Чернолобый сорокопут и кобчик в фрагментах древесных насаждений полупустынного Заволжья

Автор: Сухолозов Е.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-1 т.14, 2012 года.
Бесплатный доступ
В статье рассматривается влияние двух типов древесно-кустарниковых фрагментов (линейных и компактных) на гнездование птиц. В качестве модельных выбраны чернолобый сорокопут и кобчик. Для каждого вида показаны изменения минимальных расстояний между гнёздами. Также отмечено влияние типа фрагмента на характер пространственного распределения и внутривидовые социальные связи.
Кобчик, чернолобый сорокопут, фрагменты древесной растительности, полупустыня
Короткий адрес: https://sciup.org/148205575
IDR: 148205575 | УДК: [598.279.24+598.292.2]+591.9(470.4)
Lesser Grey shrike and red-footed falcon in wood fragments of semidesrt of Trans-Volga region
The article examines the impact of two types of wood fragments (linear and compact) for bird nesting. As model Lesser grey shrike and Red-footed falcon are chosen. The changes in the minimum distances between nests are shown for each species. The influence of the fragment type on the spatial distribution and intraspecific social relationships is also noted.
Текст научной статьи Чернолобый сорокопут и кобчик в фрагментах древесных насаждений полупустынного Заволжья
Активная деятельность человека привела к преобразованию окружающей среды. В результате хозяйственной деятельности человека стали появляться принципиально новые биотопы. Вырубка естественных лесов, а в дальнейшем создание искусственных насаждений привели к появлению фрагментов и микрофрагментов древеснокустарниковой растительности. К ним относят небольшие по площади насаждения, окруженные обширными площадями других биотопов. В результате формируются изоляты – экологические сообщества с более или менее обозначенными границами [16]. Итогом фрагментации является, как правило, обеднение видового состава, а также изменение численности и плотности птиц в сравнении с лесными массивами. Также показаны зависимости этих показателей от площади фрагмента [10]. Существуют и данные об изменении поведения птиц в изолятах, в частности, указывается на снижение песенной активности и уровня агрессии [7], но таких работ мало. Однако микрофрагменты считаются значимыми местообитаниями [6]. Важность древесно-кустарниковых насаждений в пределах глинистой полупустыни волгоградского Заволжья определяется гнездованием птиц степного и полупустынного комплексов, а также использование фрагментов в качестве мест зимовок и кормёжки [3]. Этим определяется актуальность дальнейших исследований характера использования птицами фрагментов древеснокустарниковой растительности, как реакции на фрагментирование мест обитаний.
Исследования проводились в глинистой полупустыне Заволжья. Преобладающая растительность – пустынные ассоциации и разнотравнозлаковые ассоциации степного типа. Естественная древесно-кустарниковая растительность почти отсутствует или сохраняется в виде отдельных деревьев и небольших кустарниковых зарослей, в
основном приуроченных к понижениям и балкам. Искусственные насаждения представлены плодовыми садами и различными лесополосами [5]. Именно искусственные посадки мы рассматривали как фрагменты древесно-кустарниковой растительности. Внимание уделялось анализу характера пространственного распределения модельных видов птиц во фрагментах древесной растительности, как способу оценки использования пространства птицами в гнездовой период.
МАТЕРИАЛЫ И МЕТОДЫ
Сбор материала проводился в мае-июле 2007 – 2010 гг. Нами были исследованы два типа фрагментов: линейные, к которым отнесли лесополосы, и нелинейные, или компактные, а именно различные по возрасту и структуре сады. Все изученные фрагменты расположены в северозападной части глинистой полупустыни Заволжья, в Палласовском, Николаевском и Быковском районах Волгоградской области.
Лесополосы в районе исследований представлены придорожными полосами, реже полезащитными. Были исследованы придорожные лесополосы у трасс Катричев – Солдатско-Степное, Гормаки – Прудентов, Комсомольский – Вишнёвка и несколько полезащитных, расположенных между Катричевым и Солдатско-Степным. Общая протяженность исследованных лесополос 19,8 км. Из древесных пород это в основном вяз мелколо-стный ( Ulmus pumila ), клён ясенелистный ( Acer negundo ), ясень зелёный ( Fraxinus lanceolata ).
