морей и суши Северного Полушария в раннем палеогене ( * - полярная проекция). Точками • указаны местонахождения палеоцен-эоценовых макроостатков ископаемых видов Chamaecyparis в Северном полушарии, стрелками указаны пути возможных межконтинентальных миграций (по: Викулин и др., 2010; Liu et al., 2009; Brikiatis Leonidas, 2014; с изменениями и модификациями С.В. Викулина)
Paleogeographic sceme of the seas outlines and the land of the Northern Hemisphere in the early Paleogene ( * - polar projection). The dots • indicate the locations of the Paleocene-Eocene macrofossils of Chamaecyparis in the Northern Hemisphere; the arrows indicate the paths of possible intercontinental migrations (after Vikulin et al, 2010; Liu et al, 2009; Brikiatis Leonidas, 2014, with amendments and modifications by S.V. Vikulin)
нем Поволжье и Южном Урале относятся к рубежу палеоцена и эоцена. На этом рубеже (танет) биосферный кризис, связанный с сильным потеплением (‘термальный максимум’) вызвал появление максимального количества таксонов тропического родства в средних широтах Парижского бассейна, Днепровско-Донецкой впадины, Нижнего Поволжья и Южного Урала. Отсутствие отчетливого зимнего замерзания привело к не- обычайному продвижению субтропических и тропических растений по направлению к полюсам (Тахтаджян, 1966). В Европе тетисо-вая (древне-средиземноморская) палеоценовая паратропическая флора гелинденского экологического типа проcтирается от Южной Англии к юго-востоку, включая Западную и Восточную Европу, и далее через Украину продолжается в Среднее – Нижнее Поволжье и Южный Урал. Далее палеоценовые флоры
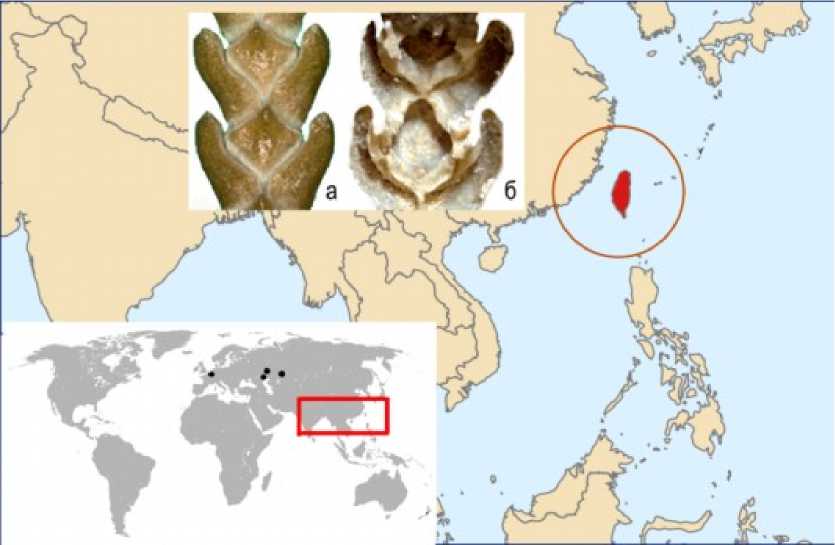
Рис. 4. Географическое распространение в Западной Евразии палеоценового Chamaecyparis belgica и современного C. formosensis на о. Тайвань
Geographical distribution of Paleocene Chamaecyparis belgica in Western Eurasia and modern C. formosensis on Taiwan island

Рис. 5. Карьер Вольск – Шиханы, палеоценовый флороносный слой скрыт осыпью и располагается в подошве карьера, фото С.В. Сергеева
Quarry Volsk – Shikhany, Paleocene flora-bearing layer is covered with a rash and is located at the base of the quarry, photo by S.V. Sergeev
Центральной Азии, Северо-Западного Китая (верховья Иртыша) и Монголии имеют иной, смешанный характер, и относятся к Восточноазиатской флористической области, являясь переходными к гумидным флорам теплоумеренной зоны. Дальнейшая флористическая перестройка при переходе от равномерновлажного на протяжении годового цикла паратропического позднепалеоценового климата к сезонному субтропическому завершилась к началу среднего эоцена (Akhme-tiev, Beniamovski, 2009; Викулин, 2013; Ах-метьев, Запорожец, 2014). Общей особенностью гелинденских палеоценовых флор было и то, что во всех этих местонахождениях на прибрежных островах Северного Перитетиса разнообразие хвойных было невелико, в изученных нами коллекциях они представлены лишь вегетативными побегами Chamaecyparis belgica Sap.
