Частоты ядерных аномалий в эритроцитах карпа при воздействии кадмия в сочетании с низкочастотным электромагнитным полем

Автор: Крюков В.И.
Журнал: Биология в сельском хозяйстве @biology-in-agriculture
Рубрика: Современные вопросы экологической генетики
Статья в выпуске: 1 (26), 2020 года.
Бесплатный доступ
Генетические эффекты одновременного действия нескольких мутагенных факторов окружающей среды на организм являются важной проблемой экологической генетики. В статье излагаются результаты исследования частот микроядер и других ядерных аномалий в эритроцитах годовалых карпов после суточного их пребывания в воде, содержащей ионы кадмия в концентрациях 0,0005, 0,001, 0,002, 0,004, 0,005, 0,008 мг/л при одновременном воздействии на них переменного электромагнитного поля частотой 50 Гц и напряжённостью, соответственно, 25, 50, 100, 200, 250 и 400 А/м. У интактных карпов контрольной группы частота микроядер была равной 0,47% и суммарная частота всех аномалий - 0,75%. У карпов 6 экспериментальных групп, подвергнутых одновременному воздействию двух факторов, частоты эритроцитов с микроядрами составили 0,76±0,12, 0,90±0,13, 0,96±0,14, 0,99±0,14 1,03±0,14, 0,98±0,14%, а суммарные частоты эритроцитов с аномалиями всех типов - 1,16±0,15, 1,33±0,16, 1,48±0,17, 1,57±0,18, 1,65±0,18, 1,52±0,18% соответственно. Обнаружены статистически достоверные различия между частотами ядерных аномалий в эритроцитах рыб экспериментальных групп, как по сравнению с контролем, так и с соответствующими вариантами раздельного воздействия на них кадмия и электромагнитного поля. Сделан вывод, что сочетанное воздействие на эритроциты рыб кадмия и низкочастотного электромагнитного поля оказывает более сильный мутагенный эффект, чем сумма эффектов этих факторов, действующих отдельно.
Рыбы, эритроциты, микроядра, ядерные аномалии, кадмий, низкочастотное электромагнитное поле, мутагенез
Короткий адрес: https://sciup.org/147231000
IDR: 147231000
Frequencies of the nuclear anomalies in the carp erythrocytes after exposure to cadmium in conjunction with a low-frequency electromagnetic field
An important problem of ecological genetics is the genetic consequences of the simultaneous action of several mutagenic environmental factors on a living organism. This article presents the results of a study of the frequencies of micronuclei and other nuclear anomalies in the erythrocytes of carps after a 24-hour exposure to cadmium ions and a low-frequency electromagnetic field. The concentration of cadmium ions was 0.0005, 0.001, 0.002, 0.004, 0.005, 0.008 mg / L. The intensity of the low-frequency (50 Hz) electromagnetic field in these 6 variants was 25, 50, 100, 200, 250 and 400 A / m, respectively. The intact carps from the control group had 0.47% of the erythrocytes with micronuclei and 0.75% of cells with all nuclear abnormalities in total. Carps from 6 experimental groups that were simultaneously exposed to two factors had erythrocyte frequencies with micronuclei 0.76 ± 0.12, 0.90 ± 0.13, 0.96 ± 0.14, 0.99 ± 0.14 1.03 ± 0.14, 0.98 ± 0.14%, and the total frequencies of all nuclear anomalies -1.16 ± 0.15, 1.33 ± 0.16, 1.48 ± 0.17, 1, 57 ± 0.18, 1.65 ± 0.18, 1.52 ± 0.18%, respectively. Statistically significant differences between the frequencies of nuclear anomalies in the erythrocytes of fish of the experimental groups were found, both in comparison with the control and with the corresponding variants of the separate exposure of cadmium and the electromagnetic field to them. It was concluded that the combined effect of cadmium and a low-frequency electromagnetic field on fish erythrocytes has a stronger mutagenic effect than the sum of the effects of these factors acting separately.