Придорожные лесополосы шириной 12-15 м состоят из нескольких (5-7) рядов деревьев, как правило, без кустарников. Частично полосы усохли, поэтому в некоторых местах разреженность достаточно значительная. Полезащитные лесополосы это посадки в виде двух лент, как правило, по одному ряду в каждой. Расстояние между лентами 5-7 м.
Исследованные сады можно разделить на две группы. К первой относятся сады у поселков Рассвет и Тракторострой, которые состоят из плодовых культур ]яблоня (Malus sp.), единично груша (Pyrus sp.)] (см. рис. 1А). Деревья образуют за- метные линии посадки. Расстояние между деревьями 3 и более метров. Вырубки составляют примерно 50% в обоих садах, из-за чего внутри сада образуются небольшие поляны. Дуб череш-чатый (Quercus robur), тополь чёрный (Populus nigra), белая акация (Robinia pseudoacacia) встречаются единично на краю садов. У поселка Рассвет имеются остатки вязовых насаждений в виде ленты. Деревья в ленте высохли, но дали прикорневую поросль. Травянистый ярус в обоих садах представлен степными растениями, преимущественно полынью Лерха (Artemisia lercheana.). Создание этих садов осуществлялось в середине ХХ в. [5].
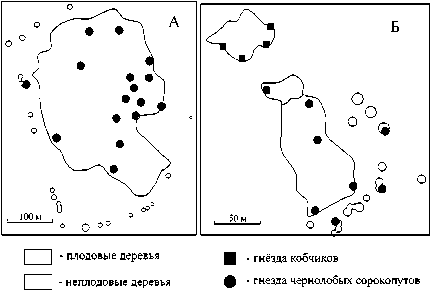
Рис. 1. Карты-схемы расположения гнёзд модельных видов в садах у пос. Рассвет (А) и Ворошилов (Б) в 2009 г.
Ко второй группе были отнесены Ворошилов и Емельянов сады (рис. 1Б). В их структуре можно выделить две резко отличающие части. Первая состоит в основном из плодовых деревьев (яблони, реже груши), которые образуют сплошной массив с небольшими участками усыхания. В травянистом ярусе преобладают сорные виды: лопух большой ( Arcticum lappa ), конопля сорная ( Cannabis sativa ), маревые (Chenopodiaceae). Вторая более разрежена. Здесь произрастают тополь чёрный, вяз мелколистный, боярышник ( Crataegus sp . ), единично – груши. Травянистый ярус не отличается от окружающей степи: преобладают полынь белая, типчак ( Festuca valesiaca ). Время создания таких садов относят к концу XIX – началу ХХ в. [15].
Все насаждения качественно отличаются типом растительности от окружающих значительных площадей степных и полупустынных ассоциаций, имеют более или менее выраженные границы. Именно это даёт нам повод к рассмотрению садов и лесополос как фрагментов древеснокустарниковых насаждений.
Сады исследовались по всей площади. Их площадь колеблется от 0,8 до 8 га. Протяжённость лесополос от 600 м до 10 км, а площадь при ширине 12-15 м – 0,4-10 га. Однако в соответствии с выбранным методом обработки результатов приходилось учитывать только определённую часть лесополос. Длина таких участков – от 600 м до 1 км, площадь – от 0,4 до 1,5 га. В среднем площадь всех изученных фрагментов, в том числе и отрезков лесополос 5,3±1,5 га.
В качестве модельных видов были выбраны чернолобый сорокопут ( Lanius minor ) и кобчик ( Falco vespertinus ). Оба эти вида обычны для региона [2]. Гнездование их приурочено к деревьям, корм собирают с открытых пространств [9]. Сорокопут – типичный энтомофаг; основную долю рациона кобчика в период размножения также составляют насекомые [13, 14].
