Цитологические особенности мононуклеарных лейкоцитов, культивированных в присутствии полиаминов

Автор: Морозов И.А., Годовалов А.П.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Иммунология
Статья в выпуске: 3, 2023 года.
Бесплатный доступ
Рассмотрены количественные и качественные изменения в клеточном составе после инкубации с путресцином и кадаверином. Цель исследования - изучение изменений цитологических характеристик лейкоцитов после их инкубации с кадаверином и путресцином. Оценивали количество, типы клеток, размеры, наличие вакуолей и включений у лейкоцитов периферической крови (n = 15) после их предынкубации с кадаверином (5 и 25 ммоль/л) и путресцином (5 и 25 ммоль/л) в течение 72 ч при 37ºС. Показано, что по мере увеличения концентрации путресцина общее количество лейкоцитов снижается, при этом кадаверин дозозависимо повышал этот показатель. В то же время после воздействия путресцина увеличилась доля больших по размеру лимфоцитов и количество вакуолей в цитоплазме. Кадаверин в концентрации 25 ммоль/л значимо снижал количество клеток с признаками цитолиза. Таким образом, в ходе проведенных исследований установлено, что кадаверин и путресцин, продуцируемые преимущественно прокариотическими клетками, оказывают влияние как на численность, так и на цитологические характеристики иммунокомпетентных клеток.
Полиамины, кадаверин, путресцин, микробная агрессия, лейкоциты
Короткий адрес: https://sciup.org/147241926
IDR: 147241926 | УДК: 571.27 | DOI: 10.17072/1994-9952-2023-3-280-286
Cytological features of mononuclear leukocytes cultured in the presence of polyamines
Quantitative and qualitative changes in the cellular composition after incubation with putrescine and cadaverine are considered. The aim of the study was to study changes in the cytological characteristics of leukocytes after their incubation with cadaverine and putrescine. The number, cell types, sizes, presence of vacuoles and inclusions in peripheral blood leukocytes (n=15) were evaluated after their preincubation with cadaverine (5 and 25 mmol/L) and putrescine (5 and 25 mmol/L) for 72 hours at 37oC. It has been shown that as the concentration of putrescine increases, the total number of leukocytes decreases, while cadaverine dose-dependently increased this indicator. At the same time, after exposure to putrescine, the proportion of large lymphocytes and the number of vacuoles in the cytoplasm increased. Cadaverine at a concentration of 25 mmol/L significantly reduced the number of cells with signs of cytolysis. Thus, in the course of the conducted studies, it was found that cadaverine and putrescine, produced mainly by prokaryotic cells, affect both the number and cytological characteristics of immunocompetent cells.
Текст научной статьи Цитологические особенности мононуклеарных лейкоцитов, культивированных в присутствии полиаминов
Разносторонние биологические эффекты действия полиаминов на функции и жизнедеятельность как прокариотических, так и эукариотических клеток [Годовалов, Карпунина, 2022] обусловливают нарастающий интерес к их изучению. Известно, что полиамины регулируют синтез белка бактериями, их устойчивость к антимикробным препаратам, формирование биопленок, quorum sensing и т.д. [Бухарин, 2018, Нестерова, Негорелова, Ткаченко, 2019]. Показано, что применение путресцина per os приводит к увеличению активности лизоцима и щелочной фосфатазы, уровня иммуноглобулинов М, а также снижает уровень продукции фактора некроза опухоли-α в кишечном содержимом [Liu et al., 2019]. Кроме того, установлено, что полиамины регулируют дифференцировку макрофагов в М2 и оказывают супрессию толерогенных дентритных клеток [Carriche et al., 2021]. В ряде других экспериментов описано ингибирование путресцином и кадаверином продукции IFNγ, а также увеличение продукции IL-4 митоген-активированными клетками [Годовалов и др., 2019]. Продемонстрировано участие полиаминов в регуляции активации и дифференцировки Т- и В-лимфоцитов [Hesterberg, Cleveland, Epling-Burnette, 