Циклы леммингов и колебания численности гнездящихся растительноядных птиц в тундре могут иметь одну общую причину

Автор: С.П. Харитонов
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 2605 т.35, 2026 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140314424
IDR: 140314424
Lemming cycles and fluctuations in the numbers of breeding herbivorous birds in the tundra may have a common caus
Текст статьи Циклы леммингов и колебания численности гнездящихся растительноядных птиц в тундре могут иметь одну общую причину
На обширных пространствах тундр Евразии из грызунов доминируют два вида леммингов: сибирский Lemmus sibiricus , который в североевропейских тундрах замещается норвежским Lemmus lemmus , и менее многочисленный копытный лемминг Dicrostonyx torquatus (Чернов 1980). Согласно общепринятому мнению, жизнь тундрового сообщества позвоночных в сезон размножения практически полностью зависит от обилия леммингов. Их численность, безусловно, определяет возможность размножения видов-миофагов: белой совы Nyctea scandiaca , среднего поморника Stercorarius pomarinus , зимняка Buteo lagopus и песца Alopex lagopus , поскольку лемминги являются их основным кормом.
Однако благодаря обилию леммингов удаётся успешно размножаться и многим другим птицам тундры – гусеобразным, куликам и отчасти воробьиным, поскольку в годы высокой численности леммингов основные хищники тундры – песцы – значительно меньше разоряют гнёзда птиц (Чернов 1980; Стишов 2004; Сыроечковский 2013; Emel’yanova 2000; и мн. др.). При этом по умолчанию считается, что прилетающие в тундру птицы в норме всегда стремятся размножаться. А как же иначе, ведь, согласно общепринятой эволюционной теории, каждый индивид стремится произвести как можно больше потомков. Ограничения безусловно возможны, однако считается, что эти ограничения исходят от внешних условий, которые могут препятствовать размножению птиц в конкретных местах. Есть даже монография, где с обильным применением математики разбираются закономерности связей в тундровых экосистемах, при этом основными элементами, «задающими ритм» динамики численности многих тундровых видов являются именно лемминги. Построения в этой книге на первый взгляд выглядят вполне логичными: мало в тундре корма для леммингов – мало леммингов, много корма – много леммингов, а когда много леммингов, то высока численность и других видов зверей и птиц в тундре. Книга содержит множество уравнений, правда, их весьма проблематично применить в конкретных тундровых экосистемах (Каменев и др. 2015).
Выяснено, что чёрные казарки Branta bernicla способны адекватно оценивать текущую численность песцов и не приступать к размножению в конкретном районе тундры при их большой численности (Сыро-ечковский мл. 1999). В случае гнездования в колониях вокруг гнёзд белых сов казарки регулируют дистанцию расположения своих гнёзд от гнезда хищника-покровителя в зависимости от численности песцов и даже от численности леммингов, о которой вполне можно получить представление не напрямую, а исходя из особенностей защиты своей гнездовой территории белыми совами (Харитонов и др. 2008; Kharitonov et al . 2013). Белолобые гуси Anser albifrons тоже в состоянии оценивать численность песцов и в годы особенно большой их численности гнездятся практически только под защитой хищников-покровителей, в роли которых выступают белые совы, реже сапсаны Falco peregrinus (Харитонов и др. 2009а).
Вместе с тем уже давно установлено, что у ряда видов гусеобразных в каждый конкретный год размножается лишь малая часть популяции. Это отмечено для белолобого гуся (Кречмар, Кондратьев 2006). У крас-нозобой казарки Branta ruficollis в размножении ежегодно участвует только до 38% (Винокуров 2001) или до 58% популяции (Костин 1985), в отдельные, неблагоприятные годы – лишь 4-5% (Винокуров 2001). У лебедя-шипуна Cygnus olor размножается лишь 24-36% популяции, у малого лебедя Cygnus bewickii – 34% (Bart et al . 1991). Эти факты приводятся, но нигде не обсуждаются, поскольку явно «бросают тень» на основы общепринятой эволюционной теории. Однако, по нашему мнению, эти факты нельзя игнорировать.
Материалы и методы
Исследования проводили в 2000-2007, 2012, 2014, 2016, 2018-2020 годах в рамках многолетнего мониторинга динамики численности крупных птиц арктических тундр и песцов в окрестностях бухты Медуза (73°21′ с.ш., 80°32′ в.д.) в Диксонском районе Таймырского Долгано-Ненецкого муниципального района (Харитонов 2015, рис. 1). Кроме того, использовались опубликованные данные исследований здесь в 1996-1999 и 2015-2017 годах (Головнюк и др. 2018; Tulp et al . 1997; Khomenko et al . 1999; Willems et al . 2002; Kleef et al . 2007). В 2015, 2017, 2021 и 2023 годах наблюдения проводили в долине реки Пуры в окрестностях стационара «Пура» (72°17′ с.ш., 85°45′ в.д., рис. 1), расположенного на северо-восточном краю Пуринского заказника, в настоящее время принадлежащего ФГБУ «Заповедники Таймыра». Наблюдения каждый год проводили с начала июня до конца июля.
По ходу работ производился поиск гнёзд и учёты численности гнездящихся, не гнездящихся и мигрирующих птиц всех обитающих в тундре видов. Координаты найденных гнёзд определяли при помощи GPS-навигаторов: Garmin-12 и Garmin-72. Численность леммингов оценивалась в баллах по 5-балльной шкале, причём применяли ещё и промежуточные баллы (методика оценки численности леммингов подробно описана в: Харитонов и др. 2008). Хотя балльная оценка численности леммингов смотрится довольно субъективной, ничего лучшего на данный момент не придумано. Балльная оценка, при отсутствии других возможностей, несомненно, очень полезна, да и достаточно объективна, если имеются более или менее устоявшиеся критерии, а наблюдатели имеют определённый опыт (Стишов 2004). Показано, что правильно оценить численность леммингов по какому-либо одному показателю крайне сложно, надо стараться пользоваться несколькими показателями, что на практике бывает не всегда возможно. Очень неплохую оценку можно получить, учитывая число встреченных в тундре леммингов на одного наблюдателя в день (Головнюк и др. 2018; Харитонов 2024). В оценке численности леммингов всегда есть элемент интерпретации, поскольку ни один из методов не даёт полного представления. На основании ряда показателей выводили результирующую оценку – балл численности леммингов. Численность леммингов считалась высокой в годы, когда балл был не менее 3, потому что в основном именно в эти годы гнездились белые совы и средние поморники.