По топографии расположения гнездящихся пар выделяли одиночно гнездящиеся пары и поселения. Под последними мы понимаем совокупность гнездящихся пар, живущих в ближайшем соседстве и непосредственно связанных друг с другом теми или иными формами сигнализации [11]. В качестве синонимов «поселения» в настоящей статье используются термины «группировка» и «скопление».
Все найденные гнёзда картировались при помощи GPS-навигатора. Координаты переносились на карту для дальнейшей обработки. Для каждого гнезда определялось минимальное расстояние до ближайшего соседа. Сравнение этих показателей осуществлялось с помощью непараметрического U-критерия Уилкоксона – Манна-Уитни.
Через характер пространственного распределения (ХПР) могут быть отражены характер использования ресурсов местообитания и внутривидовые отношения. Действие ХПР хорошо заметно на значительных по площади или протяжённости биотопах. В случае однородности среды и достаточной численности равноудалённых друг от друга особей ХПР оценивается как равномерный. При таком распределении максимально эффективно используется ресурс среды при минимальных конкурентных затратах [19]. Однако такой ХПР встречается в природе крайне редко, в том числе и по причине неоднородности среды.
Естественно, что на всей протяженности лесополос птицы встречаются не повсеместно, а сконцентрированы в определённых частях. Чернолобый сорокопут занимает участки лесополос, которые в соответствии с биологией вида окружены участками низкой степной растительности. Локализация кобчиков зависит от наличия достаточного количества пустующих гнёзд врановых, которые в местах исследований представлены в основном грачами (Corvus frugilegus), реже – сорокой (Pica pica). Стоит отметить, что в различных насаждениях глинистой полупустыни имеется избыточное количество пустующих гнёзд врано-вых, что, как мы думаем, позволяет кобчикам занимать территории в соответствии с индивидуальными предпочтениями. Таким образом, при определении ХПР для этих видов на всём протяжении даже одной лесополосы получим групповое распределение, что приведёт, как нам кажется, к неправильной оценке. Следуя этой же логи- ке, в нелинейных фрагментах, в нашем случае – садах, распределение должно быть либо равномерным, либо случайным, что также даёт искажение реальных процессов.
На небольших площадях и участках линейных биотопов ХПР легче оценить методом «ближайшего соседа», который предложили Кларк (Clark) и Эванс (Evans) и который был доработан С.П. Харитоновым [17]. Метод предполагает оценку фактических минимальных расстояний относительно ожидаемых в зависимости от занимаемой поселением площади в случае компактного размещения объектов или протяжённости в случае линейного размещения. В садах ХПР определялся для всех гнездящихся особей каждого вида, в лесополосах – только для поселений.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Всего было проанализировано расположение 250 гнёзд чернолобого сорокопута и 78 – кобчика.
В лесополосах кобчики гнездятся изолированными парами и образуют визуально определяемые скопления из 3-8 гнёзд. В среднем минимальное расстояние в таких поселениях составило 180,8±40,8 м (n = 31). Одиночные гнёзда удалены друг от друга и от скоплений, как и скопления друг от друга, на 600 и более метров. В садах одиночных гнёзд не отмечено, только групповые поселения из 3-6 гнёзд. Минимальное расстояние между гнездящимися парами в садах в среднем составляет 44,6±7,6 м (n = 8) (рис. 2).
Сорокопуты, как и кобчики, в лесополосах селятся как скоплениями (3-10 гнёзд), так и одиночными парами, в садах – только скоплениями (6-16 гнёзд). В таких группировках минимальное расстояние между гнёздами сорокопутов в лесополосах в среднем составляет 57±5,7 м (n = 57), в садах – 44±4,3 м (n = 54) Расстояния между группами гнёзд, а также между группами и одиночными гнёздами, составляет более 650 м (рис. 2).
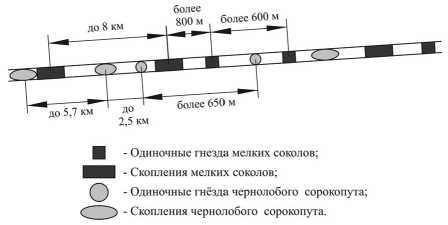
Рис. 2. Схема расположения гнёзд и скоплений чернолобых сорокопутов и кобчиков в придороджной лесополосе Гормаки-Прудентов в 2010 г.