Текст научной статьи Частоты ядерных аномалий в эритроцитах карпа при воздействии кадмия в сочетании с низкочастотным электромагнитным полем
Введение . Электрификация и автоматизация сельскохозяйственного производства существенно изменила среду обитания сельскохозяйственных животных. Развитие индустриального рыбоводства позволило выращивать рыбу в установках замкнутого водоснабжения при почти полной автоматизации процессов жизнеобеспечения. Все приборы и механизмы, используемые в современных рыбоводных и животноводческих комплексах, работают на электроэнергии и являются источниками электромагнитного излучения различной частоты и интенсивности. Со-
Summary . An important problem of ecological genetics is the genetic consequences of the simultaneous action of several mutagenic environmental factors on a living organism. This article presents the results of a study of the frequencies of micronuclei and other nuclear anomalies in the erythrocytes of carps after a 24-hour exposure to cadmium ions and a low-frequency electromagnetic field. The concentration of cadmium ions was 0.0005, 0.001, 0.002, 0.004, 0.005, 0.008 mg / L. The intensity of the low-frequency (50 Hz) electromagnetic field in these 6 variants was 25, 50, 100, 200, 250 and 400 A / m, respectively. The intact carps from the control group had 0.47% of the erythrocytes with micronuclei and 0.75% of cells with all nuclear abnormalities in total. Carps from 6 experimental groups that were simultaneously exposed to two factors had erythrocyte frequencies with micronuclei 0.76 ± 0.12, 0.90 ± 0.13, 0.96 ± 0.14, 0.99 ± 0.14 1.03 ± 0.14, 0.98 ± 0.14%, and the total frequencies of all nuclear anomalies –1.16 ± 0.15, 1.33 ± 0.16, 1.48 ± 0.17, 1, 57 ± 0.18, 1.65 ± 0.18, 1.52 ± 0.18%, respectively. Statistically significant differences between the frequencies of nuclear anomalies in the erythrocytes of fish of the experimental groups were found, both in comparison with the control and with the corresponding variants of the separate exposure of cadmium and the electromagnetic field to them. It was concluded that the combined effect of cadmium and a low-frequency electromagnetic field on fish erythrocytes has a stronger mutagenic effect than the sum of the effects of these factors acting separately.
здание благоприятных условий содержания значительно повышает продуктивность животных. Вместе с тем, наличие в животноводческих помещениях электропроводки и размещение в них электрических приборов и оборудования повысило фоновые уровни электромагнитных полей и усилило их влияние на продукционные свойства и наследственность животных. Интенсификация производства в промышленном рыбоводстве сопряжена с усилением электромагнитных полей в зоне выращивания рыбы. Последствия возрастания уровня электромагнитных полей в среде обитания сельскохозяйственных животных необходимо исследовать. Медиками накоплены многочисленные факты как благоприятного, так и вредоносного влияния электромагнитных полей различных частот на здоровье и физиологические процессы у человека. Роль этих полей в жизнедеятельности сельскохозяйственных животных изучена крайне слабо. Поэтому биологические эффекты антропогенного повышения электромагнитного фона биосферы требуют особенного внимания.
Другой важной проблемой экологии является загрязнение окружающей среды тяжёлыми металлами. К ним относят металлы с плотностью более 5 г/см3. Экологи уделяли большое внимание миграции и накоплению тяжёлых металлов в трофических цепях. Последние 25 лет реки и озера стали испытывать сильное загрязнение тяжёлыми металлами из-за всё возрастающего поступления стоков промышленных, сельскохозяйственных и бытовых стоков. Генетические механизмы действия солей тяжёлых металлов изучены ещё недостаточно. Не всегда понятны механизмы индукции ими мутаций. Не изучены отдалённые генетические последствия хронического воздействия повышенных концентраций тяжёлых металлов и не разработана общая теория их мутагенности.
У тяжёлых металлов обнаружена способность влиять на эпигеном организмов путём изменения степени метилирования ДНК и модификации гистоновых белков, влияющие на структуру хроматина и микроРНК [1]. Онкологи обнаружили доказательства сильного влияния тяжёлых металлов на частоту возникновения некоторых раковых заболеваний посредством эпигенетических влияний на активность генома.
Одним из тяжёлых металлов, загрязняющих окружающую среду, является кадмий. Токсикология этого металла хорошо изучена, но его мутагенные свойства исследованы ещё недостаточно. К настоящему времени опубликован ряд работ о генетических эффектах кадмия, но эти сведения пока не дают полного понимания его влияния на наследственность живых организмов. Поэтому необходимо дальнейшее изучение мутагенности различных соединений кадмия для организмов разного эволюционного уровня.
В современных условиях все живые организмы, включая человека и сельскохозяйственных животных, подвергаются воздействию многих антропогенных факторов одновременно. Такое одновременное воз- действие на организмы факторов химической и физической природы называют сочетанным влиянием факторов. Результат сочетанного действия нескольких факторов может отличаться от суммы результатов индивидуального их влияния. Реакции организмов на сочетанное действие факторов является важной проблемой токсикологии, биофизики и генетики. Одним из реально встречающихся в сельскохозяйственном производстве сочетаний факторов является воздействие на организмы тяжёлых металлов и электромагнитных полей низкой частоты.
Целью данного исследования был анализ сочетанного действие ионов кадмия в различных концентрациях и низкочастотного электромагнитного поля различной напряжённости на частоту возникновения микроядер и иных ядерных аномалий в эритроцитах периферической крови карпа. В работе описаны частоты микроядер и иных ядерных аномалий в периферической крови карпа, возникающие после суточного воздействия ионов кадмия в концентрациях 0, 0,0005, 0,001, 0,002, 0,004, 0,005, 0,008 мг/л и электромагнитного поля частотой 50 Гц и напряжённостью 0, 25, 50, 100, 200, 250 и 400 А/м при сочетанном действии этих двух факторов.
