Цианобактериальные цветения воды в пресноводных континентальных водоемах: обзор

Автор: Уманская М.В., Горбунов М.Ю., Тарасова Н.Г.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Гидробиология - биологические науки
Статья в выпуске: 5 т.25, 2023 года.
Бесплатный доступ
Цианобактериальные цветения пресных континентальных водоемов являются в настоящее время одним из глобальных вызовов, стоящих перед человечеством. Расширение их масштабов, увеличение частоты связано с такими факторами, как увеличение численности населения, интенсификация земледелия, рост промышленного производства увеличение потребления полезных ископаемых, и глобальными климатическими изменениями. Массовое развитие цианобактерий вызывает разобщение звеньев первичных продуцентов и консументов, избыточное накопление биомассы цианобактерий и растворимых продуктов их метаболизма и автолиза, включая биологически активные вторичные метаболиты и цианотоксины, и способствует формированию гипоксических условий в водной толще. Цианобактериальные цветения значительно снижают потребительские качества водных экосистем, в том числе, их рекреационную привлекательность, качество воды и ее пригодность для использования в питьевых и хозяйственных целях. Основной причиной чрезмерного развития фитопланктона в пресных континентальных водоемах является поступление избыточного количества биогенных элементов, в первую очередь фосфора, а также уменьшение циркуляции воды, вызывающее застойные явления и высокие температуры воздуха, приводящие к избыточному прогреву воды в течение длительного периода. Подчеркивается необходимость проведения специализированного мониторинга с целью оценки и контроля факторов, способствующих массовому развитию тех или иных видов цианобактерий. В обзоре обсуждаются основные виды возбудители цветения в пресных водоемах, особенности их развития и экологические потребности, последствия цианобактериальных цветения для водных экосистем и населения. Подчеркивается необходимость сокращения внешней биогенной нагрузки на водоемы как важнейшего условия долговременного предотвращения цианобактериальных цветений. Приведен обзор различных внутриводоемных методов восстановления и реабилитации водоемов. Поскольку их эффективность сильно зависит от лимнологических и гидробиологических характеристик водоема, то выбор тех или иных методов должен проводится на основе полной и всесторонней характеристики экосистемы реабилитируемого водоема.
Континентальные водоемы, цианобактерии, цветение воды, биогенная нагрузка, мониторинг, мероприятия по предотвращению цветений
Короткий адрес: https://sciup.org/148327978
IDR: 148327978 | УДК: 574.5 | DOI: 10.37313/1990-5378-2023-25-5-182-194
Cyanobacterial blooms in freshwater continental reservoirs: a review
Cyanobacterial blooms of continental freshwater reservoirs are currently one of the global challenges facing humanity. The main factors increasing the scale and frequency of cyanobacterial blooms are increasing human population, intensification of agriculture, growth in industrial production, increased consumption of ores and other minerals, and global climate change. Mass development of cyanobacteria causes the breakage of trophic links between primary producers and consumers, leading to the excessive accumulation of cyanobacterial biomass and soluble products of their metabolism and autolysis, including biologically active secondary metabolites and cyanotoxins, and contributes to the formation of hypoxic conditions in the water column. Cyanobacterial blooms significantly reduce the consumer qualities of aquatic ecosystems, including their recreational attractiveness, water quality and its suitability for drinking and household purposes. The main reason for the excessive development of phytoplankton in continental freshwater reservoirs is the influx of an excessive amount of nutrients, primarily phosphorus, as well as a decrease in water circulation, causing stagnation, and high air temperatures, leading to excessive heating of water for a long period. The requirement of specialized monitoring is emphasized in order to assess and control the factors leading to the mass development of certain species of cyanobacteria. The review discusses the main bloom-forming species of cyanobacteria in freshwater reservoirs, the features of their development and environmental requirements, the consequences of cyanobacterial blooms for aquatic ecosystems and the population. The necessity of reducing the external biogenic load on reservoirs as the most important condition for the long-term prevention of cyanobacterial blooms is emphasized. An overview of various intra-reservoir methods of restoration and rehabilitation of reservoirs is given. Since their effectiveness strongly depends on the limnological and hydrobiological characteristics of the reservoir, the choice of certain methods should be based on a complete and comprehensive description of the ecosystem of the rehabilitated reservoir.
Текст научной статьи Цианобактериальные цветения воды в пресноводных континентальных водоемах: обзор
др., но в пресных водоемах чаще всего происходят «цветения» цианобактерий, единственной группы бактерий, способных к оксигенному фотосинтезу. За счет многообразия комбинаций фотосинтезирующих пигментов (хлорофилл а, фикоцианин, фикоэритрин, каротиноиды) клетки цианобактерий могут быть окрещены в различные оттенки зеленого, голубого, красного, коричневого, желтого и розового, придавая соответствующий цвет воде в периоды своего массового развития [3].
ВИДЫ – ВОЗБУДИТЕЛИ ЦВЕТЕНИЯ
Цианобактерии – возбудители цветения воды – представляют собой гетерогенную группу. В функциональной классификации фитопланктона К. Рейнолдс [4] выделил три главные группы эупланктонных цианобактерий: M – колониальные хроококковые виды с газовыми вакуолями, в первую очередь виды Microcystis s.str., H – нитчатые гетероцистные (азотфик-сирующие) виды, в основном представители Anabaena s.l., Aphanizomenon s.l. и Anabaenopsis, и S – нитчатые безгетероцистные цианобактерии, очень широкая группа, типичными представителями которых являются виды родов Planktothrix, Planktolyngbya, Geitlerinema и Pseudanabaena. В дальнейшем эта классификация была дополнена и модифицирована [4-7], причем некоторые группы были разделены на несколько подгрупп с учетом экологических особенностей видов, их устойчивости и чувствительности к определенным факторам среды (табл. 1). Можно отметить, что группа K, включающая пикопланктонные колониальные виды без газовых вакуолей, почти никогда не формирует самостоятельных цветений.