Сравнение минимальных расстояний между гнёздами в скоплениях в различных биотопах по каждому виду показало, что в садах птицы гнездятся достоверно ближе друг к другу (для сорокопутов U = 345, для соколов U = 675; p = 0,05).
Считается, что на плотность влияет расположение фрагментированного биотопа относительно других мест [4]. Однако каждый рассматриваемый фрагмент значительно удален от любых других древесных насаждений. Даже если принять во внимание, что лесополоса имеет определённую ширину, а значит, и площадь, то зависимости плотности от площади фрагмента не выяв-ленно, так как все насаждения имеют примерно одинаковое значение площади. Получается, что минимальные расстояния, а значит, и плотность кобчика и чернолобого сорокопута во фрагментах древесной растительности в полупустыне зависит от типа насаждений. В линейных птицы гнездятся дальше друг от друга, чем в компактных.
На основе минимальных расстояний оценивался ХПР.
Для всех садов определено достоверно (р = 0,05) групповое распределение гнёзд чернолобых сорокопутов.
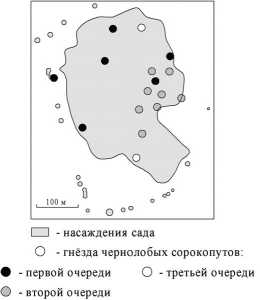
Рис. 3. Порядок заселения сада у посёлка Рассвет чернолобыми сорокопутам
По данным Е.Н. Панова [12], самки сорокопутов приступают к кладке в первые два дня после постройки гнезда, насиживание начинается, как правило, с III-V яйца. Все птицы в поселении находятся в одной стадии гнездового цикла (там же). Однако в садах было замечено изменение кладки, что естественно, и числа гнёзд. Например, в саду у посёлка Рассвет в 2009 г. при первом посещении (28 мая 2009 г.) было обнаружено 6 гнёзд (рис. 3). При втором посещении (6 июня 2009 г.) число гнёзд составило 14, в 6 из них были кладки в 1-3 яйца. В третий раз отмечалось 16 гнёзд. В гнёздах с ранее обнаруженной кладкой число яиц достигало 5. В остальных кладка либо отсутствовала, либо состояла из 1 яйца. Очевидно, что изначально сад заселялся шестью парами. Остальные заселили это насаждение позже. Подобное наблюдалось и в других садах.
ХПР гнёзд сорокопутов, которые заселили сад первыми, оценивается как случайный. Добавление гнездящихся птиц приводит к групповому распределению.
В лесополосах, к сожалению, очерёдность заселения наблюдать не удалось, зато были обна- ружены гнёзда с разной величиной кладки на основании чего, учитывая выше приведенные данные Е.Н. Панова [12], можно предположить последовательность появления пар сорокопутов в лесополосах. В качестве примера приведём отрезок придорожной лесополосы Гормаки - Прудентов в 2009 г. (рис. 4).
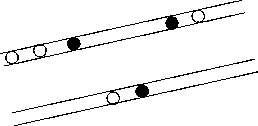
--граница насаждений
О - гнёзда чернолобых сорокопутов:
-
• - первой очереди
-
• - последующих очередей
Рис. 4. Предположительный порядок заселения отрезка придорожной лесополосы Гормаки-Прудентов
Здесь можно выделить гнёзда, которые, предположительно, заселили полосу первыми. Во время первого посещения в таких гнёздах были кладки из 3-5 яиц. В одном же были птенцы. Потом к ним примкнули другие пары.
Во всех лесополосах для скоплений чернолобого сорокопута определён случайный тип распределения гнёзд (р = 0,05).