Материалы и методы исследования
Объектом исследования были эритроциты годовалых карпов ( Cyprinus carpio ), массой 17-24 г. Процесс адаптации рыб к условиям лабораторного содержания, режимы и условия изолированного воздействия на рыб ионов кадмия и НЧ ЭМП подробно описаны ранее [2, 3]. Контрольный вариант (интактные рыбы) был один для всех опытных групп. Подопытные животные были разделены на 3 группы. Рыб первой группы подвергали 24 часовому воздействию раствора нитрата кадмия ( Cd(NO 3 ) 2 ×4H 2 O ) в концентрациях 0,0005, 0,001, 0,002, 0,004, 0,005, 0,008 мг/л. Расчёт концентраций соли выполняли по действующему веществу – Cd+2.
На рыб второй группы в течение суток воздействовали низкочастотным (50 Гц) электромагнитным полем различных (25, 50, 100, 200, 250 и 400 А/м) напряжённостей.
Рыб третьей группы подвергали в течение суток сочетанному воздействию указанных двух факторов одновременно в следующих сочетаниях (табл. 1)
Таблица 1 – Варианты сочетанного действия на рыб различных концентраций кадмия и электромагнитного поля
|
Номер экспериментальной подгруппы |
0 (К)* |
1 |
2 |
3 |
4 |
5 |
6 |
|
Концентрация Cd+2 в мг/л |
0 |
0,0005 |
0,001 |
0,002 |
0,004 |
0,005 |
0,008 |
|
Напряжённость магнитного поля, А/м |
0 |
25 |
50 |
100 |
150 |
200 |
250 |
* – К – контроль, интактные рыбы.
После окончания суточного воздействия исследуемых факторов рыбу пересаживали в аквариумы с чистой водой, и выдерживали следующие 24 часа для реализации индуцированных аномалий. По истечении указанного срока готовили мазки крови. Действие каждого фактора отдельно и в их сочетании для каждого варианта проводили в 2-х повторностях по 4 рыбы в каждой. Из каждой повторности случайным об- разом отбирали по 3 рыбы, кровь которых использовали для приготовления мазков. В мазке крови каждой рыбы просматривали не менее 3000 эритроцитов. Таким образом, частоту аномалий в каждом варианте опыта рассчитывали после анализа не менее 18000 клеток от 6 рыб.
Статистическую значимость различий в частотах нарушений между контролем и различными вариантами опыта определяли после φ-преобразования частот аномалий [4, с. 166-169]. Все расчёты были выполнены с использованием электронных таблиц MS Excel.
Результаты и обсуждение
Детальный анализ частот ядерных аномалий, индуцируемых НЧ ЭМП и нитрата кадмия, при их индивидуальном воздействии на рыбу, приведён в предыдущих публикациях [2, 3]. Поэтому в таблицах 2 и 3 приведены лишь итоговые результаты вычисления частот аномалий. Данные указанных двух публикаций использованы в настоящей работе потому, что все опыты были проведены в одно и то же время и в одних и тех же экспериментальных условиях. Различались лишь сроки проведения микроскопического ана- лиза сохраняемых микроскопических препаратов и сроки опубликования результатов этого анализа.
Спонтанные частоты возникновения микроядер в эритроцитах карповых рыб могут варьировать в зависимости от видовой принадлежности и условий обитания. Например, у карасей, обитающих в разных экологических условиях (водоёмы Брянского рыбзавода и устье реки Сулак) установлено трёхкратное различие частот микроядер 0,3±0,05 и 1,2±0,21‰, соответственно [5]. При проведении лабораторных исследований общепринятая практика адаптации рыб перед экспериментом и использование контрольной выборки нивелирует проблему варьирования спонтанных частот образования микроядер в выборках рыб, используемых в опытах.
При одновременном воздействии на карпов возрастающих концентраций ионов кадмия и напряжённости НЧ ЭМП частоты микроядер в эритроцитах возрастали от 0,76% до 1,03%. Лишь в последнем варианте опыта (0,008 мг/л кадмия и 400А/м) наблюдали некоторое снижение частоты микроядер до 0,98% (рис. 1, а). При сравнении частот микроядер, полученных в этой серии экспериментов, с частотами, полученными в контроле, установлены статистически значимые различия во всех вариантах при P≤0,001.