Результаты исследования
СИСТЕМАТИКА
Chamaecyparis belgica Saporta et Marion, 1873
Класс PINOPSIDA Burnett, 1835
Порядок P I N AL ES Gorozh . , 1904 Сем. CUPRESSACEAE Bartlett, 1830 Род CHAMAECYPARIS Spach, 1841
Род Кипари́совик включает шесть видов, распространенных дизъюнктивно во влажных прибрежных местообитаниях Северного полушария (Farjon, 2005). Четыре вида встречаются только в Азии (Chamaecyparis obtusa и Ch. pisifera– эндемики Японии, а Ch. for-mosensisи Ch. taiwanensis –эндемики Тайваня). Характерно, что Ch. taiwanensis часто рассматривается в качестве тайваньской разновидности японского вида C. obtusa(как C. obtusa var. formosana). Два вида (Chamaecyparis thyoides и Chamaecyparis lawsoniana) обитают соответственно на востоке и западе Северной Америки (Schulz et al., 2005). В ископаемом состоянии из отложе- ний палеоцена – эоцена, помимо описываемого бельгийского вида кипарисовика, известны находки других видов этого рода из палеофлор Евразии и Северной Америки (рис. 2-4).
Chamaecyparis belgica Saporta et Marion (рис. 4, б; табл. I, фиг.1, 2; табл. III, фиг. 1-6;
табл. IV, фиг. 1-7; табл. V, фиг. 1)
Chamaecyparis belgica Saporta et Marion 1873, Essai sur la végét. de Gelinden, Belgique: 31, tab. 1, fig. 3 ; ibid. 1878, Révis. de la flore heersienne de Gelinden, Belgique: 21, tab. 1, fig. 6-9; Краснов 1910, Начатк. третичн. фл. юга России: 33, фото в тексте; Байковская 1984, Палеоцен. фл. Романкульсая (Южный Урал); Thujopsis kazachstanica Makul. 1977, Макул-беков 1977, Палеоцен. фл. западн. Казахстана нижн. Поволжья: 92, табл. XIX, фиг. 5, табл. XX, фиг. 8, 9; ibid., sine descr.: Cupressus sp., табл. XIX, фиг. 4.
Г о л о т и п: Saporta et Marion 1873, Essai sur la végét. de Gelinden, Belgique: 31, tab. 1, fig. 3 (Табл. III, фиг. 1) – материалы из Gelinden хранятся в королевском институте естественных наук Бельгии. Некоторые экземпляры – в университетах Льежа и Левена.
Т и п о в о е м е с т о н а х о ж д е н и е (Locus typicus): палеоценовые мергели Ге-линдена, Бельгия. Характер образования флороносных отложений палеоценового "мергеля" из Gelinden связан с накоплением растительных остатков в спокойных водах морского залива-лимана, где морские воды смешивались с пресными водами впадающей реки. Эти условия были благоприятными как для сохранения остатков растений из морской среды – например, Posidonia (Stockmans, 1932a), так и для листьев наземных растений – Dewalquea , Litsea (Stockmans, 1932b, 1960).
Г е о л о г и ч е с к и й в о з р а с т типового местонахождения: основание ланденских слоев, средний – верхний палеоцен, палео-ген4: Heers Formation [Landenien inférieur,
Belgique (De Geyter et al., 2006 ; Schumacker-Lambry, 1976,
84 нижний ландений, Gelinden-Overbroek, Бельгия (Chateauneuf 1976; Geyter et al., 2006; Schumacher-Lambry, 1978; Steubart, 1998; Steurbaut et Nolf, 1986; Steubart et al., 2003)].