На примере садов видно, что группировки чернолобых сорокопутов формируются по принципу первопоселенца [1]. То есть, вокруг одной или нескольких гнездящихся пар, которые первыми занимают свои территории, селятся и приступают к размножению другие пары этого же вида. Такое размещение носит контагиозный характер по терминологии Ю. Одума. Для скоплений в лесополосах подобное расположение гнёзд и образование контагиозных поселений можно предположить, но это требует уточнения.
Группировки в садах, на наш взгляд, отвечают критериям полуколонии, в лесополосах – проколонии [18], это свидетельствует о разных социальных отношениях птиц в различных типах насаждений (табл.). Это подтверждается и показателями ХПР, для скоплений сорокопутов в лесополосах, свойственны слабые социальные связи между гнездящимися парами. В садах же связи становятся более сильными. Если учесть, что отрезок лесополосы по площади сопоставим с садом, то очевидно, что сила социальных связей зависит не от площади фрагмента, а от его типа.
Особая очерёдность занятия кобчиками своих территорий нами не отмечалась. Все обнаруженные гнёзда содержали полные кладки, как правило, из 4 яиц. Для всех поселений кобчика во всех фрагментах определён случайный ХПР. На это может влиять особенность размножения соколов – они занимают гнёзда врановых. Но примечательно, что в условиях избытка гнёзд врановых характер распределения сохраняется. По неустановленным причинам два грачевника в лесополосе у трассы Гормаки-Прудентов в 2010 г. оказались не заселенными. Большая часть соколов заселила краевые участки пустующих грачевников. Правда, увеличилось число изолированных пар в сравнении с предыдущими годами. В 2010 г. их было 6, до этого 1-2. Ранее скопления соколов в этой лесополосе располагались на этих же самых местах, но в условиях присутствия грачей. Расстояния между гнёздами в 2010 г. недостоверно больше, чем в предыдущие годы. В силу увеличения расстояния между гнёздами и появлением большего числа одиночно гнездящихся пар можно было бы ожидать изменения распределения. Однако характер распределения сохранился – случайный.
Таблица 1 . Характеристика пребывания гнездящихся пар чернолобого сорокопута в различных насаждениях глинистой полупустыни
|
Признак |
Тип фрагмента |
|
|
нелинейные |
линейные |
|
|
min. расстояния между гнёздами |
меньше |
больше |
|
топография гнёзд |
поселения |
одиночные гнёзда |
|
ХПР |
случайный |
|
С одной стороны, приуроченность к местам успешного гнездования у кобчиков довольно высока [8]. С другой, увеличение расстояний между гнёздами и числа одиночно гнездящихся пар, на наш взгляд, свидетельствует о расселении птиц по лесополосе, но с сохранением ХПР. Следовательно, кобчикам свойственно сохранение социальных связей не зависимо от площади и типа фрагмента.
Часто для поселений кобчиков в литературе используется термин колония и «рыхлая» колония [8]. Но по определённому ХПР им характерны слабые социальные связи. И если использовать терминологию для неколониальных воробьинообразных, им свойственны проколониальные или полуколинальные отношения. Этот вопрос также требует уточнения.
ЗАКЛЮЧЕНИЕ
В пределах глинистой полупустыни искусственные насаждения можно рассматривать как фрагменты древесно-кустарноковой растительности двух типов: линейные (лесополосы) и нелинейные, или компактные (сады).
От типа насаждений зависят минимальные расстояния расположения гнёзд модельных видов – кобчика и чернолобого сорокопута. В линейных фрагментах гнёзда расположены достоверно дальше друг от друга, чем в нелинейных.
В лесополосах у обоих видов отмечаются одиночно гнездящиеся пары и скопления со случайным ХПР. В садах модельные виды гнездятся только скоплениями, а распределение оценивается как случайное и для кобчиков, и для сорокопутов и как групповое для сорокопутов.
Случайный ХПР характерен для животных, у которых социальные связи выражены слабо. При групповом ХПР степень социальности возрастает [19], что и наблюдалось у чернолобого сорокопута. Для всех поселений в лесополосах употребим термин проколония. В садах могут образовываться как проколонии, так и полуколонии. Следовательно, от типа фрагмента у сорокопутов зависит не только сближение особей в пространстве, но и изменение социальных отношений между особями одного вида.