Таблица 2 – Частоты аномалий ядер (в %) в эритроцитах карпов после суточного воздействия низкочастотного переменного электромагнитного поля различной напряжённости
|
Напряженность ЭМП, А/м |
Всего изучено клеток |
Всего аномальных клеток |
В числе аномальных клеток |
|||||
|
с микроядрами |
двуядерных |
в стадии амитоза |
с пузырящимися (blebbed) ядрами |
с лопастными (lobed) ядрами |
с зазуб-реннми (notched) ядрами |
|||
|
Контроль |
19813 |
0,75±0,01 |
0,47±0,01 |
0,08±0,01 |
0,06±0,01 |
0,005±0,01 |
0,02±0,02 |
0,11±0,01 |
|
25 |
21207 |
0,76±0,01 |
0,48±0,01 |
0,08±0,01 |
0,07±0,01 |
0,042±0,01* |
0,01±0,01 |
0,13±0,01 |
|
50 |
19569 |
0,94±0,01* |
0,54±0,01 |
0,08±0,01 |
0,07±0,01 |
0,072±0,01* |
0,03±0,02 |
0,15±0,01 |
|
100 |
19112 |
1,10±0,01* |
0,61±0,01 |
0,12±0,01 |
0,07±0,01 |
0,094±0,01* |
0,05±0,03 |
0,16±0,01 |
|
200 |
24103 |
2,03±0,01* |
0,61±0,01* |
0,11±0,01 |
0,10±0,01 |
0,100±0,01* |
0,07±0,03* |
0,19±0,01* |
|
250 |
19112 |
1,21±0,01* |
0,62±0,01 |
0,13±0,01 |
0,12±0,01 |
0,120±0,01* |
0,08±0,04* |
0,20±0,01* |
|
400 |
22880 |
1,26±0,01* |
0,63±0,01* |
0,15±0,01 * |
0,12±0,01* |
0,120±0,01* |
0,05±0,03 |
0,19±0,01* |
Примечание. Величины, статистически достоверно отличающиеся от контрольной: * – при Р<0,05;
Таблица 3 – Частоты аномалий ядер (%) в эритроцитах карпов после суточного воздействия ионов кадмия в различных концентрациях
|
Концентрация Cd+2, мг/л |
Всего изучено клеток |
Всего аномальных клеток |
В числе аномальных клеток |
|||||
|
с микроядрами |
двуядерных |
в стадии амитоза |
с пузырящимися (blebbed) ядрами |
с лопастными (lobed) ядрами |
с зазубреннми (notched) ядрами |
|||
|
Контроль |
19813 |
0,75±0,01 |
0,47±0,10 |
0,08±0,01 |
0,06±0,01 |
0,005±0,001 |
0,02±0,02 |
0,11±0,05 |
|
0,0005 |
18915 |
0,80±0,02 |
0,50±0,10 |
0,07±0,01 |
0,05±0,01 |
0,021±0,002 |
0,05±0,03 |
0,11±0,05 |
|
0,001 |
19336 |
0,82±0,02 |
0,51±0,10 |
0,06±0,01 |
0,09±0,01 |
0,026±0,002 |
0,02±0,02 |
0,12±0,05 |
|
0,002 |
18487 |
0,90±0,02 |
0,51±0,10 |
0,10±0,01 |
0,09±0,01 |
0,027±0,002 |
0,06±0,04* |
0,11±0,06 |
|
0,004 |
19137 |
1,02±0,02** |
0,55±0,10 |
0,13±0,01 |
0,10±0,01 |
0,021±0,002 |
0,03±0,02 |
0,19±0,05* |
|
0,005 |
18981 |
1,19±0,02*** |
0,63±0,11* |
0,12±0,01 |
0,13±0,01* |
0,032±0,003* |
0,06±0,04* |
0,22±0,07* |
|
0,008 |
19045 |
0,89±0,02 |
0,54±0,10 |
0,07±0,01 |
0,10±0,01 |
0,032±0,003* |
0,06±0,04* |
0,08±0,04 |
Примечание. Величины, статистически достоверно отличающиеся от контрольной: * – при Р<0,05; ** – при
Р<0,01 и *** – при Р<0,001