М е с т о н а х о ж д е н и я (рис. 2 A, Б): средний – верхний палеоцен, геерсиенские мергели (Heers Formation), [50°46' с.ш., 5°15' в.д. Гелинден5 (Желинден)], окрестности Сент-Трейден (Gelinden6, Sint-Truiden7), провинция Лимбург, восточная Бельгия, палеоцен, окрестности ж.д. ст. Привольская, г. Вольск, Вольский р-н, Саратовская обл. [правый берег Волги в 111 километрах к северо-востоку от Саратова, координаты: 52°2'27" с.ш., 47°23'34" в.д. [по: Краснов, 1910: 33, найдено около ст. Привольской г. Архангельским]; там же, новые сборы: В.П. Моров, С.В. Сергеев, г. Вольск, окрестности поселка Шиханы, Саратовская обл.; палеоцен, Ульяновская область, верховья реки Свияги в пределах Ульяновско-Саратовского прогиба [по: Краснов, 1910: 33, собрано “в Симбирской губ. по р. Сiязи г. Палибиным”]; горизонт глауконитовых песчаников, в. палеоцен, Ро-манколь8 (Романкуль), сай (овраг) в 0,5 км выше устья р. Тогеушсай, Южный Урал, Актюбинская обл., Казахстан.
Многие из вышеупомянутых местонахождений были открыты пионерами изучения поволжского палеогена – А.П. Павловым, А.Д. Архангельским и А.Н. Красновым, а на Южном Урале уже в советское время – геологами и палеоботаниками – А.А. Петренко, Н.Д. Василевской, В.И. Барановым, Н.М. Макул-бековым, И.В. Васильевым, С.Г. Жилиным и М.А. Ахметьевым. Палеоботанические сборы известного русского ботаника Краснова (1910) по палеоценовым хвойным Поволжья, хранящиеся в Санкт-Петербурге в лаборатории палеоботаники БИН РАН, колл. LVI, также как и материалы из колл. Н.Д. Василевской из Романколя (Южный Урал, колл. №
1385, БИН РАН) были заново исследованы (С.В.В.) в настоящей работе (Юж. Урал: табл. II, 1-3); (Поволжье: табл. II, 4-6; табл. V, 1).
Заключение о геологическом возрасте стратиграфически неоднородных отложений (кварцитовидные и глауконитовые песчаники, мергели) Бельгии, Поволжья и Южного Урала в ряде случаев построены на макрофлори-стических комплексах (Байковская, 1984). До середины 1970-х гг. основными таксономическими группами, на которых была основана стратиграфия палеоцена Поволжья, были двустворчатые и брюхоногие моллюски, из ранних слоев палеоцена – фораминиферы, а также палиноморфы, встреченные по всему разрезу поволжского палеоцена. Позднее в биостратиграфии региона стали использовать группы кремневого микропланктона (диатомеи и радиолярии), известковый наннопланктон и диноцисты (Александрова, 2013). Часто фаунистические материалы носят сборный характер и происходят из местонахождений, обычно не связанных с флористическими горизонтами. Поэтому для нижнепалеогеновых отложений с растительными остатками был установлен широкий возрастной диапазон: верхний палеоцен – средний эоцен, без четкой дифференциации стратонов конкретных обнажений осадочных пород. Дальнейшее изучение флор Поволжья (Макулбе-ков, 1977) и Южного Урала (Байковская, 1984) подтверждает флористические связи этих палеоценовых флор с Парижским Бассейном и с флорой Гелиндена в Бельгии.
П а л е о г е о г р а ф и я (рис. 2Б; рис. 3): для Поволжья и Западного Казахстана палеогеновая эпоха характеризовалась сравнительно медленными тектоническими движениями, способствовавшими накоплению многосо-тметровых толщ морских осадков (Геология СССР…, 1970). В это время, в отличие от
Бельгии, где происходило накопление главным образом карбонатных пород (мергелей, известняков, карбонатных глин), на территории Среднего – Нижнего Поволжья накапливались преимущественно песчаные отложения и, особенно в нижней части разреза, кремниевые породы.