Каждая из перечисленных групп имеет свои экологические потребности и различную чув-
Таблица 1. Функциональные группы цианобактерий – возбудителей цветения в пресноводных экосистемах (по: [4, 5, 7]; с учётом современных таксономических ревизий)
|
Группа |
Предпочитаемые условия |
Характерные таксоны |
Устойчивы к |
Чувствительны к |
|
S 1 |
Перемешиваемые слои в водоемах с низкой прозрачностью воды, эвтрофные водотоки |
Leptolyngbyales, Limnothrix, Pseudanabaena, Planktothrix agardhii |
Сильному световому голоданию |
Высокому водообмену ("промывке") |
|
S N |
Теплые перемешиваемые слои воды |
Raphidiopsis |
Световому голоданию, недостатку азота |
Высокому водообмену ("промывке") |
|
Z |
Глубокие перемешиваемые слои воды в водоемах с высокой прозрачностью |
одиночные пикоцианобактерии ( Prochlorococcaceae ) |
Низкому содержанию биогенов |
Световому голоданию, выеданию |
|
K |
Мелководные богатые биогенами водоемы |
колониальные пикоцианобактерии ( Anathece, Aphanocapsa, Cyanodictyon ) |
– |
Увеличению глубины еремешиваемого слоя |
|
H |
Крупные мезо- и эвтрофные водоемы (при лимитировании азотом) |
Азотфиксирующие Dolichospermum flos-aquae, Aphanizomenon, Cuspidothrix, Sphaerospermopsis |
Низкому содержанию азота и углерода |
Перемешиванию, слабой освещенности, низкой концентрации фосфора |
|
L 0 -L M -M |
Летний эпилимнион мезотрофных и эвтрофных водоемов, малые эвтрофные водоемы |
Chroococcales: Woronichinia, Snowella, Coelosphaerium, Gomphosphaerium, Chroococcus, Microcystis |
Очень низкому содержанию CO 2 (эффективная карбоангидраза), стратификации, сильной инсоляции |
Отсутствию стратификации или глубокому перемешиванию, световому голоданию, высокому водообмену ("промывке") |
|
R |
Металимнион мезотрофных стратифицированных водоемов |
Planktothrix rubescens, Planktothrix mougeotii |
Низкой освещенности, высоким градиентам условий |
Нестабильности стратификации |
ствительность к ключевым гидрологическим факторам (световому голоданию, перемешива-нию/стратификации), выеданию зоопланктоном, отношению N:P и т.д. (табл. 1), а также особенности сезонного цикла развития (табл. 2).
В зависимости от условий среды, цианобактериальные цветения неодинаковы по видовому составу и доминирующим видам [6, 8]. В частности, изменения в составе и соотношении доминирующих видов цианобактерий в пери оды их цветения происходят по мере эвтрофикации водоемов и климатических изменений. В пределах одного трофического типа на характер этих изменений оказывают влияние и такие факторы, как стратификация, глубина перемешиваемого слоя, световые условия, отношение N:P и др. [4,9-15]
Так, например, для мезотрофно-эвтрофного Куйбышевского водохранилища в последние 4050 лет характерна смена сообществ по типичной
Таблица 2. Сезонная сукцессия видов фитопланктона, вызывающих цветение пресных континентальных водоемов различного трофического статуса (по: [4,5, 7]). Жирным шрифтом отмечены таксоны цианобактерий
|
Трофический статус |
Период |
|||
|
Весенний |
Раннелетний |
Позднелетний |
||
|
Олиготрофный |
Группа A : Cyclotella, Rhizosolenia |
Группа L0 : крупные Dinophyceae, Woronichinia , Gomphosphaeria , Staurodesmus |
||
|
Мезотрофный |
Группа B : Asterionella formosa, Aulacoseira italica, Cyclotella, Synedra |
Группа E : Dinobryon, Mallomonas |
Группа Lm : крупные Dinophyceae , Chroocales s.str. |
|
|
Группа X3 : Kolliella, Schroederia, Chrysococcus , эукариотный пикопланктон |
Группа F : колониальные и пальмеллоидные Chlorophyta : Botryococcus, Sphaerocystis, Oocystis ets. |
Группа N : Asterionella, Tabellaria flocculosa, Fragillaria, Cosmarium, Desmidium |
||
|
Группа Y : крупные Cryptophyceae, мелкие Dinophyceae |
||||
|
Группа R: Planktothrix rubescens/mougeotii (в металимнионе со вспышками в пов. слое) |
||||
|
Эвтрофный |
Группа С : Asterionella, Fragillaria, Stephanodiscus |
Группа G : Eudorina, Pandorina, Volvox |
Группа H1 : Aphanizomenon, Dolichospermum , Gloeotrichia |
Группа M : Ceratium, Microcystis |
|
Группа X1 : Ankistrodesmus, Crucigenia, Scenedesmus, Tetrastrum . |
Группа X2 : жгутиконосцы: Chlamydomonal es, Chrysophyta, мелкие Cryptophyceae |
Группа P : Asterionella, Fragillaria, Aulacoseira granulata, Closterium, Staurastrum |
||
|
Группа Y : крупные Cryptophyceae, мелкие Dinophyceae |
||||
|
Гипертрофный |
Группа D : Stephanodiscus, Diatoma, Synedra Группа X1 : Ankistrodesmus, Crucigenia, Scenedesmus, Tetrastrum |
Группа J : Pediastrum, Coelastrum, Oocystis borgei |
Группа K : Aphanocapsa, Anathece, Cyanodictyon |
|
|
Группа X1 : Ankistrodesmus, Crucigenia, Scenedesmus, Tetrastrum, Chlorella ets. |
||||
|
Группа Y : крупные Cryptophyceae, мелкие Dinophyceae |
||||
|
Группа S1 : Leptolyngbyales, Pseudanabaena/Limnothrix, Planktothix aghardii/rubescens |
||||
эвтрофной схеме С(B,D)->G->H->M->P(Y). [16, 17]. В малых водоемах Самарской области можно наблюдать практически все варианты сезонной сукцессии. 18, 19]
ПОСЛЕДСТВИЯ ЦИАНОБАКТЕРИАЛЬНОГО ЦВЕТЕНИЯ ДЛЯ ПРЕСНОВОДНЫХ ЭКОСИСТЕМ И ЧЕЛОВЕКА
В умеренных масштабах интенсивное развитие одного или нескольких видов нетоксичных цианобактерий может иметь благоприятные последствия для прочих гидробионтов [20], однако массовое «цианобактериальное цветение» представляет собой глобальную угрозу функционированию пресноводных экосистем, здоровью людей и экологическим услугам [21].