Таблица 4 – Количества и частоты аномалий ядер (%) в эритроцитах карпов после их суточного пребывания в воде, содержащей кадмий в различных концентрациях, при одновременном воздействии НЧ ЭМП различной напряжённости
|
Концентрация Cd+2, мг/л |
Напря жён-ность ЭМП, А/м |
Всего изучено клеток |
Всего аномальных клеток |
В числе аномальных клеток |
|||||
|
с микроядрами |
двуядерных |
в стадии амитоза |
с пузырящимися (blebbed) ядрами |
с лопастными (lobed) ядрами |
с зазуб-реннми (notched) ядрами |
||||
|
Контроль |
19813 |
149 0,75±0,01 |
94 0,47±0,10 |
16 0,08±0,01 |
12 0,06±0,01 |
1 0,005±0,001 |
4 0,02±0,02 |
22 0,11±0,05 |
|
|
0,0005 |
25 |
19019 |
221 1,16±0,15*** |
145 0,76±0,12*** |
28 0,15±0,04 |
11 0,06±0,03 |
4 0,021±0,021 |
6 0,03±0,03 |
27 0,14±0,05 |
|
0,001 |
50 |
18939 |
251 1,33±0,16*** |
170 0,90±0,13*** |
25 0,13±0,05* |
19 0,10±0,05 |
5 0,026±0,023 |
5 0,03±0,02 |
27 0,14±0,05 |
|
0,002 |
100 |
19275 |
285 1,48±0,17*** |
185 0,96±0,14*** |
31 0,16±0,05* |
27 0,14±0,05* |
6 0,031±0,025* |
6 0,03±0,02 |
30 0,16±0,06 |
|
0,004 |
200 |
19003 |
298 1,57±0,18*** |
189 0,99±0,14*** |
30 0,16±0,06 |
35 0,18±0,06*** |
6 0,032±0,025* |
7 0,04±0,03 |
31 0,16±0,06 |
|
0,005 |
250 |
18954 |
312 1,65±0,18*** |
195 1,03±0,14*** |
38 0,20±0,06** |
36 0,19±0,06*** |
3 0,016±0,018 |
6 0,03±0,03 |
34 0,18±0,06 |
|
0,008 |
400 |
18915 |
287 1,52±0,18*** |
185 0,98±0,14*** |
35 0,19±0,06* |
31 0,16±0,06** |
7 0,037±0,027* |
9 0,05±0,03 |
20 0,11±0,05 |
Примечание. Величины, статистически достоверно отличающиеся от контрольной: * – при Р<0,05; ** – при Р<0,01 и *** – при Р<0,001
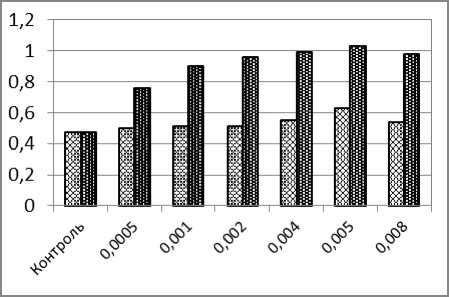
б
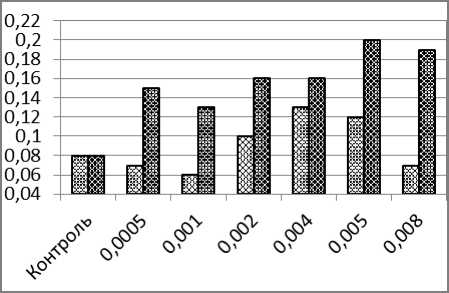
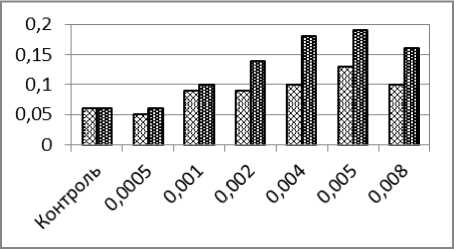
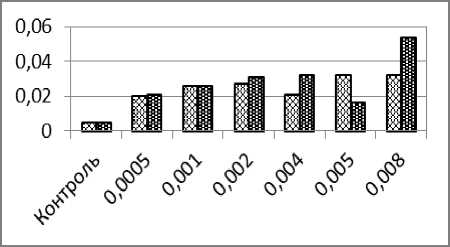
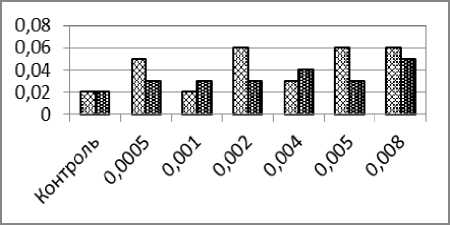
Рис. 1. – Частоты образования ядерных аномалий в эритроцитах карпа (%) после суточного воздействия ионов кадмия (светлые столбики) и одновременного воздействия ионов кадмия и низкочастотного электромагнитного поля (тёмные столбики): а) микроядра; б) двуядерные эритроциты; в) амитозы; г) пузырящиеся ядра. д) лопастные ядра; е) зазубренные ядра
Если сравнивать частоты микроядер в вариантах опыта с воздействием только одного кадмия – с одной стороны и соответствующими вариантами сочетанного (совместного) воздействия кадмия и электромагнитного поля – с другой, то статистически значимые различия наблюдаются также во всех парах вариантов опытов.
Соотношение частот микроядер, индуцируемых условиями трёх серий опытов (воздействие только НЧ ЭМП, изолированное действие возрастающих концентраций ионов кадмия и мутагенный эффект воздействия обоих факторов) показано на рис. 2.