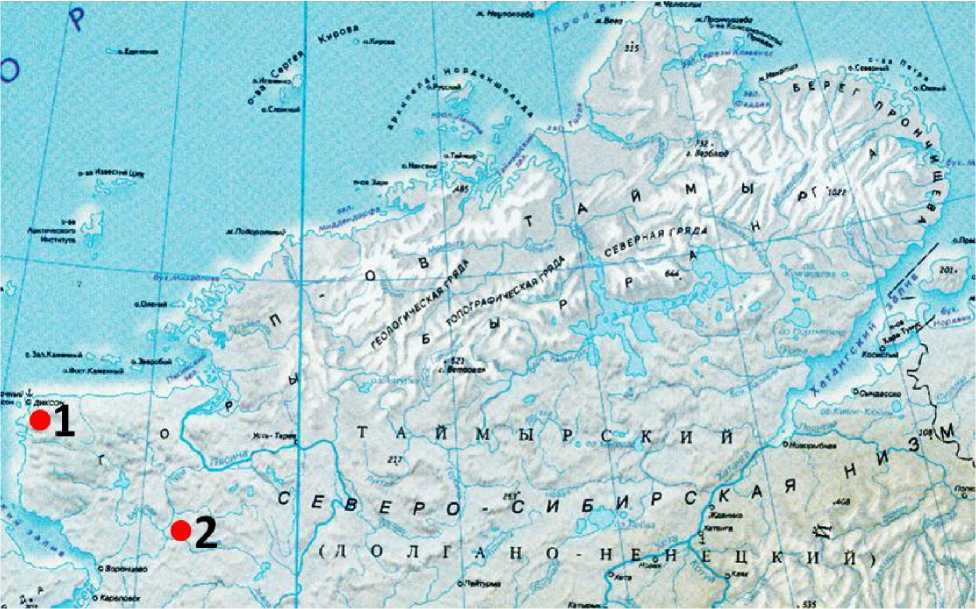
Рис. 1. Карта районов работ на полуострове Таймыр. 1 – окрестности бухты Медуза; 2 – долина реки Пуры
Площадь основной обследованной территории округ Станции имени Виллема Баренца (окрестности бухты Медузы), где базировалась экспедиция, составляла до 340 км2, хотя в первые годы работы (1996 и 1999) обследованная площадь была гораздо меньше (Tulp et al . 1997; Volkov et al . 2000). Кроме того, некоторое количество гнёзд разных видов птиц найдено южнее реки Ефремова во время дальних походов до 75 км на юг от Станции. В окрестностях стационара «Пура» обследуемая область материковой тундры составляла примерно 30 км2.
Результаты
Динамика численности белолобых гусей в окрестностях бухты Медузы и в районе работ в материковой тундре в долине реки Пуры рассмотрены нами ранее (Харитонов 2025). В упомянутом исследовании получилась довольно странная закономерность: численность белолобых гусей – растительноядного вида – зависела не от активности главного хищника тундры – песца*. Хотя в течение всего ряда рассматриваемых лет число гнёзд белолобых гусей положительно коррелировало с численностью леммингов (баллом численности леммингов), наибольшее число гнёзд белолобых гусей встречено в 2016 году при относительно небольшой численности леммингов (балл 2.5).
В долине реки Пуры во все годы нашей работы численность леммингов оценивалась баллами от 1 («очень мало») до 2 («мало»). Вместе с тем гнездящиеся белолобые гуси и гуменники Anser fabalis порой имели высокую численность (2016 и 2023 годы) на фоне очень низкой численности леммингов. В 2016 году в долине Пуры, как и в окрестностях бухты Медуза, численность леммингов только начала повышаться. А в 2023 году в районе стационара «Пура» зимой на плакорах практически не было снега, снег накопился только в оврагах, которые и стали небольшими рефугиумами для леммингов. Поэтому лемминги зимой 2022/23 года не могли повысить численность. Тем не менее по обилию гнездования птиц год был очень сходен с годами, когда численность леммингов была высокой (Харитонов 2025).
Отмечен ещё один странный факт: величина кладки белолобых гусей оказалась больше в годы с более высоким баллом численности леммингов, хотя гуси миофагами не являются (Харитонов 2025). Из чего было сделано заключение, что величина кладки свидетельствует о степени готовности белолобых гусей размножаться. И в годы с более высоким баллом численности леммингов эта готовность оказывается выше. В результате высказано предположение, что численность леммингов не обязательно является причиной этой готовности, последняя может происходить от некоего неизвестного нам фактора, который обусловливает и более высокую численность леммингов, и готовность белолобых гусей размножаться в данный год (Харитонов 2025).
Другие виды гусей также демонстрируют некие неожиданные поведенческие особенности, связанные с динамикой численности леммингов. В 2005 и 2014 годах, когда численность леммингов была очень высокой (балл 5), все гнёзда чёрных казарок находились только в колониях вокруг гнёзд белых сов в радиусе 680 м от гнезда совы, что обосновано нами как граница колоний чёрных казарок у сов (Харитонов и др. 2008). В эти же годы была велика численность песцов (Харитонов и др. 2008; Харитонов 2025). В 1996 году, хотя численность леммингов была высокой, песцов районе работ не наблюдалось вовсе (Tulp et al. 2002), и за пределами колоний без всякой защиты со стороны белых сов находилось 6 гнёзд чёрных казарок (см. таблицу). В другие годы при балле численности леммингов меньше 5 в тундре всегда имелось какое-то количество гнёзд чёрных казарок, которые предпочли гнездиться в тундре без всякой защиты со стороны белых сов.
Число гнёзд гусей вблизи гнёзд зимняков не приведено, поскольку таковые защищёнными не считаем. Дело в том, что зимняк хорошо защищает только своё гнездо, пространство вокруг гнезда он защищает даже хуже, чем средний поморник (Харитонов и др. 2017).