Уровни и продолжительность «цветения» воды цианобактериями, их видовой и функциональный состав влияют на относительную численность основных планктонных организмов, включая нефототрофных прокариот, протистов, ракообразных и коловраток [22-23 и др.]. В последнее время все чаще проводятся исследования влияния цветения цианобактерий (и их различных видов) на структурные характеристики разных групп про- и эукариотического планктона [24-31 и др.]. По данным Xu et al. [31] уровень цветения цианобактерий в оз. Хунцзэ (Hongze, КНР) влияет на макротак-сономическую структуру планктонных микроэукариот. Авторы отмечают, что стабильность связей внутри сообщества микроэукариот в озере снижается по мере усиления цветения цианобактерий. В ряде работ исследовано влияние цианобактериальных цветений на функциональный состав эукариотического фитопланктона [26], на соотношение автотрофных и гетеротрофных компонентов планктона [32]. Лабораторные эксперименты [30] и натурные наблюдения [33, 34] показали зависимость численности и разнообразия инфузорий от видового состава цианобактерий. Исследование в одном из пригородных водоемов Champs-sur-Marne (Франция) показало, что таксономический состав сообществ свободноживу-щих бактерий во время цветения Anabaena (Dolichospermum) и Microcystis существенно различается [28]. При доминировании других видов цианобактерий состав сообществ гетеротрофных бактерий менее исследован. Хотя имеются работы по изучению состава бактерий в экспериментальных условиях на культурах разных видов цианобактерий, относящихся к родам Aphanizomenon, Planktothrix, Microcystis [35], возможность экстраполяции этих результатов на природные экосистемы неочевидна.
В результате антропогенной эвтрофикации и глобальных климатических изменений в последние десятилетия интенсивность, частота и продолжительность цианобактериальных цветений пресных континентальных водоемов значительно возрастает. Массовое развитие цианобактерий в континентальных водоемах вызывает разобщение звеньев первичных продуцентов и консументов, избыточное накопление биомассы цианобактерий и растворимых продуктов их метаболизма и автолиза, включая дурно пахнущие вещества, например, геосмин и формирование гипоксических условий в водной толще [3, 5, 15, 22, 36-39].
Широко известна способность многих видов цианобактерий синтезировать высокотоксичные вторичные метаболиты, цианотоксины (табл. 3). Не исключением в этом смысле являются и виды цианобактерий – возбудителей цветения воды [8, 22, 40, 41]. Гены синтеза токсинов (mcyE, aerA, sxtA) обнаружены в геномах многих штаммов цианобактерий, принадлежащих к рр. Microcystis, Anabaena, Planktothrix, Anabaenopsis, Oscillatoria, Phormidium, Nostoc и др. (табл. 3). Морфологически токсичные и нетоксичные штаммы обычно неразличимы, a частота их присутствия в пределах одного вида и даже одного водоема и одного сезона может широко варьировать.
Таблица 3. Цианотоксины (в скобках указано известное число изомеров и модификаций) и цианобактерии, их продуцирующие
|
Группа |
Токсины |
Цианобактерии |
|
Гепатотоксины |
Микроцистины (>100) |
Anabaena, Anabaenopsis, Nostoc, Microcystis, Planktothrix, Oscillatoria, Hapalosiphon |
|
Нодулярины (9) |
Nodularia |
|
|
Цилиндроспермопсин (3) |
Anabaena, Aphanizomenon, Cylindrospermopsis |
|
|
Нейротоксины |
Анатоксины (6) |
Anabaena, Aphanizomenon, Oscillatoria, Phormidium |
|
Сакситоксины (20) |
Anabaena, Aphanizomenon, Lyngbya, Cylindrospermopsis, Planktothrix |
|
|
Дерматотоксины и цитотоксины |
Лингбиатоксин- а (1) |
Lyngbya, Oscillatoria, Schizothrix |
|
Аплисиатоксины (2) |
В настоящее время проблема токсичных цианобактериальных цветений – одна из наиболее животрепещущих практических проблем в пресных водоемах. Эффективными инструментами для обнаружения токсигенных штаммов цианобактерий и определения потенциальной токсичности цветений являются молекулярно-генетические подходы. Для анализа фактической токсичности (т.е. наличия и концентраций цианотоксинов в воде природных водоемов) используются методы иммуноферментного анализа (ИФА), включая различные тест-системы, а также метод высокоэффективной жидкостной хрома-тографии/масс-спектрометрии [42, 43 и др.].
Изменения климата, интенсификация и глобализация хозяйственной деятельности человечества приводит к изменению ареалов распространения отдельных видов цианобактерий – возбудителей цветения. В ряде водоемов отмечаются изменения в составе комплекса доминирующих видов и функциональных групп цианобактерий. Все это вызывает не только существенные изменения структуры всего планктонного сообщества пресноводных экосистем, но и приводит к значительному ухудшению качества получаемых экосистемных услуг, в том числе, снижению качества воды и ее пригодности для использования в питьевых и хозяйственных целях.
СПОСОБЫ ПРЕДОТВРАЩЕНИЯ
ЦИАНОБАКТЕРИАЛЬНОГО ЦВЕТЕНИЯ И МЕТОДЫ БОРЬБЫ С НИМ
Общепризнанно, что первопричиной чрезмерного развития фитопланктона в водоемах является поступление избыточного количества биогенных элементов, в первую очередь фосфора. Кроме этого, вспышкам массового развития отдельных групп или видов водорослей способствует отсутствие циркуляции воды и низкая скорость течения, вызывающие застойные явления, а таже высокие температуры воздуха, приводящие к избыточному прогреву воды в течение длительного периода.