От палеоцена к олигоцену размеры трансгрессий нарастали и все большие районы покрывались водами палеогеновых бассейнов. В палеоцене значительную часть территории Южной России и Западного Казахстана в пределах Руссоскандии покрывало море, среди которого возвышались поднятия отдельных островов Поволжья (Павлов, 1896, 1897; Геология СССР…, 1970), и на юго-востоке древнего континента Руссоскандии крупным полуостровом вдавались южная оконечность Уральских гор – Мугоджары (Барбот деМарни, 1874; Архангельский, 1905; Архангельский, Доброе, 1913; Беньямовский, 1995), рис. 3.
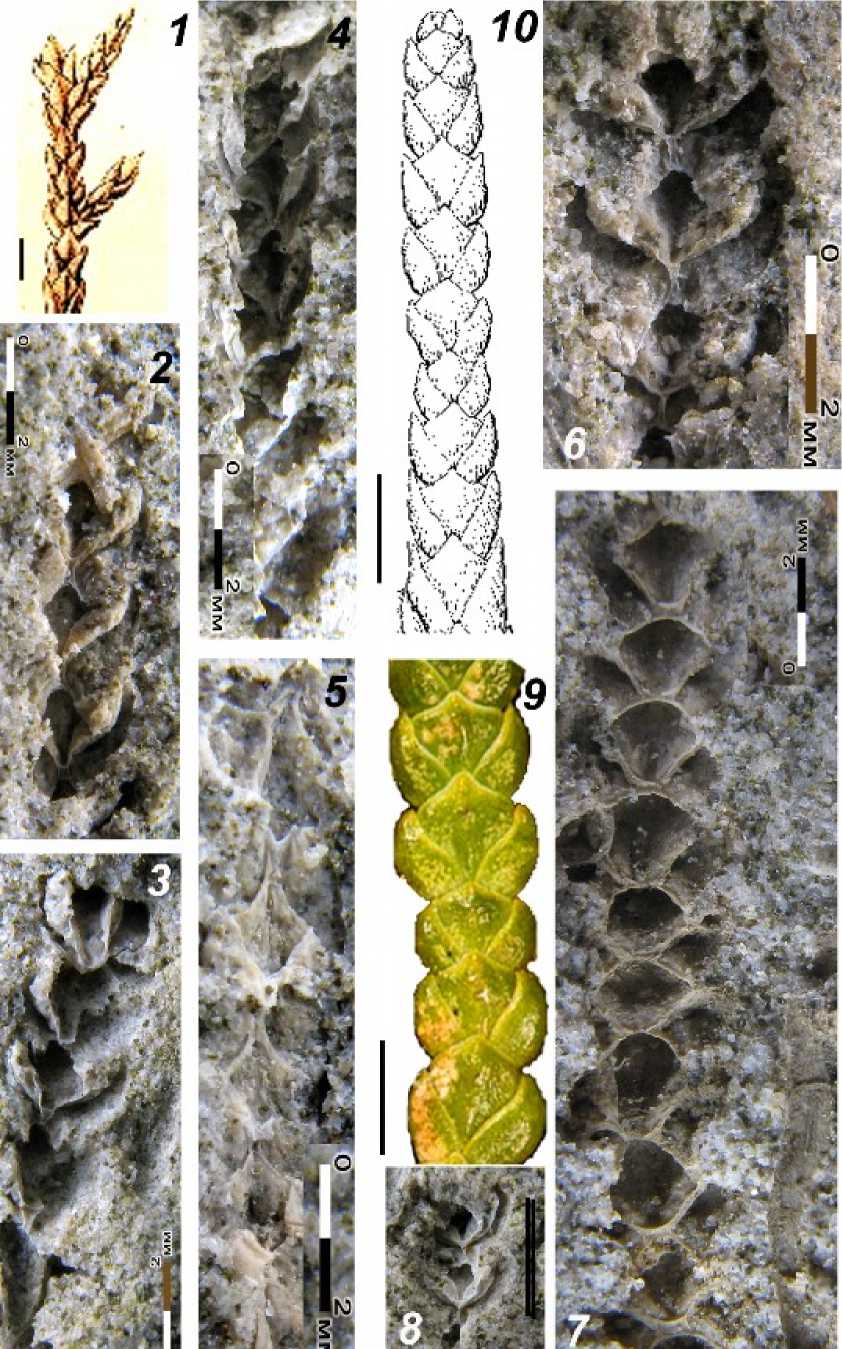
О п и с а н и е морфологии побегов. Побеги сохранились объемно, в виде вдавленных в песчаник отпечатков. По-видимому, в процессе фоссилизации минерализованные побеги, растворившись, оставили после себя полости в виде скульптурных отпечатков. Образцы всех исследованных отпечатков побегов Chamaecyparis belgica как из палеоценовых местонахождений Волжского бассейна, так и Южного Урала, весьма близки к образцам из Гелиндена, приведенным Сапорта и Марион (Saporta et Marion, 1873, Pl.1, fig. 3 и Saporta, Marion, 1878, Pl.1, fig. 7). Всего, с учетом новых материалов, полученных В.П. Моровым и С.В. Сергеевым (табл. III, фиг. 36; табл. IV, фиг. 2-8), собранных в местонахождениях окрестностей г. Вольска: Шиханы, Саратовская обл., совокупно имеется более 20 отпечатков фрагментов плоских неветвя-щихся облиственных конечных (терминальных) декуссатных побегов с чешуевидными листьями, прижатыми к побегу не плотно. Фрагменты побегов достигают длины 20 мм. Ширина сегмента в среднем около 1.4 мм, а высота сегмента – 2.2 мм. Филлотаксис (листорасположение) побегов декуссатный (пе- рекрёстно-парный): листья двух видов – фациальные и латеральные. Сохранившиеся фрагменты побегов состоят от 2 до 17 ‘узлов’-сегментов. Листья диморфные: 1) маргинальные (= латеральные), расположенные