Численность леммингов и количество гнёзд чёрных и краснозобых казарок в материковой тундре в окрестностях бухты Медуза под защитой хищников-покровителей и без неё
|
Год |
Численность леммингов. баллы |
Число гнёзд |
||||
|
Чёрных казарок |
Краснозобых казарок |
|||||
|
Под защитой белых сов и средних поморников |
Без защиты |
Под защитой сапсанов |
Под защитой белых сов |
Без защиты |
||
|
1996* |
5.0 |
27 |
6 |
– |
0 |
0 |
|
1999* |
4.0 |
125 |
23 |
– |
0 |
0 |
|
2000 |
2.0 |
0 |
2 |
0 |
||
|
2001 |
1.0 |
2 |
6 |
0 |
||
|
2002 |
3.0 |
76 |
11 |
встречены только выводки |
||
|
2003 |
2.5 |
1 |
3 |
1 |
||
|
2004 |
3.5 |
26 |
4 |
5 |
0 |
0 |
|
2005 |
5.0 |
39 |
0 |
3 |
4 |
0 |
|
2006 |
1.0 |
0 |
4 |
0 |
||
|
2007 |
3.5 |
1 |
5 |
7 |
0 |
0 |
|
2012 |
1.0 |
0 |
1 |
0 |
||
|
2014 |
5.0 |
23 |
0 |
6 |
3 |
0 |
|
2016 |
2.5 |
0 |
4 |
2 |
2 |
9 |
|
2018 |
2.0 |
0 |
0 |
0 |
||
|
2019 |
3.0 |
20 |
0 |
2 |
0 |
1 |
|
2020 |
1.0 |
0 |
1 |
0 |
||
Примечания: * – за 1996 и 1999 годы данные получены из статей: Tulp et al . 1997; Volkov et al . 2000, Willems et al .
2002, Kleef et al . 2007. Тире – нет данных; пустые клетки в годы, когда белые совы не гнездились.
Краснозобые казарки за все годы, исключая 2016, только в два гнездовых сезона сформировали по 1 гнезду в сезон без всякой защиты, все остальные гнёзда этих гусей в эти годы были под защитой сапсанов на берегах рек Лемберова, Максимовка и Ефремова, либо под защитой белых сов, и если такие колонии были в плоской тундре, и то – совместно с чёрными казарками. В то время как в 2016 году без всякой защиты было 9 гнёзд, хотя начальная численность леммингов была 2.5 балла, как и в 2003 году, но в 2003 году она во время сезона размножения не росла, а в 2016 росла очень интенсивно. В 2016 году было очень малое разорение гнёзд белолобых гусей, а разорения гнёзд чёрных и красно-зобых казарок не отмечено вовсе.
2016 год своей необычностью поднял дополнительные вопросы. Два гнезда краснозобых казарок находились на речных островах рек Мак-симовки и Ефремова. Гнёзда на островах гораздо лучше, если не пол- ностью, защищены от песцов, чем гнезда в открытой тундре, поскольку песцы не любят переплывать реки (Е.В.Сыроечковский, устн. сообщ., собственные наблюдения автора)*. Почему краснозобые казарки не гнездились на этих речных островах во все остальные годы, хотя явно были бы с высокой вероятностью там защищены? Подобное в два года наблюдалось у белолобых гусей, которые гнездились на одном из островов реки Максимовки, а в большинстве лет на этом острове не гнездились. Получается, что есть более сильный, чем просто защищённость места, фактор, который заставляет гусей гнездиться в одни сезоны и не даёт им гнездиться в другие.
2016 год отличался тем, что тогда в тундре было хорошо всем: гуси и кулики успешно размножались (Головнюк и др. 2018), песцы размножались лишь в одной норе, но щенков было 8. Этот сезон показал, какая может быть в тундре экологическая гармония. Поскольку такой год был только один, во всех остальных кому-то было плохо, это показало, что тундра в норме является весьма несбалансированным биомом.
В 2018 году первая пара чёрных казарок совершала поисковый облёт тундры уже 9 июня. С этого времени по 27 июня включительно в тундре повсеместно наблюдались кормящиеся пары и группы до 3 пар чёрных казарок. При этом не отмечено ни одной попытки гнездования. Причина неясна, поскольку активность песцов была даже меньше, чем в 2016 году, когда дисперсные пары по тундре гнездились. С 28 июня не отмечено ни одной пары чёрных казарок в тундре.
Что касается численности леммингов в 2018 году, то тут тоже ситуация не совсем ясна. После 2016 года депрессия численности леммингов началась в 2017 году (Головнюк и др. 2018). 2018 год, казалось бы, был весьма благоприятен по погодным и кормовым условиям. Зима 2017/18 года была многоснежной, причём снег лёг на непромёрзшую землю, прикрыв норы леммингов и не заморозив их, создав таким образом хорошие условия для характерного для леммингов подснежного размножения (Чернявский, Ткачёв 1982). Обилие травы в этом году было столь высоким, что к лету тундра стала похожа на нескошенную степь. Тем не менее, подъёма численности леммингов не произошло.
Ситуации с численностью песцов в 2018 году была весьма благоприятна для гнездования птиц: численность и активность песцов в этом году были очень низкими. Видимо, 2018 год был годом одной из самых низких численностей песцов, отмечаемых в здешней тундре. Если не считать песцов у нор, то в 2018 году в тундре песцы встречены всего 5 раз: 1 июня у Станции, остальные 4 — от 5 км и далее на юг от Станции. Размножение зафиксировано лишь в одной норе в 4.7 км на юго-запад от Станции. Однако, судя по очень малому размеру «игровой площадки» у норы, выводок невелик, возможно, всего 2 щенка. Почему не отмечено бурного размножения разных видов гусей при весьма благоприятных экологических условиях, с рациональной точки зрения не ясно.
И наоборот, сильно отрицательные факторы вовсе не всегда останавливают подъём численности леммингов, если таковой начался. Конечно, катастрофические условия, как ранняя оттепель с последующим замерзанием, могут вызвать массовую гибель этих зверьков, как было весной 2000 года в окрестностях бухты Медуза, или при замерзании талых вод, например, на Чукотке (Щипанов и др. 1990). Однако это происходит не всегда. Подобная ситуация с ранней оттепелью и последующим замерзанием весной 2014 года в окрестности бухты Медуза заметно не отразилась на численности леммингов. В тот год вода залила норы леммингов, потом почти все норы оказались заткнутыми ледяными пробками. Лемминги оказались на оттаявшей поверхности тундры вовсе без нор. Это был сезон, когда удалось наблюдать массовую агрессию леммингов по отношению к идущему по тундре наблюдателю. При этом высокая численность леммингов сохранялась весь сезон.