Поэтому, основным и обязательным условием сколь либо длительного предотвращения любых «цветений» фитопланктона является максимально возможное ограничение поступления биогенов с водосборной территории [44, 45]. Но снижение потока биогенов часто не дает немедленного эффекта из-за внутренней биогенной нагрузки, обусловленной запасами биогенов, накопленными в самом водоеме в виде биомассы и в донных осадках. Как правило, для быстрого прекращения «цветения», наряду с мероприятиями на водосборе, необходимы те или иные внутриводоемные мероприятия. Их проведение, однако, не может дать длительного эффек- та, если не ликвидированы или существенно не ограничены внешние источники поступления биогенов в водоем.
Для снижения внутренней биогенной нагрузки на водоем можно использовать механические воздействия, направленные на прямое изъятие накопленных биогенов. Это, прежде всего, удаление донных осадков с целью предотвращения обратного поступления накопленных в них биогенных элементов (фосфора и азота) в водную толщу [44]; скашивание и удаление макрофитов, а также удаление из поверхностных слоев водоема биомассы водорослей, в результате чего вместе с биомассой изымаются связанные в ней азот, фосфор и органические соединения.
В стратифицируемых озерах определенный эффект может дать механическая дестратификация [46], в результате которой увеличивается глубина перемешиваемого слоя, и фитопланктон начинает испытывать световое голодание. Дестратификация может проводиться либо путем откачивания придонной воды в поверхностные горизонты (часто с фонтанированием), либо, наоборот, закачкой воздуха или кислорода в придонный горизонт. Принудительная аэрация придонного слоя дополнительно приводит к окислению верхнего слоя донных отложений, что, в определенной степени, предотвращает диффузию из них фосфора за счет его связывания железом (III).
Относительно новым способом является ультразвуковое облучение водной толщи [47]. Эта обработка разрушает крупные колонии цианобактерий, разбивая их на короткие филаменты или отдельные клетки, которые, в отличие от колоний, легко поедаются нанофлагеллятами и более крупным зоопланктоном. Однако имеются данные о том, что такая обработка вызывает гибель крупного зоопланктона (дафний) [48]. В целом, большая часть данных об эффективности этого метода получена в лабораторных условиях, а в полевых условиях он остается недостаточно апробированным.
Цветения часто удается предотвратить, просто повысив коэффициент водообмена водоема путем увеличения объема поступающей в него воды с низким содержанием биогенных элементов. Такая «промывка» снижает влияние биогенной нагрузки водоема за счет увеличения выноса биогенов в составе водного стока. Однако этот способ применим только для проточных водоемов и требует наличия источника воды с низкой концентрацией биогенов [3].
Химические методы сводятся либо к снижению концентрации в воде биогенных элементов (обычно фосфора), либо к токсическому воздействию на организмы-возбудители цветения [44]. К первой группе относятся такие обработки, как гипсование, обработка солями алюминия, железа (III) или лантана, которые связывают неорганический и часть органического фосфора в нерастворимые соединения, осаждающиеся затем в донные осадки [49]. В целом, эти методы считаются достаточно надежными, однако их эффективность и длительность действия в различных водоемах варьирует в широких пределах. Ко второй группе можно отнести обработку соединениями меди (сульфат меди, и ее различные хелатные соединения), органическими гербицидами и окислителями (хлорноватистой кислотой, перекисью водорода, озоном и солями пероксикислот, например, персульфатом). Все они действуют не только на целевую группу цианобактерий, но и на большинство гидробионтов, включая беспозвоночных и простейших. Поэтому обработки любыми соединениями меди и гербицидами приводят к существенным нарушениям в экосистемах водоемов и поэтому не могут широко применяться в природных экосистемах. В последнее время в качестве альгицида предлагается перекись водорода, которая полностью разлагается в водоеме, не оставляя токсичных продуктов [50]. Применяются также пероксиды кальция и магния, механизм действия которых сочетает мягкое окислительное воздействие и иммобилизацию фосфатов в грунты [51].
Еще одна группа внутриводоемных воздействий – это биологические методы, т.н. биоремедитация или «биологическая рекультивация». Биологические методы традиционно считаются более мягкими и щадящими, в сравнении с «чужеродными» химическими, однако зачастую предполагают интродукцию новых видов, а этот тип биологических воздействий может вызывать крайне опасные экосистемные последствия и поэтому подвергается критике. Следует учитывать также, что успех биоремедитации зависит от множества факторов, причем методы, показывавшие хорошие результаты в условиях лаборатории, могут оказаться совершенно непригодными в естественной среде, как из-за значительно отличающихся условий, так и из-за не обнаруженных в эксперименте отрицательных последствий .
Наиболее широко используются методы интенсификации выедания цианобактерий организмами высших трофических уровней. Они заключается в отлове рыб-планктофагов и бентофагов и интродукции или увеличении численности рыб-хищников и крупного зоопланктона, в частности крупных дафний [52]. В результате, как ожидается, мелководное озеро, в котором вода станет прозрачной до дна на значительной части акватории, перейдет из «фитопланктонного» в «макрофитное» устойчивое состояние [53]. Такая биоманипуляция, как правило, дает хорошие первоначальные результаты в плане снижения интенсивности или полного подавления цветения воды, но долговременный переход в «макрофитное» состояние наблюдается нечасто [54].
Большая часть других биоманипуляционных методов остается пока на стадии лабораторных экспериментов или показывает свою неэффективность (или малую эффективность) при апробации в природных условиях [55-58].