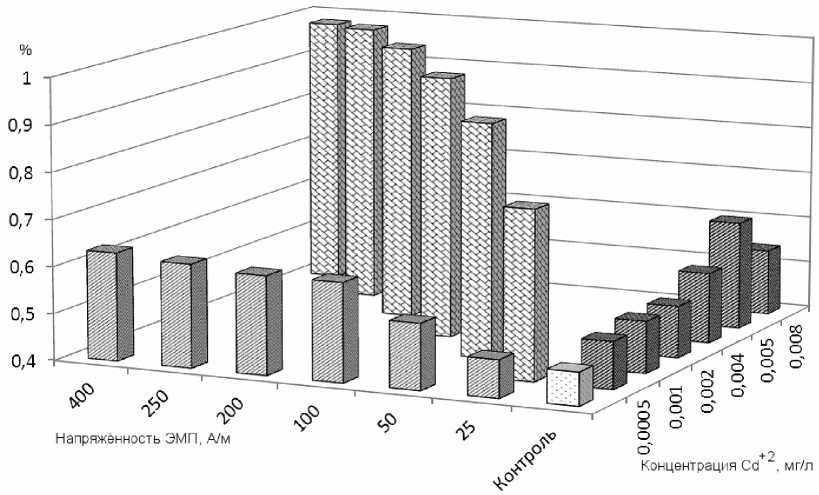
Рис. 2. Частоты эритроцитов с микроядрами у карпов после суточного воздействия на них каждого из факторов отдельно – кадмия (мг/л, правый ряд величин) и НЧ ЭМП (А/м, левый ряд), а также при сочетанном действии двух факторов (диагональный ряд) по сравнению с контрольной величиной (светлый столбик)
Частота двуядерных клеток в контроле составила 0,08%. При возрастании концентрации ионов кадмия и напряжённости электромагнитного поля просматривается тенденция увеличения частот (рис. 1, б). Небольшое снижение частоты двуядерных клеток обнаружено при максимальных значениях концентрации кадмия (0,008 мг/л) и напряжённости электромагнитного поля 400 А/м. Во всех случаях частоты при сочетанном воздействии двух факторов оказываются статистически достоверно более высокими, чем в соответствующих вариантах изолированного действия факторов.
Частоты амитотически делящихся клеток при сочетанном действии кадмия и ЭМП были выше частот амитозов, регистрируемых при действии только оного кадмия (рис. 1, в), причём в 4-х последних вариантах эксперимента (при концентрациях кадмия 0,002-0,008 мг/л и напряжённостях ЭИП 100-400 А/м) различия были статистически достоверными.
Частоты пузырящихся ядер в эритроцитах карпа после суточного сочетанного воздействия ионов кадмия и НЧ ЭМП были выше частот этих аномалий, установленных в вариантах с воздействием только ионов кадмия, однако в первых двух вариантах сочетания факторов (0,0005 мг/л + 25 А/м и 0,001 мг/л + 50 А/м) увеличение частот аномалий не было статистически достоверным. Два следующих варианта сочетанного воздействия исследуемых факторов (0,002 мг/л + 100 А/м и 0,004 мг/л + 200 А/м), а также последний вариант с максимальной концентрацией кад- мия и наибольшей напряжённостью ЭМП (0,008 мг/л и 400 А/м) приводили с статистически достоверному росту частот пузырящихся ядер. Значительно более низкая частота пузырящихся ядер при сочетанном воздействии электромагнитного поля напряжённостью 250 А/м и ионов кадмия в концентрации 0,005 мг/л нарушает общую тенденцию роста аномалий и пока трудно объяснима.
Изменение частот лопастных ядер в эритроцитах карпа, образующихся при сочетанном действии кадмия и НЧ ЭМП по мере роста интенсивности воздействующих факторов, было неравномерным. Повышения частоты при одних значениях двух действующих факторов чередовались с резкими её снижениями. Причина этого явления может быть связана с особенностью воздействия электромагнитного поля на биологические процессы в эритроцитах. Статистически достоверное увеличение частот лопастных ядер при сочетанном действии кадмия и ЭМП по сравнению с частотами, индуцируемыми одним только кадмием, установлены при концентрациях 0,002, 0,005 и 0,008 мг/л.
Частоты зазубренных ядер в эритроцитах карпа, индуцируемых при сочетанном действии кадмия и НЧ ЭМП, при малых интенсивностях воздействующих факторов были выше, чем в аналогичных вариантах воздействия только одного кадмия. Однако при напряжённости ЭМП 200 и 250 А/м происходит статистически достоверное снижение частот этих аномалий по сравнению с частотами, индуцируемыми толь- ко одним кадмием. Последующее увеличение напряжённости электромагнитного поля до 400 А/м приводит к новому снижению частоты зазубренных ядер. Однако она остаётся несколько выше, чем при изолированном воздействии одного кадмия.