Ещё раз подчеркнём, что начавшийся пик численности, особенно если это суперпик, случающийся раз в 11 лет (Литвин, Овсяников 1990; Харитонов и др. 2008), то его сложно остановить. В 2005 году суперпик численности леммингов был такой, что буквально преобразил поверхность тундры: в 2006 году тундру около бухты Медуза было не узнать: буквально вся поверхность земли была изрыта норами леммингов, вырытых в 2005 году. Обилие леммингов в 2005 году кормило необычно большое количество хищников, что на численности леммингов видимым образом не отражалось. Рост численности леммингов остановила (сумела остановить) только начавшаяся во второй половине июля эпизоотия (Харитонов и др. 2008). То есть кроме внешних факторов необходимо рассматривать факторы, обеспечивающие «силу» или «слабость» самой популяции леммингов в конкретную фазу цикла. Известно, что на стадии подъёма численности жизнеспособность леммингов гораздо выше, чем на спаде: лемминг, взятый на пике, живёт ощутимо дольше, чем взятый на спаде численности (Чернявский, Ткачёв 1982; А.Г.Бубличенко, устн. сообщ.). Кроме того, проявляется и так называемый эффект Читти — на подъёме численности грызуны крупнее (Chitty 1952). Тогда можно ожидать, что на подъёме численности внешние погодные факторы меньше оказывают негативное воздействие, чем на спаде численности и депрессии. Действительно, в 2014 году очень плохие весенние условия (закупоривание нор льдом) подъём численности не остановили. Хорошие усло- вия во время депрессии, например в 2018 году, не имели значения (хорошая погода во время депрессии численности леммингов не оказывает положительного влияния на численность) – они не помогут из-за внутреннего депрессивного состояния популяции леммингов.
Обсуждение
Наблюдения и литературные данные показывают, что гуси вовсе не стремятся как можно больше размножаться. Бывают годы, когда они не настроены размножаться, а бывают и такие, когда они стремятся размножаться более интенсивно, даже помещая себя в более опасные для жизни условия, чем в норме характерно для вида, как те же чёрные и краснозобые казарки в тундре без всякой защиты.
Приведённые факты гнездования гусей разных видов без защиты в тундре и отсутствие связи с популяционными циклами леммингов требует объяснений. Численность гнездящихся гусей колеблется сообразно колебаниям численности леммингов, но в отдельные годы выглядит независимой, при этом в одном случае это совпало, пусть и с малой численностью, но с ростом популяции леммингов, а в другом случае – с полным отсутствием экологических условий для роста численности этих грызунов. Увеличение величины кладки белолобых гусей в годы более высокой численности леммингов тоже входит в эту особенность гнездования гусей.
Большое число гнёзд чёрных и краснозобых казарок без всякой защиты и большая успешность гнездования, которая имел место в 2016 году, позволяет ещё раз подтвердить, что краснозобые казарки, как и чёрные казарки, белолобые гуси и гуменники в состоянии оценить численность песцов в данном году. Однако оценка численности песца тут явно недостаточный аргумент – были и другие годы, когда песцов мало, а краснозобые казарки без защиты не гнездились. Допустить, что они в 2016 году столь точно предвидели ситуацию, что резко изменили характер гнездования, отказавшись от защиты, вряд ли уместно. Тем более, что ситуация в самом деле так точно не может быть предсказана в принципе. В этом плане надо отметить 2002 год, когда большинство чёрных казарок гнездилось под защитой белых сов, но были и тундровые гнёзда без защиты. Всё шло хорошо до середины июля, когда в район работ произошло вторжение бродячих песцов, которые разорили окраинные гнёзда гусиных колоний вокруг гнёзд белых сов и почти все гнёзда в тундре без защиты (хотя 1 гнездо уцелело). Краснозобые казарки в этот год без защиты в тундре не гнездились. Теоретически подобное вторжение бродячих песцов могло произойти и в 2016 году.
Поскольку допустить способность к таким точным предсказаниям у гусей крайне сложно, необходимо искать фактор (побудительную причину), которая заставила краснозобых и чёрных казарок гнездиться необычным образом, да ещё и максимально увеличила численность белолобых гусей. У белолобых гусей разорения были (2 гнезда из 22), но в ближайших окрестностях бухты Медуза они не случались: все 10 гнёзд в этой части района работ выводки благополучно покинули. Не было разорено даже гнездо, расположенное в менее чем в 1800 м от репродуктивной норы песца, что совершенно необычно. Гнездо было найдено в начале периода насиживания и несколько раз проверялось до момента вылупления птенцов. Причём известно, что песцы нередко ходят по следам человека и разоряют найденные им гнёзда. Здесь такого явно не было, то есть гнёзда птиц песцов не интересовали. Понять это можно, ориентируясь на малую численность леммингов в этот год (2016). Бродячих песцов в районе работ не отмечено, а самцы-хозяева репродуктивных нор должны были носить еду самкам и щенкам. При этом песец может нести в пасти только одно яйцо гуся, а леммингов за один раз он может нести много – целую гроздь. Получается, добывать яйца птиц в сезон, когда и леммингов немного, невыгодно: всё своё время самцы песцов явно тратили на добычу леммингов.
В долине реки Пуры в 2016 и 2023 годах в материковой тундре гнездилось значительно больше белолобых гусей и гуменников, чем в другие годы. Численность леммингов в эти годы была очень низка. Поэтому сезон 2023 года мною в интервью для печатных изданий «Заповедников Таймыра» был назван «лемминговым годом без леммингов» (Стрючкова
2023). Данное явление позволило мне сделать заключение, что обильное гнездование птиц, особенно гусеобразных, характерное для сезонов с высокой численностью леммингов, обусловлено вовсе не численностью леммингов, как принято считать, а имеет некую более общую причину, которая и обусловливает подъём численности всех растительноядных видов: и леммингов, и гусеобразных птиц. 2003 год в окрестностях бухты Медуза тоже можно назвать «лемминговым годом без леммингов», зим-няков в том году было много, теоретически вполне могли бы гнездиться и белые совы (Харитонов 2024, 2026).
Как уже было предложено, объяснить это можно не тем, что именно численность леммингов влияет на численность и «желание размножаться» у гусей, а наличием некоего общего фактора, который влияет одновременно и на гусей, и на леммингов, стимулируя их к более интенсивному размножению. Что это за общий фактор, пока не ясно. Представляется, что в этом плане необходимо учесть, что и лемминги, и гуси являются растительноядными. Как один из вариантов, логично предположить, что существует агент (некое вещество или группа веществ), который усиливает стремление размножаться у столь разных групп организмов, и этот агент находится в их пище, то есть в растениях.