Несмотря на массу предлагаемых методов, лишь немногие из них прошли реальную апробацию и показали свою эффективность в природных водоемах. Большинство предлагаемых методов пока находится на стадии лабораторных экспериментов или небольших микрокосмов. Кроме того, все они имеют ограничения, и их эффективность сильно зависит от лимнологических и гидробиологических характеристик водоема, который необходимо реабилитировать. Поэтому следует с огромной осторожностью подходить к практическому применению различных методов восстановления и реабилитации в каждом конкретном водоеме. Методы предотвращения и прекращения цветений воды цианобактериями должны разрабатываться с учетом условий в водоеме и доминирования и присутствия организмов тех или иных групп, т.е. на основе результатов систематического мониторинга водоемов. При этом, безусловно, применению любых внутриводоемных методов должно предшествовать ограничение поступления биогенных элементов с территории водосбора. Без решения этой задачи любые манипуляции в водоеме будут давать лишь очень кратковременный эффект.
МОНИТОРИНГ ЭКОЛОГИЧЕСКОГО
СОСТОЯНИЯ ВОДОЕМОВ, СВЯЗАННЫЙ С ПРОБЛЕМОЙ ИХ ЦИАНОБАКТЕРИАЛЬНОГО
ЦВЕТЕНИЯ
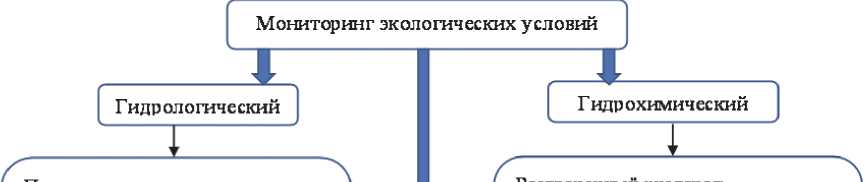
В связи с возрастающий в последние десятилетия интенсивностью, частотой и продолжительностью цианобактериальных цветений пресных континентальных водоемов насущной необходимостью является систематический и регулярный контроль состояния водоемов, направленный на получение данных о возникновении условий, способствующих цианобактериальному цветению, и отслеживание самих цветений в их динамике [3, 59]. Он должен включать в себя несколько взаимосвязанных блоков (рис.).
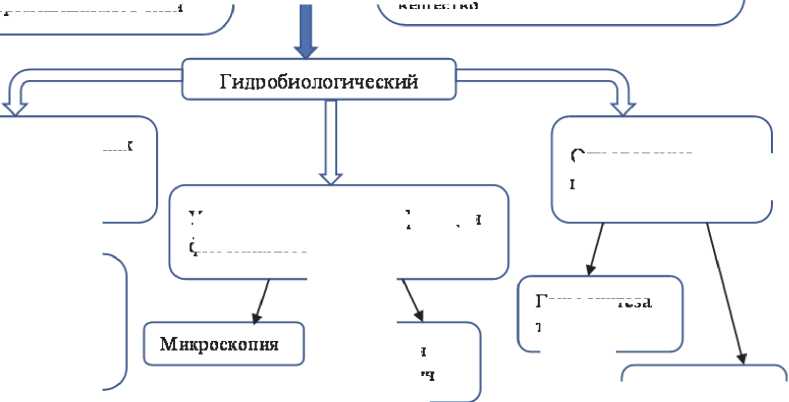
Гидрологический и гидрохимический мониторинг являются достаточно традиционными, а перечень определяемых показателей общепризнанным (рис.). Следует отметить, однако, что такой мониторинг не должен ограничиваться только поверхностным слоем, т.к. факторы возникновения цветений зависят и от состояния грунтов и глубоких слоев воды. Что касается гидробиологического мониторинга, то в аспекте отслеживания и предотвращения цианобактериальных цветений в нем можно выделить некоторые особенности.
Учет и видовую идентификацию фитопланктона можно проводить как традиционными микроскопическими методами, так и используя различные варианты проточной цитометрии [60, 61]. В настоящее время интенсивно развиваются молекулярно-генетические методы и в дальнейшем они будут находить все более широкое применение. Все эти методы имеют свои преимущества и ограничения и должны взаимно дополнять друг друга, повышая точность мониторинга [3, 59, 62]. Большие перспективы имеет использование дистанционных методов мониторинга, в частности анализ спутниковых снимков с использованием различных спектральных индексов [63]. Эти методы могут быть полезны для обнаружения пятен цветения на крупных водоемах, отслеживания сезонной динамики и других особенностей массового развития цианобактерий в разнообразных пресных водоемах [64]. Однако, для получения оптимального результата крайне желательно применять дистанционные методы одновременно с традиционным учетом цианобактерий в водоемах.
При разработке схем мониторинга необходимо обязательно включать в них определение штаммов цианобактерий, способных к синтезу токсинов, и концентрацию токсинов в воде. В России в настоящее время определение токсичности не является обязательным параметром при анализе качества воды, однако имеются данные о том, что в водохранилищах Волго-Камско-Донского каскада концентрация микроцистинов может изменяться в пределах 0,1-16,4 мкг/л в зависимости от условий, а основными их продуцентами являются виды родов Microcystis и Dolichospermum , обычные обитатели водоемов Европейской части России [65]. Данные о составе продуцентов микроцистинов и других цианотоксинов должны учитываться при прогнозировании рисков для населения от массового развития разных видов цианобактерий в поверхностных источниках питьевого водоснабжения.
Таким образом в целях развития системы раннего предупреждения о цианобактериаль-
Прозрачность
Температура
Мутность
Скорость течения
Глубина фотнческого слоя
Глубина перемешиваемого слоя
Растворенный кислород pH, Eh, электропроводность Бногены (в первую очередь, общин азот и общий фосфор)
Общее и растворенное органическое ярттт естил
Определение фотосинтетических пигментов (хлорофиллов, каротиноидов, фикоцианина)
Погружные зонды, Спектрофотометрия, Хроматография.
Анализ спутниковых снимков
Гены синтеза токсинов
Проточная цитометрия концентрации токсинов
Учет и видовая идентификация фитопланктона
Определение цианотоксичности
Рис. Блок-схема экологического мониторинга пресных водоемов, в которых наблюдаются опасные цианобактериальные цветения
ных цветениях в пресных водоемах и их пространственно-временного динамического отслеживания необходим синтез новых современных и традиционных методов мониторинга.