2018]. Так, дефицит орнитиндекарбоксилазы, фермента, необходимого для синтеза полиаминов, приводит к нарушению дифференцировки CD4+ Т-хелперов из-за снижения концентрации субстрата для дезоксигипузин-синтазы [Puleston et al., 2021]. Ранее было показано, что в очаге воспаления, там, где наблюдается активное размножение условно патогенных микроорганизмов, нарастает уровень полиаминов [Shah et al., 2011; Морозов, Карпунина, Годовалов, 2018; Годовалов, Карпунина, 2022]. Однако изменение активности клеток иммунной системы в таких условиях не изучено. В связи с этим наибольшее внимание исследователей привлекают полиамины, синтезируемые условно патогенными микроорганизмами, как, например, путресцин и кадаверин. Эти полиамины относятся к классу низкомолекулярных алифатических поликатионных аминов, состоящих из насыщенных углеводородных цепей различной длины с аминогруппами [Hesterberg, Cleveland, Epling-Burnette, 2018]. Известно, что полиамины могут активно проникать через цитоплазматическую мембрану клеток при участии специфического переносчика [Mei et al., 2007; Kurihara, 2022] и (или) за счет пассивной диффузии [Силачев, Плотников, Горюнов, 2018]. После проникновения полиаминов в клетку наблюдается процесс их связывания с полианионными соединениями (белки, нуклеиновые кислоты, фосфолипиды, глюкозаминогликаны), а более 90% всех полиаминов находятся в связанном с ДНК и РНК состоянии [Mei et al., 2007; Hesterberg, Cleveland, Epling-Burnette, 2018]. Внутриклеточные функции полиаминов многообразны: стабилизация плазматических мембран, суперсперилизация структуры ДНК, индуцирование транскрипции, связывание с рибосомами, активация ДНК-лигаз, эндонуклеаз, протеинкиназ. Связавшись с нуклеиновыми кислотами, полиамины влияют на структуру хроматина, регулируя тем самым экспрессию генов [Shah, Swiatlo, 2008]. Однако исследований морфологических особенностей эукариотических клеток относительно мало.
Цель исследования – оценка изменений цитологических характеристик лейкоцитов после их инкубации с кадаверином и путресцином.
Материалы и методы
У практически здоровых доноров брали пробы венозной крови с последующим выделением моно-нуклеарной фракции лейкоцитов путем центрифугирования на градиенте плотности фикола-верографина (1.077 г/см3). Рабочее количество лейкоцитов – 200 мононуклеарных клеток на 1 мкл. В стерильные круглодонные планшеты внесли полиамины в концентрациях 5 и 25 ммоль/л. Использовали путресцина гидрохлорид, кадаверина гидрохлорид. В контрольные лунки вносили равный объем среды. Использовали полную культуральную среду, включающую RPMI-1640, 10% эмбриональной телячьей сыворотки, 20 мМ HEPES , 0.05 мМ 2-меркаптоэтанола, 50 мкг/мл гентамицина и 2 мМ L -глютамина. Культивирование осуществляли в следующих условиях: влажная атмосфера с 5%-ным СО 2 при 37ºС в течение 72 ч. После этого готовили микропрепараты из клеточного осадка и фиксировали их по методу Май-Грюнвальда с последующей окраской по методу Романовского – Гимза. В препаратах учитывали типы клеток, определяли их размеры, а также наличие вакуолей и включений.
Статистическую обработку результатов выполняли с помощью парного варианта t -критерия Стьюдента. За пороговый уровень значимости принимали величину p <0.05.
Результаты
Установлено, что по мере увеличения концентрации путресцина в пробах, общее количество моно-нуклеарных лейкоцитов не увеличивается относительно аналогичного показателя в контрольных пробах, что, возможно, обусловлено ингибированием пролиферативной активности этих клеток путресцином. При этом существенных различий между пробами с 5 и 25 ммоль/л не выявлено (рис. 1). В случае культивирования клеток в присутствии кадаверина выявлено увеличение численности моноцитов. При этом в пробах с 5 ммоль/л наблюдается незначительное увеличение их числа, а с увеличением концентрации кадаверина – статистически значимое отличие от контрольных проб (рис. 1). Кроме того, численность клеток в пробах с 5 и 25 ммоль/л кадаверина статистически значимо различалась.