В этом отношении проводились исследования на грызунах и были обнаружены вещества, которые стимулируют размножение этих зверьков. Во-первых, обнаружено, что в кормовых растениях, которые поедаются леммингами, временами вырабатываются легко перевариваемые протеины с высоким содержанием азота (Selas 2024). Такие протеины появляются в растениях циклически, причиной их появления считают выбросы радиоактивного газа радона, возникающие после периода высоких весенних приливов и отливов в периоды новолуния и полнолуния. Есть также гипотеза, что усиленная ионизация, способствующая накоплению радона под снегом, происходит в конце декабря — начале января, когда Земля находится в перигелии своей орбиты. Легко перевариваемые богатые азотом протеины возникают в растениях не сразу после накопления радона, а после некоего периода ионизации, длящегося примерно 2 года (Selas 2024).
Мало того, довольно давно обнаружено и выделено из растений вещество, которое явно стимулирует размножение млекопитающих. Оно воздействует не только на леммингов, но и на полёвок разных видов и даже на кенгуру, которые наедаются свежей травы после засухи, после чего следует период бурного спаривания. Учёные называют это вещество супергормоном (Sullivan 1981). Отмечается, что в условиях северных биоценозов это вещество появляется в растениях время от времени и непредсказуемо, примерно как циклы леммингов, отчего его появление и считают причиной вспышек размножения леммингов. Это вещество — коричная кислота, в искусственно синтезированном варианте это циклический углеводород 6-метоксибензоксазолин. Результаты опубликованы в 2 статьях в журнале «Science» в 1981 году (Sullivan 1981).
Хочу ещё раз подчеркнуть, что вещества в растениях вовсе не обязательно являются причиной, стимулирующей размножение и леммингов и гусей. Эта гипотеза лишь одна из возможных. Серия исследований на эту тему проведена в прошлом веке, получить подтверждение данной гипотезы в естественных условиях очень трудно, поэтому данные работы были не столько раскритикованы, сколько оставлены без должного внимания. На мой взгляд, они незаслуженно забыты, и в современных условиях с их большими инструментальными возможностями неплохо бы вернуться к изучению этих вопросов.
Возвращаясь к обсуждению, отмечаем, что в вышеприведённых исследованиях особо указано, что такие вещества возникают в растениях циклически. Хотя в указанных работах речь идёт только о млекопитающих, синхронность бурного размножения леммингов и гусей позволяет предположить, что с большой вероятностью это вещество также стимулирует и размножение у травоядных птиц. Кроме того, возможно допустить, что этот стимулирующий размножение агент (или агенты) содержатся в растениях в разных дозах в разные годы. Поэтому эти вещества могут действовать то сильнее, то слабее. Когда его действие сильное, то те особи, у которых действие этого агента наиболее сильное, стремятся размножаться скорее. Им «не хочется» тратить время, которое неизбежно требуется на завоевания определённых мест в колониях, поскольку в там обязательно присутствует конкуренция (Kleef at al . 2007; Харитонов и др. 2009б), и они становятся «индивидуалистами» (термин из: Сапельников и др. 2006). В этом плане примечательно, что в 2016 году возле 2 гнёзд сапсана было не более 1 гнезда краснозобых казарок, тогда как в другие годы это число в окрестностях бухты Медуза достигало 3, как, например, в 2014 году, когда при высокой численности леммингов крас-нозобые казарки тем не менее в открытой тундре не гнездились. В 2016 году краснозобые казарки явно не предпочитали гнездиться возле гнёзд сапсана. Индивидуалистами в тундре были и чёрные казарки в 2016 и некоторое количество чёрных казарок в 1996, 1999, 2002, 2004 и 2007 годах, которые гнездились вне колоний, несмотря на полное отсутствие защиты от песцов. В 2016 году абсолютное число и доля краснозобых казарок, загнездившихся под защитой сапсанов, была наименьшей за все годы работ.
Получается, что в отдельные годы казаркам двух видов не хватает «терпения» дождаться завоевания места в колониях возле хищников-покровителей и они начинают гнездиться в открытой тундре, где выбор места ничем не ограничен. Тогда возникает вопрос, а почему в 2005 и 2014 годах такого терпения хватило, хотя леммингов было много, а значит и стимулирующего размножение вещества тоже должно было хва- тать? Здесь, очевидно, сработало то, что в эти годы была высока численность песцов, а гуси, как неоднократно было показано, в состоянии оценивать численность песца. Если же в условиях большой численности леммингов песцов мало или вовсе нет, то тогда это «нетерпение» вполне реализуется, как произошло в 1996, 1999, 2002, 2004, 2007 и 2016 годах.
В другие года, кроме 2016, стимулирующий размножение фактор в растениях появлялся ранней зимой, так что лемминги успевали увеличить к весне численность. А в 2016 – в год суперпика численности леммингов – это вещество появилось почему-то позже, и лемминги к началу сезона гнездования белых сов не успели увеличить численность. А вещества этого было много, поскольку оно только весной появилось, то на гусей оно сильно подействовало. В результате в этом году было наибольшее за все сезоны работы число гнёзд белолобых гусей тундре и большое число краснозобых казарок в тундре, чего до этого не было никогда. Получается так, что со временем количество этого вещества в растениях как-то уменьшается: ведь если лемминги увеличивают численность зимой, то белолобых гусей и казарок становится больше, чем в годы низкой численности леммингов.
Отдельные годы в экологическом плане воспринимаются как «есть все условия»: и корма много, и песцов мало, а белолобые гуси и чёрные казарки перемещаются по тундре группами, кормятся, но не гнездятся или гнездятся в очень малом числе (например, в 2018 году). В другие годы хотя корма и хватает, но песцов ощутимо больше, и они ещё и размножаются (2016 год), но гуси всё равно гнездятся.
Получается необычное явление: растения при помощи неких веществ (или процессы, обуславливающие появление в растениях этих веществ) управляют физиологическим состоянием и поведением позвоночных животных, определяя их репродуктивное состояние. Иными словами, некие вещества коренным образом управляют целой экосистемой. Похоже, ситуация усиливается вот чем: растения с этими веществами могут становиться более привлекательной пищей, чем растения без них. В результате растения с этими веществами задерживают птиц в местах своего произрастания, причём порой настолько, что птицы не ищут других, даже более характерных для их гнездования мест. Например, в том же 2016 году одна из пар краснозобых казарок, что гнездилась близко от базы экспедиции, была отмечена за 2 дня до начала её гнездования в тундре, и не видно было, чтобы куда-то улетала.