ЗАКЛЮЧЕНИЕ
Значительный рост населения городов и поселков на берегах водоемов, интенсификация земледелия, рост промышленного производства, а также глобальные климатические изменения угрожают стабильности внутриконти-нентальных водных экосистем. Обусловленная этим антропогенная эвтрофикация пресных водоемов, являющихся важным источником питьевой воды и просто неотъемлемой частью среды обитания человека, является одним из текущих глобальных вызовов человечеству.
Поэтому дальнейшее научное исследование процессов антропогенной эвтрофикации и вызванных ею цианобактериальных цветений пресных континентальных водоемов, способов защиты населения от его последствий и поиск путей и механизмов его предотвращения являются в настоящее время одной из необходимых и важнейших задач, стоящих перед научным сообществом и населением в целом.
Список литературы Цианобактериальные цветения воды в пресноводных континентальных водоемах: обзор
- Scientifi c Assessment of Freshwater Harmful Algal Blooms / C.B. Lopez, E.B. Jewett, Q. Dortch [и др.]. Washington, DC.: Interagency Working Group on Harmful Algal Blooms, Hypoxia, and Human Health of the Joint Subcommittee on Ocean Science and Technology, 2008. 79 p.
- Wurtsbaugh W.A., Paerl H.W., Dodds W.K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum // Wiley Interdisciplinary Reviews: Water. 2019. V. 6(5), e1373.
- Huisman J., Codd G.A., Paerl H.W., Ibelings B.W., Verspagen J.M., Visser P.M. Cyanobacterial blooms // Nature Reviews Microbiology. 2018. V. 16. № 8. P. 471-483.
- Reynolds C.S. Phytoplankton periodicity: the interactions of form, function and environmental variability// Freshwater biology. 1984. V. 14. № 2. P.111–142.
- Reynolds C.S. The ecology of phytoplankton. Cambrige: Cambrige Univ. Press, 2006. 536 p.
- Reynolds C.S., Huszar V., Kruk C., Naselli-Flores L., Melo S. Towards a functional classifi cation of the freshwater phytoplankton // Journal of plankton research. 2002. V. 24. № 5. Р. 417–428.
- Padisák J., Crossetti L. O., Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classifi cation: a critical review with updates // Hydrobiologia. 2009. V. 621. №. 1. P. 1-19.
- Jankowiak J., Hattenrath Lehmann T., Kramer B.J., Ladds M., Gobler C.J. Deciphering the effects of nitrogen, phosphorus, and temperature on cyanobacterial bloom intensifi cation, diversity, and toxicity in western Lake Erie // Limnology and oceanography. 2019. V. 64. № 3. P. 1347-1370.
- Корнева, Л. Г. Фитопланктон водохранилищ бассейна Волги [Под ред. А.И. Копылова] / Л.Г. Корнева. – Кострома: Костромской печатный дом, 2015. – 284 с.
- Корнева, Л.Г. Структура и динамика фитопланктона больших мелководных зарастающих озер (Воже и Лача, Вологодская и Архангельская области, Россия) / Л.Г. Корнева, И.В. Митропольская, Н.Н. Макаренкова, А.И. Цветков // Труды Института биологии внутренних вод им. И.Д. Папанина РАН. – 2021. – № 94 (97). – С. 17-29.
- Бабаназарова О.В. Уровень воды в мелководном высокоэвтрофном озере – фактор развития по макрофитовому или фитопланктонному типу (оз. Неро, Ярославская область) / О.В. Бабаназарова, С.И. Сиделев, С.М. Жданова, А.С. Литвинов, А.С. Овсеенко, К.П. Коровкина // Водные ресурсы. – 2018. – Т. 45. – № 6. – С. 645-655.
- Gibson C.E., Fitzsimons A.G. Periodicity and morphology of planktonic blue green algae in an unstratifi ed lake (Lough Neagh, Northern Ireland) // Int. Rev. Gesamten Hydrobiol. 1982.V.67, № 4. P. 459-476.
- Moed J.R., Hoogveld H.L. The algal periodicity in Tjeukemeer during 1968–1978 // Hydrobiologia. 1982. V. 95. P. 223-234.
- Nõges P., Adrian R., Anneville O., Arvola L., Blenckner T., George G., Jankowski T., Jarvinen M., Maberly S., Padisiik J., Straile D., Teubner K., Weyhenmeyer G. The impact of variations in the climate on seasonal dynamics of phytoplankton // The impact of climate change on European lakes. Springer Netherlands, 2010, pp. 253-274
- Ho J.C., Michalak A.M. Exploring temperature and precipitation impacts on harmful algal blooms across continental US lakes // Limnology and Oceanography. 2020. V. 65. №. 5. P. 992-1009.
- Паутова, В.Н. Продуктивность фитопланктона Куйбышевского водохранилища / В.Н. Паутова, В.И. Номоконова. – Тольятти, 1994. – 188 с.
- Тарасова, Н.Г. Состав, сезонная динамика и инвазийные виды фитопланктона Куйбышевского водохранилища: дис. ... канд. биол. наук / Тарасова Н.Г.. – Тольятти, 2005. – 146 c.
- Жариков, В.В. Протисты и бактерии озер Самарской области [Под ред. В.В. Жарикова] / В.В. Жариков, М.Ю. Горбунов, С.В. Быкова, М.В. Уманская, Н.Г. Тарасова, Т.Н. Буркова, Н.Г. Шерышева, Ю.М. Ротарь. – Тольятти: Кассандра, 2009. – 240 с.
- Шерышева, Н.Г. Экологическое состояние урбанизированного высокопродуктивного водоема (озеро Большое Васильевское) / Н.Г. Шерышева, М.В. Уманская, С.В. Быкова, М.Ю. Горбунов, Н.Г. Тарасова, Е.С. Кривина, О.В. Мухортова, Е.С. Краснова. – Тольятти, 2021. – 187 c.
- Водоросли, вызывающие “цветение” водоемов северо-запада России. – М.: Товарищество научных изданий КМК, 2006. – 367 с.