Изменение частот некоторых аномалий с ростом концентрации кадмия и напряжённости НЧ ЭМП объяснить достаточно сложно. Более информативной может быть динамика суммарных частот всех шести анализируемых ядерных аномалий. Она показана на рис. 3. Как видно на графике, в обоих случаях заметна общая тенденция роста суммарных частот ядерных аномалий от контрольной величины (0,75%) до величин, обнаруживаемых при концентрации 0,005мг/л. В этом диапазоне, вероятно, преобладают механизмы, влияющие на генетические структуры клетки. Увеличение же концентрации ионов кадмия до 0,008 мг/л, вероятно, превышает определённую способность клетки к гомеостазу. Интоксикация кадмием приостанавливает внутриядерные процессы, что отражается на частоте возникновения аномалий - эта величина несколько снижается.
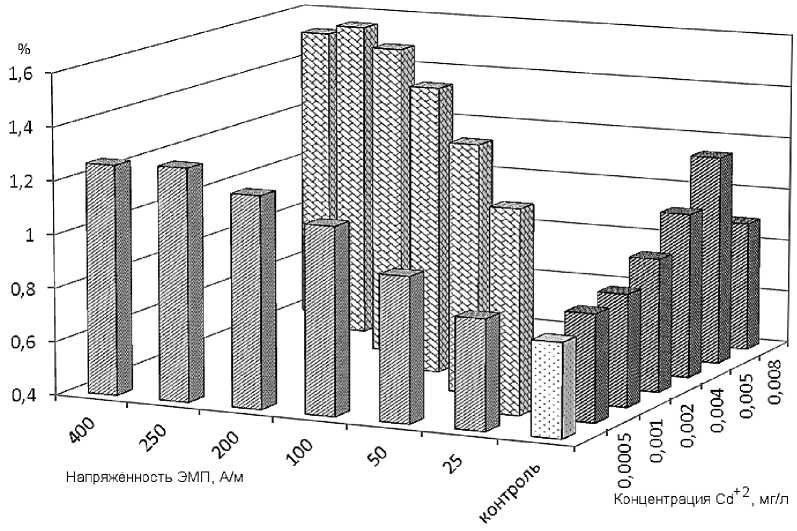
Рис. 3. Суммарные частоты всех ядерных аномалий у карпов после суточного воздействия на них каждого из факторов отдельно - кадмия (правый ряд величин, мг/л) и НЧ ЭМП (левый ряд, А/м), а также при сочетанном действии двух факторов (диагональный ряд) по сравнению с контрольной величиной (светлый столбик)
Сравнивая результаты индукции кадмием ядер-ных аномалий у рыб, полученные разными авторами, следует иметь в виду ряд обстоятельств. Интенсивность мутагенеза, индуцируемого кадмием, вероятно, будет зависеть от путей его экспериментального введения в организм - с кормом, внутрибрюшинной инъекцией или через воду, в которой находятся ионы кадмия. Скорость наступления отклика на воздействие также может различаться. В ряде исследований было показано, что максимальная частота нарушений возникает через 48 часов после начала воздействия кадмия [6]. В другом исследовании [7] максимальные частоты микроядер и сопутствующих ядерных нарушений после воздействия сублетальных концентраций ионов кадмия (0,5 и 1,0 мг/л) наблюдали на 4 сутки после начала эксперимента. После обобщения данных нескольких других публикаций, был сделан вывод, что пик частот индуцированных микроядер в эритроцитах рыб возникает в период от 1 до 5 суток после воздействия ионов металлов, но у большинства видов это происходит через 2 или 3 суток. При этом время, необходимое для достижения пика частот микроядер в периферической крови, у разных видов костистых рыб может различаться [8, 9].
К настоящему времени исследований генетических эффектов ионов кадмия, воздействующих на организм одновременно с другими химическими веществами или физическими факторами опубликовано очень немного. Было установлено, что у сои присутствие в клетках ионов кобальта уменьшает вызванную кадмием индукцию экспрессии генов MAPKK2, DOF1, bZIP62 и увеличивает зависимую от кадмия индукцию гена NR [10, 11]. Одновременное воздействие ионов кадмия (4,63 мг/л) и цинка (7,50 мг/л) в течение 24-96 часов приводило к уменьшению частоты микроядер и сопутствующих ядерных аномалий в эритроцитах тиляпии ( Oreochromis niloticus ) по сравнению с действием этих веществ изолированно друг от друга. На этом основании авторы сделали вывод о наличии антагонистических взаимодействий этих двух химических факторов [6].
Заключение
Результаты нашего исследования показали, что сочетанное действие ионов кадмия и низкочастотного электромагнитного поля усиливает повреждающие эффекты кадмия. Это изменение частот в ряде случаев оказывается статистически значимым. Биохимические 7
и биофизические механизмы этого эффекта остаются пока непонятными и требуют дальнейшего изучения.