Кобчик, как в садах, так и в лесополосах, демонстрирует случайное распределение, хотя и с уменьшением расстояний между гнёздами в первых. Значит, степень социальности для него постоянна, не зависимо от типа насаждений.
Таким образом, каждый вид специфично реагирует на тип фрагмента.
Список литературы Чернолобый сорокопут и кобчик в фрагментах древесных насаждений полупустынного Заволжья
- Бируля Н.Б. О структуре зооценотических группировок певчих птиц леса в сезон гнездования//Бюл. МОИП, отд. Биол. 1971, Т. LXXVI (6). С. 5-21.
- Букреев С А., Чернобай В.Ф. Птицы Приэльтонья//Биоразнообразие и проблемы природопользования в Приэльтонье. Волгоград: ПринТерра, 2006. С. 59-74.
- Быков А.В., Линдеман Г.В, Лопушков В.А. Значение древесно-кустарниковой растительности для позвоночных животных Зльтонского природного парка//Биоразнообразие и проблемы природопользования в Приэльтонье. Волгоград: ПринТерра, 2006. С. 17-23.
- Головатин М.Г. К вопросу о пространственной структуре населения птиц//Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии. Мат-лы международ. конф. Казань, 2001. С. 181.
- Животные глинистой полупустыни Заволжья (конспекты фаун и экологические характеристики). М.: КМК, 2009. 164 с.
- Захарова-Кубарева Н.Ю. Лесные фрагменты как значимые гнездовые местообитания хищных птиц//III конф. по хищным птицам Восточной Европы и Северной Азии. Ставрополь: СГУ, 1998. Ч. 1. С. 36-37.
- Зубцовский Н.Е., Матанцев В.А., Тюлькин Ю.А. Некоторые итоги изучения структуры и экологии птиц малых сообществ в УАССР//Экология птиц Волжско-Уральского региона. Информ. мат-лы. Свердловск: УрО АН СССР, 1988. С. 33-35.
- Ильюх М.П. Особенности экологии кобчика в трансформированных степных экосистемах Предкавказья//Вест. ОГУ №80. 2008. С. 131-139.
- Линдеман Г.В., Лопушков В.А. Многолетние изменения видового состава и численности птиц Приэльтонья и соседних территорий//Биоразнообразие и проблемы природопользования в Приэльтонье. Волгоград: ПринТерра, 2006. С. 74-80.
- Матанцев В.А. 2004. Влияние фрагментации местообитаний на структуру населения и экологию птиц: обзор исследований//Вестн. Удм. ун-та. 2004, № 10. С. 3-38.
- Наумов Н.П. Механизмы взаимодействия популяций (на примере некоторых млекопитающих и птиц)//Зоол. журн. 1975. Т. 54, вып. 6. С. 832-847.
- Панов Е.Н. Сорокопуты (семейство Laniidae) мировой фауны. Экология, поведение, эволюция. М., 2008. С. 365-391.
- Птицы Советского Союза. Т. I. М.: Советская наука. 1951. С. 157-162.
- Птицы Советского Союза. Т. VI. М.: Советская наука. 1951. С. 44-48.
- Труды комплексной научной экспедиции по вопросам полезащитного лесоразведения. Т. 1. Вып. 1. 1951. 504 с.
- Уилкокс Б. А. Островная экология и охрана природы//Биология охраны природы. М.: Мир, 1983. С.117-140.
- Харитонов С.П. Метод «ближайшего соседа» для математической оценки распределения биологических объектов на плоскости и на линии//Вест. Нижегородского ун-та им. Н. И. Лобачевского. Сер.: Биол. 2005, № 1. С. 213-221.
- Цветков А.В. Групповые поселения птиц как способы пространственного размещения видов: Автореф. дис. … канд. биол. наук. Москва, 2004. 16 с.
- Шилов И.А. Экология: учеб. для биол. и мед. Спец. вузов. М.: Высш. шк., 2003. С. 245-248.