- Moal M., Pannard A., Brient L., Richard B., Chorin M., Mineaud E., Wiegand C.Is the Cyanobacterial Bloom Composition Shifting Due to Climate Forcing or Nutrient Changes? Example of a Shallow Eutrophic Reservoir // Toxins. 2021. V. 13. № 5. 351.
- Havens K.E. Cyanobacteria blooms: effects on aquatic ecosystems // Cyanobacterial harmful algal blooms: state of the science and research needs. Advances in Experimental Medicine and Biology, v. 619. Springer, New York, NY, 2008. P. 733-747.
- Moustaka-Gouni M., Sommer U. Effects of harmful blooms of large-sized and colonial cyanobacteria on aquatic food webs // Water. 2020. V. 12. №. 6. 1587.
- Bouvy M., Pagano M., Troussellier M. Effects of a cyanobacterial bloom (Cylindrospermopsis raciborskii) on bacteria and zooplankton communities in Ingazeira reservoir (northeast Brazil) // Aquatic Microbial Ecology. 2001. V. 25. №. 3. P. 215-227.
- Amorim C.A., do Nascimento Moura A. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning // Science of the Total Environment. 2021. V. 758. 143605.
- Корнева, Л.Г.Динамика морфофункциональных групп фитопланктона Рыбинского водохранилища и оценка качества его вод по индексу сообществ / Л.Г. Корнева, В.В. Соловьева // Водные ресурсы. – 2021. – Т. 48. – № 1. – С. 52-60.
- Zhao K., Wang L., You Q., Zhang J., Pang W., Wang Q. Impact of cyanobacterial bloom intensity on plankton ecosystem functioning measured by eukaryotic phytoplankton and zooplankton indicators // Ecological Indicators. 2022. V. 140. 109028.
- Louati I., Pascault N., Debroas D., Bernard C., Humbert J. F., Leloup J. Structural diversity of bacterial communities associated with bloom-forming freshwater cyanobacteria differs according to the cyanobacterial genus // PloS one. 2015. V. 10. № 11. e0140614.
- Krztoń W., Kosiba J., Pociecha A., Wilk-Woźniak E. The effect of cyanobacterial blooms on bio-and functional diversity of zooplankton communities // Biodiversity and Conservation. 2019. V. 28. №. 7. P. 1815-1835.
- Kosiba J., Wilk-Woźniak E., Krztoń W. The effect of potentially toxic cyanobacteria on ciliates (Ciliophora) // Hydrobiologia. 2019. V. 827. P. 325-335.
- Xu H., Liu W., Zhang S., Wei J., Li Y., Pei H. Cyanobacterial bloom intensities determine planktonic eukaryote community structure and stability // Science of The Total Environment. 2022. V. 838. 156637.
- Копылов, А.И. Многолетняя динамика гетеротрофного бактериопланктона в крупном эвтрофном водохранилище / А.И. Копылов, Д.Б. Косолпов, И.С. Микрякова // Биология внутренних вод. – 2020. – № 5. – С. 469-476.
- Tirjaková E., Krajčovičová K., Illyová M., Vďačný P. Interaction of ciliate communities with cyanobacterial water bloom in a shallow, hypertrophic reservoir // Acta Protozoologica. 2016. V. 55. №. 3. P. 173-188.
- Андреева, В.А. Свободноживущие инфузории Усинского залива (Куйбышевское водохранилище) в разгар цианобактериального «цветения» / В.А. Андреева, С.В. Быкова, М.В. Уманская, Н.Г. Тарасова // Известия Самарского научного центра РАН. – 2021. – Т. 23. – № 5. – С. 127-133.
- Zhu, L., Zancarini, A., Louati, I., De Cesare, S., Duval, C., Tambosco, K., Bernard C., Debroas D., Song L., Leloup J., Humbert, J. F. Bacterial communities associated with four cyanobacterial genera display structural and functional differences: evidence from an experimental approach // Frontiers in Microbiology. 2016. V. 7. 1662.
- Gromov B.V., Vepritsky A.A., Mamkaeva K.A., Voloshko L.N. A survey of toxicity of cyanobacterial blooms in Lake Ladoga and adjacent water bodies // Hydrobiologia. 1996. V. 322. № 1-3. P. 149-151.
- Корнева, Л.Г. «Цветение» воды цианобактериями (синезелеными водорослями) – реальная угроза ухудшения качества воды в водохранилищах Волги / Л.Г. Корнева, Н.М. Минеева, А.И. Копылов // Материалы докладов Всероссийской конференции «Бассейн Волги в XXI веке: Структура и функционирование экосистем водохранилищ». – Ижевск, 2012. – С.135–138.
- Корнева, Л.Г. Экология и метаболическая активность цианобактерий крупных разнотипных равнинных водохранилищ европейской части России / Л.Г. Корнева, В.В. Соловьева, С.И. Сиделев, Е.Н. Чернова, Я.В. Русских // Вопросы современной альгологии. – 2021. – № 2 (26). – С. 29-37.
- Сиделев, С.И. Обнаружение цианобактериальных токсинов в источниках водоснабжения и водопроводной воде некоторых городов России: поиск продуцентов и апробация методов удаления / С.И. Сиделев, О.В. Бабаназарова // Водные ресусы. – 2020. – Т. 47. – № 2. – С. 218-229
- Metcalf J.S., Codd G.A. Cyanotoxins // Ecology of cyanobacteria II: their diversity in space and time. Springer, Dordrecht. 2012. P. 651-675.
- Moreira C., Vasconcelos V., Antunes A. Cyanobacterial blooms: Current knowledge and new perspectives // Earth. 2022. V. 3. № 1. P. 127-135.
- Сиделев, С.И. Анализ фитопланктона Цимлянского водохранилища (Россия) на наличие цианобактериальных гепато- и нейротоксинов / С.И. Сиделев, Т.Б. Голоколенова, Е.Н. Чернова, Я.В. Русских // Микробиология. – 2015. – Т. 84. – № 6. – С. 732-742.