Модель популяционных циклов леммингов
После всего вышесказанного мне хочется представить собственную гипотезу о механизме популяционных циклов леммингов. Чтобы лучше обосновать такую гипотезу, полезно выработать специальную математическую модель. Для такой модели имеем начальное условие, что чис- ленность леммингов меняется циклически. В качестве увеличенной численности леммингов возьмём балл 3 и выше. Хотя балл 3 – это «средняя численность», именно при такой численности леммингов в норме начинают гнездиться белые совы (Харитонов 2024). Сведения о реальных колебаниях численности леммингов на Таймыре с 1960 года содержатся в статье Кокорева и Куксова (Kokorev, Kuksov 2002). Сведения о последних годах в серии циклов, приводимых этими авторами, подтверждаются тем, что совпадают со сведениями, изложенными в статье Рыхликовой и Попова (Rykhlikova, Popov 2000). Дальнейшие сведения содержатся в упомянутых статьях по окрестностям бухты Медуза, а далее – нашими наблюдениями. В результате мы имеем временной ряд с 1960 по 2020 год, лишь с небольшим перерывом в 2008-2011 годах, когда сведений о численности леммингов нет (рис. 2).
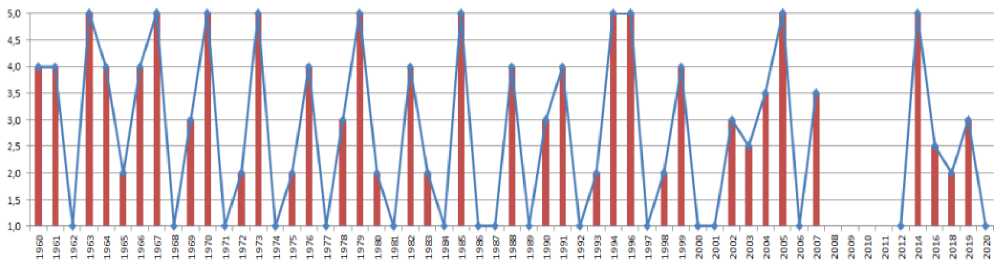
Рис. 2. Баллы численности леммингов на Таймыре в 1960-2020 годах.
Баллы представлены в виде столбиков и в виде графика. По: Kokorev, Kuksov 2002; Rykhlikova, Popov 2000 и собственным наблюдениям (Kharitonov 2024). Ось абсцисс – годы, ось ординат – баллы численности леммингов по 5-балльной шкале
Из предыдущего изложения ясно, что колебания численности леммингов часто никак не связаны с экологическими условиями. Поэтому попробуем предсказать годы с баллом численности лемминга 3 и выше на Таймыре чисто нумерологически – прибавлять к каждому имеющемуся подъёму 3 года* и смотреть, насколько это совпало с прогнозом. Если мы будем предсказывать подъёмы численности по такой схеме, то есть к каждому подъёму прибавляем 3 года и смотрим, есть ли через 3 года тоже подъём численности или нет. В результате за период 19602020 годов 18 раз прогноз сбылся, 15 раз – нет, то есть верных предсказаний 55% – более половины. В этом плане есть совпадение, подтверждающее подобного рода прогноз: если для прогноза циклических колебаний численности леммингов (возможно, и любых грызунов) берётся наиболее частый интервала между одинаковыми фазами цикла, то на материале, гораздо большем, чем наш, 649 годо-мест по всей Арктике, и принимая не балл 3, а более высокие баллы численности леммингов, которые в указанном случае следовали с интервалом 3.7 года, такой циклический интервал наблюдался именно в 55% случаев (Gauthier et al. 2024).
Теперь попытаемся понять, как формируются популяционные циклы леммингов. Известно, что в качестве возможных причин рассматривались очень многие — космические факторы, флуктуации климатических условий, циклические изменения питательных свойств растений, физиологические и эндокринные процессы в популяциях и т.д. Однако все эти факторы, как внешние по отношению к самим популяциям, так и связанные с различными внутрипопуляционными процессами, не выдержали критики в качестве единственных и основных причин наблюдаемых колебаний численности зверьков, сопровождаемых, как было установлено, и определёнными изменениями физиологического состояния популяций (Стишов 2004). Понятно, что на циклы влияет множество факторов, часть из которых мы, возможно, никогда и не узнаем. Тем не менее вопрос, на наш взгляд, поставлен неверно: необходимо не искать единственную причину (которая то ли есть, то ли нет) и рассматривать не то, что влияет, а именно то, что формирует и определяет наличие самих циклов, что создаёт сам колебательный процесс.
Для этого имеет смысл рассмотреть весьма оригинальную гипотезу, сильно отличающуюся от всех остальных, поскольку она видит в циклах леммингов чисто математическую причину (Розенберг 1987). Автор этой гипотезы рассмотрел цикличность изменения случайного ряда чисел и в средних интервалах между разными значениями чисел случайного ряда чисе л* и получил примерно такие же 3-4-фазные периодические колебания, которые наблюдаются во многих процессах: высота морских волн, солнечная активность и пр. Автору удалось получить даже те самые суперпики, которые случаются раз в 11 лет и наблюдаются в циклах солнечной активности. Хотя лемминги в статье не упоминаются, но автор говорит о всех природных циклических процессах, к которым относятся и циклы леммингов, тем более что и у леммингов имеются 3-4летние колебания численности и есть 11-летние суперпик и^.
К сожалению, автор сравнил только средние интервалы между значениями в реальных и случайных рядах, но не сравнил реальные природные распределения фаз циклов с распределением случайных чисел. И вот тут оказалась принципиальная разница. Для сверки распределе - ний при помощи генератора случайных чисел (функция RND в языке программирования Basic) нами был получен ряд случайных чисел в 1100 значений. Число 1100 было выбрано произвольно для того, чтобы в нем «содержалось» 100 11-летних суперпиков. Уже первые проверки показали, что в случайном ряду нередко наблюдаются подряд идущие несколько максимумов или долгие периоды минимумов, суперпики не имеют 11-летней периодичности, могут встречаться и подряд, могут через 1 или 2 года, чего в природе не отмечено. И само распределение интервалов между баллами численности леммингов 3 и выше в природе (имеем 24 таких интервала за период 1960-2020 годов) имеет иной характер, чем распределение интервалов между значениями в случайном ряду (рис. 3). Справедливости ради следует отметить, что сравнения наблюдаемого распределения с теоретическим в случае, если от теоретического мы используем только 24 значения, по критерию х2 получается незначимым: х2 = 23, P = 0.54. Однако следует учесть, что выборки невелики, да и недостоверное значение не означает, что выборки не различаются, а означают, что статистически вопрос остался невыясненным (Плохинский 1978).