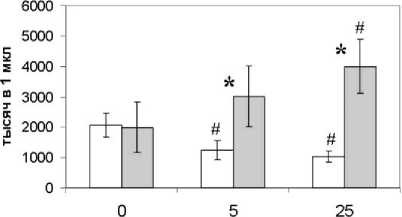
концентрация полиаминов, ммоль/л
□ путресцин □ кадаверин
Рис. 1. Численность мононуклеарных клеток в пробах с полиаминами к 72 ч культивирования.
* – p <0.05 при сравнении кадаверина и путресцина; # – p <0.05 при сравнении с пробами без соответствующего полиамина. Результаты представлены в виде среднего арифметического и его ошибки
[The number of mononuclear cells in samples with polyamines by 72 hours of cultivation.
* – p<0.05 when comparing cadaverine and putrescine; # – p<0.05 when comparing with samples without the corresponding polyamine. The results are presented in the form of an arithmetic mean and its error]
С помощью окрашивания трипановым синим в пробах с разными концентрациями путресцина не выявлено снижения числа жизнеспособных клеток, а также увеличения числа мертвых клеток. При этом в микропрепаратах наблюдается увеличение относительного числа клеток с признаками цитолиза согласно повышению уровня путресцина. При концентрации этого поликатиона 5 ммоль/л – 4.6±0.6%, а при 25 – 5.8±0.6% (в контрольных пробах – 2.6±0.7%, p = 0.051 к пробам с 5 ммоль/л и p = 0.032 к пробам с 25 ммоль/л путресцина).
К 72 ч культивирования клеток с путресцином установлено, что с увеличением концентрации этого поликатиона увеличивается доля моноцитарных клеток, а численность лимфоцитов снижается относительно аналогичных показателей в образцах без полиаминов (рис. 2).
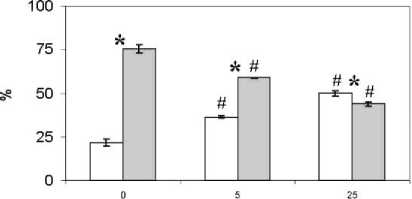
концентрация путресцина, ммоль/л
-
□ моноциты □ лимфоциты
Рис. 2. Лимфоцитарно-моноцитарное соотношение при культивировании клеток в присутствии путресцина.
-
* – p <0.05 при сравнении кадаверина и путресцина; # – p <0.05 при сравнении с пробами без соответствующего полиамина. Результаты представлены в виде среднего арифметического и его ошибки
[Lymphocyte-monocyte ratio in cell culture in the presence of putrescine.
* – p<0.05 when comparing cadaverine and putrescine; # – p<0.05 when comparing with samples without the corresponding polyamine. The results are presented in the form of an arithmetic mean and its error]
При увеличении уровня путресцина в пробах возрастало количество больших по размеру лимфоцитов до 22.1±2.4% при 5 ммоль/л и 34,5±2.9% при 25 ммоль/л (в контроле – 5.3±1.7%, p = 0.001 к пробам с 5 и 25 ммоль/л). Отмечено дозозависимое увеличение клеток, содержащих вакуоли: 20.2±2.3% при 5 ммоль/л; 33.4±1.4% при 25 ммоль/л (в контроле – 10.9±1.5%; p = 0.001 к пробам с 5 и 25 ммоль/л). Однако число клеток, содержащих прочие включения, оставалось наименьшим при 25 ммоль/л (18.9±1.2%) и наибольшим – в контрольных пробах (41.6±7.9%; p = 0,049).
При подсчете клеток с трипановым синим в пробах с разными концентрациями кадаверина установлено увеличение числа жизнеспособных клеток при повышении концентрации этого поликатиона. В микропрепаратах, полученных из культуральной взвеси после 72 ч инкубации в присутствии 25 ммоль/л кадаверина число клеток с признаками цитолиза существенно меньше, чем таковое в контрольных пробах (1.3±0.2 и 3.7±0.6% соответственно; p = 0.02). Изменения лимфоцитарно-моноцитарного соотношения в пробах мононуклеарных лейкоцитов, культивированных в присутствии разных концентраций кадаверина, представлены на рис. 3.