по бокам, и 2) фациальные, расположенные в одной плоскости. Маргинальные листья – длинные и узкие, изогнутые округло-серповидно (с округло-выпуклой нижней и округловогнутой верхней поверхностью), и с тупозаостренной или округлой верхушкой, достигающей уровня расположения верха фациальных листьев 2.0-2.5 мм длины, 1.0-1.5 мм ширины, сжатые двусторонне, в очертании овально удлиненные. Фациальные листья супротивные, ромбические, или округлоромбические, с округленной верхушкой, которая расположена на уровне вершин маргинальных листьев.
Сравнение и обсуждение
Отпечатки побегов из Поволжья и Южного Урала близки к образцам из Гелиндена, судя по изображениям побегов, приведенных Сапорта и Марионом (Saporta et Marion, 1873: табл. I, фиг. 3; Saporta et Marion,1878: табл. I, фиг. 7). Полная идентичность между собой для всех российских образцов из Поволжья и Южного Урала несомненна, также как и их морфологическая близость к тайваньскому кипарисовику Chamaecyparis for-mosensis. Однако поскольку материалы из бельгийского местонахождения известны лишь по рисункам из монографий XIX в. (Saporta et Marion, 1873, 1878) [табл. III, фиг. 1, 2; табл. IV, фиг. 1], для полной уверенности в их отнесении к Chamaecyparis [табл. III, фиг. 3, 8], а не Thuja [табл. III, фиг. 9] или Thujop-sis[табл. V, фиг. 3], требуется фотографирование их побегов [close-up] и детальное морфологическое сопоставление с современными кипарисовыми [табл. III, фиг. 8-9; табл. IV, фиг. 9-10; табл. V, фиг. 2-4]. В случае утраты образцов в Бельгийских депозитариях, необходимы новые сборы в гелинденских отложениях. Предварительно, до детального переис-следования бельгийского материала по палеоценовым чешуелистным хвойным, мы бу- дем продолжать условно использовать биномен – Chamaecyparis belgicaSaporta et Marion. для обозначения чешуелистных побегов из палеоцена Поволжья и Южного Урала.