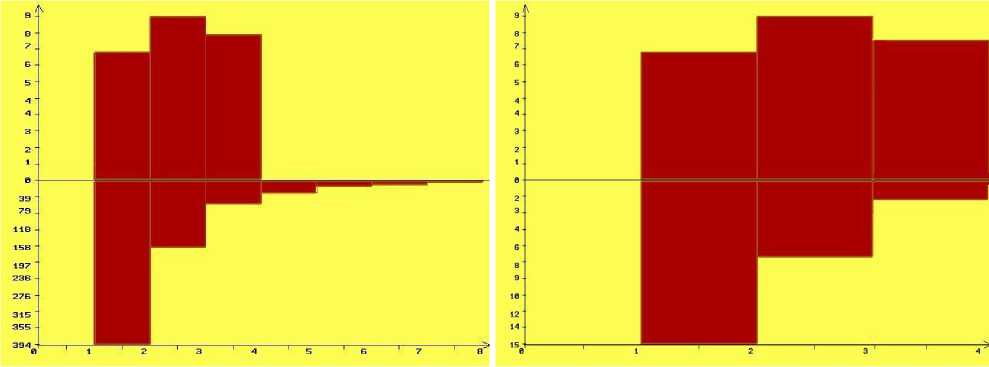
Рис. 3. Распределения интервалов между значениями численности леммингов и случайного ряда, соответствующие баллам численности леммингов 3, 4 и 5. Верхние графики – реальная природная ситуация за период 1960-2020 годов, внизу значения, построенные на основании случайного ряда: слева – использован весь случайный ряд в 1100 значений, что дало 655 интервалов между баллами численности леммингов 3 и выше, справа – взято только 24 первых значения ряда из 1100 значений, чтобы соответствовать выборке в 24 значения, что у нас имеется по природной популяции.
По оси абсцисс – интервалы (в годах) между сезонами высокой численности леммингов, по оси ординат – число интервалов
Ясно, что для описания всего процесса циклических колебаний численности леммингов чисто случайный процесс не годится: реальный процесс весьма похож на гауссово распределение с пиком в середине и меньшими значениями по краям, тогда как распределение интервалов между баллами 3 и выше в случайном ряду даёт обратную экспоненту (рис. 3). На наш взгляд, в чисто математическом подходе на основе ряда случайных чисел «не хватает биологии». Генератор случайных чисел не может
«знать», что интенсивно размножаться несколько лет подряд или с малыми интервалами между пиками лемминги не могут из-за истощения либо ресурсов (корма), либо возможностей организма для интенсивного размножения (Чернявский, Ткачёв 1982). И слишком большие перерывы между пиками размножения в норме тоже не характерны, так как лемминги, во-первых, восстанавливают силы, а во-вторых, время от времени получают возможное стимулирующее воздействие от специфических веществ в растениях.
Поскольку далеко не всегда наблюдается соответствие колебаний численности экологическим условиям, понятно, что одной биологической причины тоже недостаточно. Создаётся впечатление, что за колебания численности леммингов, кроме биологических причин, должна отвечать ещё и случайная функция. Так ли это, попробуем проверить на модели. Сформулируем основы предлагаемой здесь модели: циклы леммингов формируют в основном внутренние факторы: 1) вещества из растений, которые стимулируют размножение; 2) популяция как целостная система – есть периоды расхода энергии (размножение и рост численности), есть периоды восстановления энергии и получение стимулов из корма. Модель строится таким образом: если у нас в «текущем году» (очередном значении) балл численности леммингов получился 3 и больше, то следующее значение балла численности леммингов будет формироваться по формуле: предыдущее значение (обозначим как N i -1 ) умножить на некий коэффициент истощения (E) и умножить на значение случайной функции (RND) . Если балл численности лемминга в текущем значении 1 и 2, то это значение балла численности делится на коэффициент стимуляции (S) и делится на значение случайной функции: N i -1 : S : RND. Здесь для упрощения объяснений примем E = S по значениям.
Численные эксперименты позволили предварительно определить коэффициент истощения/стимулирования; он в нашем случае оказался равным 0.75. Кроме того, в модели необходимы верхнее и нижнее ограничения для случайного ряда. Для верхнего ограничения «численности» в случайном ряду логично принят 1, для нижнего порога (численность леммингов в природе не может быть ниже какой-то величины, иначе они вымрут) случайного ряда после численных экспериментов принято значение 0.1. Применяемый нами подход дал очень неплохое соответствие модели реальной ситуации (рис. 4). Сравнение распределений интервалов между значениями баллов численности 3 и выше в реальности и в разных выборках из нашей модели дали очень хорошее соответствие: 1) для первых 24 значений модельного ряда χ2 = 16.167, df = 23, P = 0.85; 2) для 24 значений этого ряда начиная от произвольно выбранного 40-го значения χ 2 = 13.333, df = 23, Р = 0.944; 3) для 24 значений ряда начиная от произвольно выбранного 45-го значения сравнение рядов вообще не показывает значимых различий: χ 2 = 9.5, df = 23, P = 0.994.