Учитывая, что современная среда обитания растений, животных и человека содержит множество мутагенных веществ и подвергается воздействию электромагнитных полей различных частот, актуальность моделирования одновременного воздействия на геномы организмов нескольких факторов будет возрастать, и исследования в этом направлении необходимо интенсифицировать.
Кадмий, являющийся одним из тяжёлых металлов, может накапливаться в объектах окружающей среды и в живых организмах. Попадая в живые организмы, кадмий может оказывать токсическое воздействие на биохимические процессы, модифицируя активность ряда ферментов, играющих важную роль в метаболизме. Образуя комплексы с ДНК, кадмий может нарушать успешность репарационных процессов, а также изменять активность некоторых генов. Эти свойства кадмия обусловливают его мутагенные и канцерогенные свойства. Однако указанные механиз- мы ещё недостаточно ясны и для получения возможности управления ими необходимы дальнейшие исследования.
Выводы.
-
1. Одновременное воздействие на карпов ионов кадмия и низкочастотного электромагнитного поля промышленной (50 Гц) частоты вызывает статистически значимое увеличение частоты микроядер и суммарной частоты всех анализируемых ядерных аномалий в эритроцитах периферической крови по сравнению с интактными рыбами контрольной группы, а также рыбами, подвергнутыми действию только одного из двух факторов равной интенсивности.
-
2. Сочетанное воздействие на эритроциты рыб кадмия и низкочастотного электромагнитного поля оказывает более сильный мутагенный эффект, чем сумма эффектов этих факторов, действующих отдельно.
Список литературы Частоты ядерных аномалий в эритроцитах карпа при воздействии кадмия в сочетании с низкочастотным электромагнитным полем
- Arita A., Costa M. Epigenetics in metal carcinogenesis: nickel, arsenic, chromium and cadmium // Metal-lomics. 2009. V 1. -P. 222-228.
- Крюков В.И и др. Индукция микроядер в эритроцитах карпа низкочастотным электромагнитным полем / Крюков В.И., Климов А.Л., Красова Н.В. // «Состояние и пути развития аквакультуры в Российской Федерации в свете импортозамеще-ния и обеспечения продовольственной безопасности страны»: Матер. национальн. научно-практ. конф., Саратов, 4-5 окт. 2016 г. / Под ред. А.В. Молчанова. - Саратов: Изд. «Научная книга», 2016. - 152 с. -С. 60-66.
- Крюков В.И. Влияние ионов кадмия на индукцию ядерных аномалий в эритроцитах карпов // Биология в сельском хозяйстве. 2019. №1 (22). -С. 11-17.
- Урбах, В.Ю. Статистический анализ в биологических и медицинских исследованиях. М.: Медицина, 1975. 295 с.
- Абдуллаева Н.М., Гафурова С.С. Анализ эритроидных клеток карповых рыб (Cyprinus caprio L.) на основе микроядерного тестирования // Известия Самарского научного центра РАН, 2016. Т. 18, № 2. -С. 263-266.
- Abu Bakar S. N. N. et al. Genotoxic effect of zinc and cadmium following single and binary mixture exposures in tilapia (Oreochromis niloticus) using micro-nucleus test / S.N.N. Abu Bakar, A. Ashriya, A.S. Shuib and S.A. Razak // Sains Malaysiana. 2014. V. 43. № 7. -P. 1053-1059.
- Ozkan F. et al. Induction of micronuclei and other nuclear abnormalities in peripheral erythrocytes of Nile tilapia, Oreochromis niloticus, following exposure to sub-lethal cadmium doses / Ferbal Ozkan, Suna Gul Gunduz, Mehmet Berkoz, Arzu Ozluer Hunt // Turk. J. Zool. 2011. V. 35. № 4. -P. 585-592.
- Udroiu I. The micronucleus test in piscine erythrocytes. // Aquat. Toxicol., 2006. V. 79. № 2. -P. 201204.
- Udroiu I. The micronucleus test for aquatic toxicology // In book: Aquatic Toxicology Research Focus, Editors: Svensson E.P. Chapter: 7. Publisher: Nova Science Publishers, 2008. -P.145-160.
- Chmielowska-Bqk J. et al. Short term signaling responses in roots of young soybean seedlings exposed to cadmium stress. / Chmielowska-B^k J, Lefevre I, Lutts S, Deckert J. // J. Plant. Physiol. 2013. V. 170. № 18. -P. 1585-1594.
- Chmielowska-B^k J. et al. Effect of cobalt chloride on soybean seedlings subjected to cadmium stress / Jagna Chmielowska-B^k, Isabelle Lefevre, Stanley Lutts, Agata Kulik, Joanna Deckert // Acta Societatis Botanicorum Polonica. 2014. V. 83. № 3. -P. 201207.