Как отмечала еще Т.Н. Байковская (1984: 22), ошибочно описанный Н.М. Макулбеко-вым9 (1977: 92) без каких-либо ссылок на предшествующие находки кипарисовых хвойных в палеоценовых флорах Гелиндена, Южного Урала или Поволжья – Thujopsis kazachstanica Makul. из Романколя на Южном Урале (Макулбеков, 1977: табл. XIX, 5; табл. XX, 8, 9), в действительности идентичен ки-парисовнику Chamaecyparis belgica Sap. et Marion, описанному позже из того же самого Романкольского местонахождения Т.Н. Байковской (1984: 21). Отнесение Н.М. Макул-бековым (1977) побегов из Романколя к роду Thujopsis не правомерно и ботанически не обосновано. Исследованные нами ископаемые чешуелистные побеги, как из самого Ро-манколя, так и из классических и новых палеоценовых местонахождений Поволжья, имеют иную – серповидно-округлую форму маргинальных листьев, свойственную тайваньскому кипарисовнику Chamaecyparis formosensis(Schulz et al., 2005).Длятуевика и видов туи характерна иная
–
ладьевидная
форма маргинальных листьев, заметно расширяющихся книзу. Хотя поверхностное сходство палеоценовых побегов Ch. belgica наблюдается и с такими родами сем. кипарисовых, как Thujopsis (L.f.) Siebold et Zucc., Thuja L. значительно большее сходство наблюдается с видами рода Chamaecyparis Spach., как по морфологии побегов, так и шишек. К ископаемому виду Ch. belgicaпобе-ги Chamaecyparis formosensisблизки своими округло-серповидными листьями с затупленно-округлой верхушкой. Причем, фациальные листья и маргинальные узко-серповидные листья как у ископаемого вида Ch. belgica, так и у Ch. formosensis, достигают одинакового уровня. И хотя по этому признаку сходство наблюдается с некоторыми видами Thujopsis и Thuja, их маргинальные листья заметно расширяются книзу, имеют ладьевидную форму, тогда как у Chamaecyparisformosensis и ископаемого Ch. belgica маргинальные листья округло-узкосерповидные. Бóльшая обоснованность отнесения рассмотренных ископаемых побегов к Chamaecyparis подтверждается также незначительной величиной их хвой (листьев). Особенно характерны для кипарисовиков отстающие от побега узкие боковые хвои, что нетипично для видов рода Thuja. Самый ближайший по морфологии вегетативных побегов к ископаемому ге-линденскому кипарисовнику (Ch. belgica) – это современный родственный вид Chamae-cyparis formosensis, произрастающий на высотах от 1 км и выше в горных смешанных муссонных субтропических лесах острова Тайвань. На этом субтропическом острове в Южно-Китайском море, помимо кипарисо-вика, сохраняются и другие реликтовые хвойные: куннингамия и тайвания – Cunningha-mia lanceolata var. konishii и Taiwania cryp-tomerioides. Тайваньский кипарисовик (Ch. formosensis) – это весьма крупное дерево, достигающее высоты 65 м, при диаметре ствола 6.5 м; растет в смешанных лесах из эндемичного камфорного лавра, ели, пихты и древовидных папоротников (Li, 1963). Небольшие округлые шишки Ch. formosensis весьма похожи на изображения ископаемой шишки кипарисовика из Гелиндена (табл. III, фиг. 2, 7).
В ы в о д ы. Подтверждается представление А.Л. Тахтаджяна (1966) о том, что палеоценовые флоры гелинденского экологического типа, распространенные в Европе от Парижского бассейна в Бельгии до Южного Урала, составляли отдельный фитохорион в пределах палеоценовой паратропической Те-тисовой области (Викулин, 2010, 2011, 2013, 2014). Выявленные нами таксономические

Таблица I. Фрагменты побегов кипарисовика (1а, б, в и 2а) на образцах с ископаемым растительным детритом. Номера образцов из коллекции ИЭВБ РАН – карьер Вольск – Шиханы, палеоценовый флороносный слой: фиг. 1 - № 3636 – большой (фото двух сторон В.П. Морова) и фиг. 2 – № 3637 – маленький треугольный, фото В.П. Морова; коллектор В.П. Моров, 1а, б, в и 2а – фото Д.В. Варенова