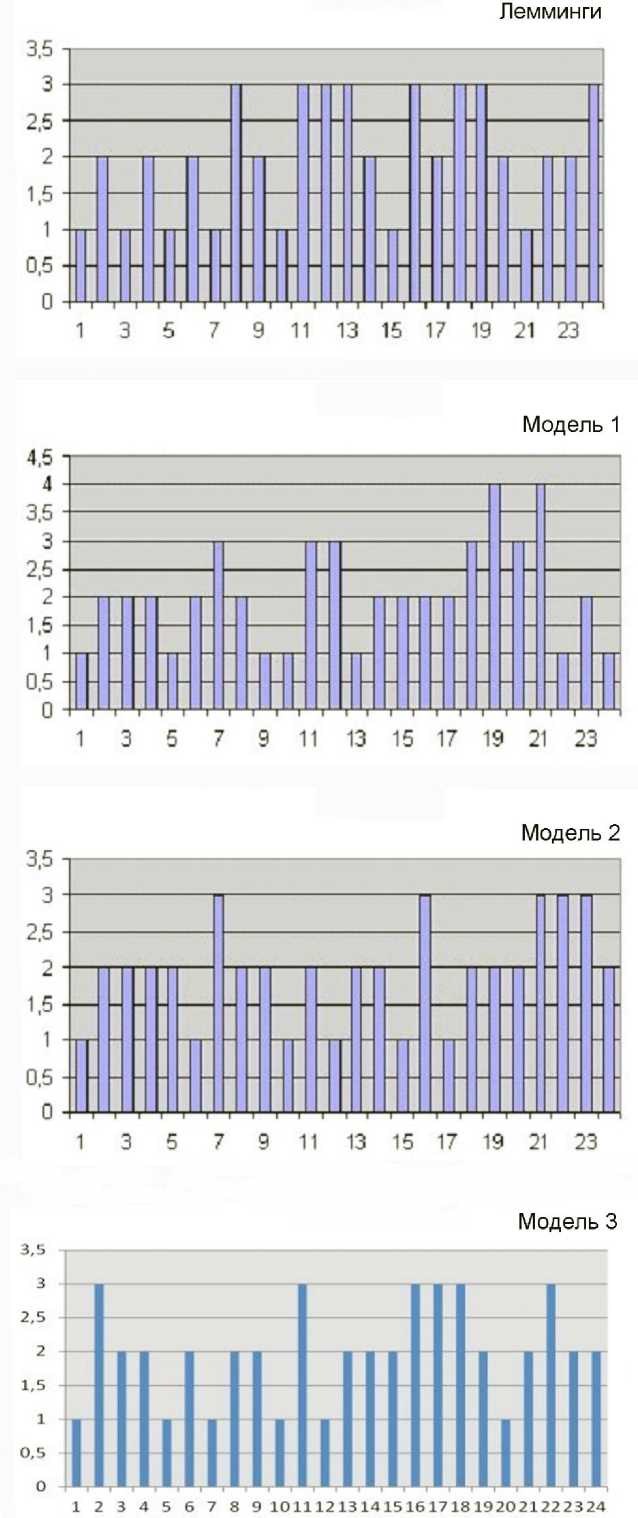
Рис. 4. Реальные и теоретические интервалы между последовательными баллами численности леммингов 3 и более. В реальности (верхний график, «лемминги»); в первых 24-х значения ряда, построенного на основе предложенной модели («модель 1»); в 24 значениях, взятых от 40-го (произвольного) значения модельного ряда («модель 2»); взятых от 45-го (произвольного) значения модельного ряда («модель 3»).
По оси абсцисс – номер интервала между высокими численностями леммингов (балл 3 и выше), по оси ординат – баллы численности леммингов
Понятно, что получены ещё только предварительные результаты, модель требует доводки. Однако надо иметь в виду, что поскольку в формировании колебательного процесса численности леммингов присутствует случайная функция, точных предсказаний во всех случаях мы не получим никогда.
Понимание сути процесса необходимо, чтобы представлять себе, что на самом деле формирует популяционные циклы леммингов. Для этого сравним долю верных предсказаний между баллами 3 и выше в естест -венном цикле численности и в циклах, полученных на основе ряда случайных чисел и циклов, полученных на основании предлагаемой модели. Если мы берём значения с первого члена случайного ряда и первого члена модельного ряда, то в случайном ряду имеем 7 из 24 верных интервалов между высокими значениями численностей (29.2%), а в модельном ряду - 12 из 24 (50%).
Теперь для сравнения возьмём произвольный начальный номер, но одинаковый в случайном и модельных рядах. Пусть это будет 40-е значение (потому 40-е, что оно следует сразу за периодом 4, которого нет в естественном ряду). Для случайного ряда получаем 9 из 24 (37.5%), для модельного — тоже 9 из 24 (37.5%). Здесь случайный и модельный ряды по своим предсказательным возможностям совпали.
А теперь проведём сравнение, например, с 45-го значения обоих рядов. Для случайного ряда будет 7 совпадений из 24 (29.2%), для модельного ряда — 13 из 24 (54.2%). Последнее значение весьма близко к 55% предсказательной точности, если использовать нумерологические предсказания подъёмов численности леммингов, просто прибавляя по 3 года к предыдущему подъёму.
Если проделать много таких сравнений, то в подавляющем большинстве случаев модель даст значительно более точные предсказания, нередко близкие к предсказаниям на основе естественного ряда подъёмов численности леммингов. При этом надо ещё принимать во внимание, что модель даёт гауссовско-подобное распределение значений, как и естественный ряд баллов численности, в отличие от полностью случайного ряда.
Поэтому мы можем утверждать, что предлагаемая модель в значительной степени верно описывает популяционные циклы леммингов. Это даёт нам в руки два следующих момента знаний о популяционных циклах леммингов: 1) такая величина, как коэффициент истощения и/или стимулирования популяции имеет биологический смысл и его ве- личина может быть уточнена в последующих исследованиях; 2) в формировании циклических колебаний принимает участие чисто случайная функция, не зависимая ни от каких внешних условий, что в принципе не даёт возможности точно предсказывать подъёмы численности леммингов.
Заключение
Принято считать, что высокая численность леммингов обусловливает не только увеличение численности хищников-миофагов, особенно птиц-миофагов, но и создаёт условия, при которых гуси разных видов тоже начинают интенсивно размножаться. В данном исследовании осуществлена попытка показать, что интенсивностью размножения растительноядных птиц, по крайней мере гусей управляют не колебания численности леммингов, а состояние растений и содержание в них особых веществ, которые и обуславливают колебания численности и леммингов, и гусей. Оказалось, что такие «настройки» размножаться или не размножаться в конкретные годы могут проявляться не только на уровне индивидуумов или видов, но и охватывать целые группы организмов, относящиеся даже к разным систематическим группам, но объединённые экологически как растительноядные консументы тундры.
Предлагаемая здесь модель механизма, формирующего популяционные циклы леммингов, выглядит перспективной и даёт возможность глубже понять эти циклы через роль разных биологических, а также не зависимых ни от каких биологических особенностей чисто случайных причин. Поднятый в статье вопрос на самом деле должен быть поставлен более широко. Дело в том, что не только с растительноядными животными, но и с хищниками-миофагами всё обстоит не так просто — у них тоже бывают разные состояния, которые могут обуславливать их размножение в неподходящих кормовых условиях и, наоборот, неразмно-жение при вполне подходящих условиях (Харитонов 2024). Эти вопросы требуют дополнительного изучения. Не исключено, что на разных фазах популяционного цикла сами лемминги могут иметь разную питательность для миофагов, например оттого, что величина тела грызунов разная на подъёме и на спаде численности, и на подъёме численности зверьки крупнее (Chitty 1952). Кроме того, сами зверьки на разных фазах динамики численности могут содержать разные вещества, а также находиться при разных состояниях эндокринной системы организма (Чернявский, Ткачёв 1982).