- Chernova E., Sidelev S., Russkikh I., Voyakina E., Zhakovskaya Z. First observation of microcystinand anatoxin-a-producing cyanobacteria in the easternmost part of the Gulf of Finland (the Baltic Sea). Toxicon. 2019. V.157. P. 18-24.
- Cooke G.D., Welch E.B., Peterson S., Nichols S.A. Restoration and management of lakes and reservoirs. Boka Raton, CRC Press, 2005. XX. 591 p.
- O’Sullivan P.E., Reynolds C.S. The Lakes Handbook. V. 2. Lake Restoration and Reabilitation. Oxford: Blackwell Publishing, 2005. 560 p.
- Visser P.M., Ibelings B.W., Bormans M., Huisman J. Artifi cial mixing to control cyanobacterial blooms: a review // Aquatic Ecology. 2016. V. 50. № 3. Р. 423-441.
- Румянцев, В.А. Воздействие ультразвука малой интенсивности на состояние токсигенных цианбактерий / В.А. Румянцев, В.Н. Рыбакин, И.В. Рудский, А.Н. Коровин // Доклады РАН. Науки о Земле. – 2021. – Т. 498. – № 1. – С. 101-104.
- Lürling M, Tolman Y. Beating the blues: Is there any music in fi ghting cyanobacteria with ultrasound? // Water research. 2014. V. 66. P. 361-73.
- Lürling M, van Oosterhout F. Case study on the effi cacy of a lanthanum-enriched clay (Phoslock®) in controlling eutrophication in Lake Het Groene Eiland (The Netherlands) // Hydrobiologia. 2013. V. 710. P.253-63.
- Huang I.S., Zimba P.V. Hydrogen peroxide, an ecofriendly remediation method for controlling Microcystis aeruginosa toxic blooms // Journal of Applied Phycology. 2020. V. 32. Р. 3133-3142.
- Keliri E., Paraskeva C., Sofokleous A. Sukenik A., Dziga D., Chernova E., Brient L., Antoniou M.G. Occurrence of a single-species cyanobacterial bloom in a lake in Cyprus: monitoring and treatment with hydrogen peroxide-releasing granules // Environ Sci Eur. 2021. V. 33. № 1. P. 1-14.
- Triest L., Stiers I., Van Onsem S. Biomanipulation as a nature-based solution to reduce cyanobacterial blooms // Aquatic ecology. 2016. V. 50. № 3. Р. 461-483.
- Scheffer M., Hosper S.H., Meijer M.L., Moss B., Jeppesen E. Alternative equilibria in shallow lakes // Trends in ecology and evolution. 1993. V. 8. № 8. Р. 275-279.
- Gulati R.D., Pires L.M.D., Van Donk E. Lake restoration studies: failures, bottlenecks and prospects of new ecotechnological measures // Limnologica. 2008. V. 38 (3–4). Р. 233-247.
- Suttle C.A. Cyanophages and their role in the ecology of cyanobacteria // The ecology of cyanobacteria. Their Diversity in Time and Space. [Eds B.A. Whitton, M. Potts]. Springer, Dordrecht, 2002. Р. 563–589.
- Singh D., Kaur G., Singh J., Satija S. Microbial Strategies for Controlling Harmful Cyanobacterial Blooms // Microbial Biotechnology: Basic Research and Applications. Environmental and Microbial Biotechnology. [Eds. J. Singh, A. Vyas, S. Wang, R. Prasad]. Springer, Singapore. 2020. P. 189-204.
- Kobayashi Y., Hodoki Y., Ohbayashi K., Okuda N., Nakano S.I. Grazing impact on the cyanobacterium Microcystis aeruginosa by the heterotrophic fl agellate Collodictyon triciliatum in an experimental pond // Limnology. 2013. V. 14. № 1. Р. 43-49.
- Mohamed Z.A., Al-Shehri A.M. Grazing on Microcystis aeruginosa and degradation of microcystins by the heterotrophic fl agellate Diphylleia rotans // Ecotoxicology and environmental safety. 2013. V. 96. Р. 48-52.
- Xu S., Lyu P., Zheng X. Yang H., Xia B., Li H., Zhang H., Ma S. Monitoring and control methods of harmful algal blooms in Chinese freshwater system: a review // Environ Sci Pollut Res. 2022. V. 29. P. 56908–56927.
- Toepel J., Wilhelm C., Meister A., Becker A., del Carmen Martinez-Ballesta M. Cytometry of freshwater phytoplankton // Methods in cell biology. 2004. V. 75. P. 375-407.
- Patel R., de Oliveira A., Newby Jr R., Chu T. Flow cytometric analysis of freshwater cyanobacteria: A case study. Water. 2019. V. 11. № 7. 1422.
- Howard M. D., Smith J., Caron D. A., Kudela R. M., Loftin K., Hayashi K., Fadness R., Fricke S., Kann J., Roethler M., Tatters A., Theroux S. Integrative monitoring strategy for marine and freshwater harmful algal blooms and toxins across the freshwater to marine continuum // Integrated Environmental Assessment and Management. 2022
- Кутявина, Т.И. Использование нормализованного относительного альгоиндекса для оценки «цветения» воды в водохранилищах Вятско-Камского предуралья / Т.И. Кутявина, В.В. Рутман, Т.Я. Ашихмина // Теоретическая и прикладная экология. – 2021. – № 3. – С. 60-65.
- Rolim S.B.A., Veettil B.K., Vieiro A.P. Kessler A.B., Gonzatti C. Remote sensing for map-ping algal blooms in freshwater lakes: a review // Environ Sci Pollut Res. 2023. P. 1-15.
- Chernova E., Sidelev S., Russkikh I., Korneva L., Solovyova V., Mineeva N., Stepanova I., Zhakovskaya Z. Spatial distribution of cyanotoxins and ratios of microcystin to biomass indicators in the reservoirs of the Volga, Kama and Don Rivers, the European part of Russia // Limnologica. 2020. V. 84. 125819.