Givi Л fasti
Таблица II. 1-2 – Chamaecyparis belgica Saporta et Marion, отпечаток побега, в. палеоцен, Роман-коль, Ю. Урал, фото С. В. Викулина, обр. 116, колл. 1385 [БИН РАН]; Байковская, 1984: 21, табл. I, фиг. 3, 3a; 3 – C. belgica, то же, фото С.В. Викулина, обр. 117, колл. 1385 [БИН РАН]; Байковская, 1984: 21, табл. I, фиг. 4, 4a; 4-6 – C. belgica, палеоцен, окрестности ж.д. ст. Привольская, г. Вольск, Саратовская обл., обр. 3, колл. 56 [БИН РАН]; Краснов 1910, Начатк. третичн. фл. юга России: 33, фото в тексте. Масштабн. линейка – 1, 3, 5 – 10 мм; 2, 4, 6 – 2 мм

Таблица III. 1, 2 – Chamaecyparis belgica Saporta et Marion, отпечаток побега и шишка, Heers Formation, ср.-в. палеоцен, Gelinden, восточная Бельгия, 1– тип, отпечаток побега: Saporta et Marion 1873: 31, tab. 1, fig. 3; 2– шишка: ibid. 1878: 21, tab. 1, fig. 9; 3-5 –C. belgica , отпечатки побегов, палеоцен, Саратовская область, Вольск – Шиханы, карьер, палеоценовый флороносный слой в подошве карьера, коллектор С.В. Сергеев; 6 –C. belgica , палеоцен, Саратовская область, Шиханы, фрагмент побега (два сегмента) на обр. № 3637 из коллекции ИЭВБ РАН, фото Д.В. Варенова, мас-штабн. линейка – 1 мм; 7 – C. formosensis Matsum., шишка, эндемик влажных горных лесов о. Тайвань; 8 – C. lawsoniana , побеги, Калифорния, Орегон, в долинах горных рек; 9 – Thujaoccidentalis , побеги, восток Сев. Америки
"ЧТ .^к^ЯКи в*Л№ТК§Ж УН6^^.,-#‘чЗ •гУНк.. Л^Я№
Таблица IV. 1 – Chamaecyparis belgica Saporta et Marion, 1878: 21, tab. 1, fig. 6 – отпечаток побега;
2-8 – C. belgica, палеоцен, Саратовская обл., Шиханы, фрагменты побегов (от 2-х до 12 сегментов) на обр. № № 3636, 3637 из колл. ИЭВБ РАН, фото Д.В. Варенова, масштабн. линейка ( 1-8 ) – 2 мм

Фототаблица V. 1 – Chamaecyparis belgica, палеоцен, окрестности ж.д. ст. Привольская, г. Вольск, Саратовская обл., обр. 3, колл. 56 [БИН РАН]; Краснов 1910, Начатк. третичн. фл. юга России: 33, фото в тексте; фото С.В. Викулина; 2 – C. formosana, сегмент вегетативного побега, эндемик влажных горных лесов о. Тайвань; 3 – Thujopsisdolabrata,леса среднегорного пояса, о. Хонсю, Япония; 4– Thujaplicata, береговая полоса Тихого океана с-з Сев. Америки; масштаб. линейка (1-4) – 1 мм связи и экологические особенности современных видов-аналогов подтверждают па-леофлорогенетические представления акад. А.Л. Тахтаджяна (1966) о том, что Европейские флоры гелинденского экологического типа происходят из древней субтропической флоры Юго-Восточной Азии.
Список литературы Chamaecyparis belgica - хвойное из палеоценовой Волжско-Южноуральской провинции Гелинденской палеофлористической области
- Александрова Г.Н. Диноцисты палеоцена Среднего и Нижнего Поволжья: cтратиграфия палеообстановки: автореферат дис.. канд. геолого-минер. наук. М., 2013, 22 c.
- Архангельский А.Д. Некоторые данные о палеоценовых отложения Симбирской и Саратовской губерний. Материалы для геологии России, 1905, т. XXII, вып. 2, с. 385-415.
- Архангельский А.Д., Доброе С.А. Геологический очерк Саратовской губернии. М.: Печатня С.П. Яковлева, 1913, 256 с.
- Ахметьев М.А., Беньямовский В.Н. Палеоцен и эоцен Российской части Западной Евразии. Стратиграфия. Геологическая корреляция, 2006, т. 14, № 1, с. 69-93.
- Ахметьев М.А., Запорожец Н.И. События палеогена в Центральной Евразии, их роль в развитии флоры и растительного покрова, смещении границ фитохорий и изменениях климата. Стратиграфия. Геологическая корреляция, 2014, т. 22, № 3, с. 90-114.