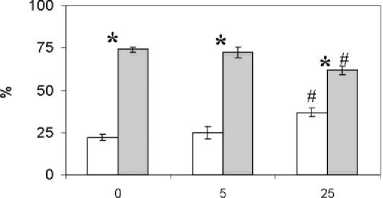
концентрация кадаверина, ммоль/л
□ МО ноциты □ лимфоциты
Рис. 3. Лимфоцитарно-моноцитарное соотношение при культивировании клеток в присутствии кадаверина.
* – p <0.05 при сравнении между кадаверином и путресцином; # – p <0.05 при сравнении с пробами без соответствующего полиамина. Результаты представлены в виде среднего арифметического и его ошибки
[Lymphocyte-monocyte ratio in cell culture in the presence of cadaverine.
* – p<0.05 when compared between cadaverine and putrescine; # – p<0.05 when compared with samples without the corresponding polyamine. The results are presented in the form of an arithmetic mean and its error]
Отмечено значительное уменьшение количества малых лимфоцитов (до 9.0±0.8%) и повышение лимфоцитов большого размера (до 20.9±1.0%) при уровне кадаверина 25 ммоль/л (в контрольных пробах – 22.3±1.0%, p = 0.001 и 9.6±0.9%, p = 0.001 соответственно). Кроме того, в пробах с 25 ммоль/л кадаверина выявлено значимое увеличение доли клеток, содержащих вакуоли и включения – 26.9±3.0% (в контроле – 9.5±1.6%, p = 0.001).
Обсуждение
Известно, что в метаболизме моноцитарно-макрофагальных клеток существенное значение играют полиамины, однако большинство исследователей сконцентрированы на изучении влияния спермина и спермидина, являющихся продуктами эукариотических клеток. Кроме этого, есть сведения, что концентрация полиаминов увеличивается при воспалении, особенно в случае наличия полиамин-синтезирующих видов бактерий, среди которых следует выделить представителей семейства Enterobacte-riaceae , рода Neisseria и других [Tabor, Tabor, 1985; Lavizzari et al., 2010]. Однако эти виды микроорганизмов синтезируют другие поликатионы – кадаверин и путресцин, о влиянии которых на иммунокомпетентные клетки относительно мало сведений.
Путресцин оказывает большее влияние на моноциты, чем кадаверин, поскольку может включаться в метаболизм этих клеток с последующим образованием спермина и спермидина [Kurihara, 2022]. Известно, что рост числа клеток с вакуолями среди моноцитов может указывать на их активацию [Сидельников, Начева, 2016; Бархина, Гущин, Алексеев, 2018; Kurihara, 2022], которая, как показано в настоящем исследовании длится дольше, поскольку число клеток с включениями существенно не изменяется. Кроме того, в проведенных ранее исследованиях установлено, что путресцин оказывает преимущественное влияние на фагоцитарную активность нейтрофилов, но не моноцитов [Годовалов и др., 2019].
Зарегистрированное повышение числа жизнеспособных клеток при их инкубации с кадаверином обусловлено протективным действием этого полиамина при наличии факторов, инициирующих цитолиз клеток, что можно наблюдать в искусственных условиях. Увеличение числа клеток с включениями можно рассматривать как отражение увеличения поглотительной активности моноцитов, что подтверждается проведенными нами ранее исследованиями [Морозов, Карпунина, Годовалов, 2018; Годовалов и др., 2019].
Наблюдаемая в настоящем исследовании вакуолизация цитоплазмы, вероятно, не является признаком развивающейся клеточной смерти, а относится к побочным эффектам действия полиаминов [Shubin, Demidyuk, Komissarov, 2016]. От эндоплазматического ретикулюма отщепляются вакуоли, что повышает емкость этого органоида, в том числе для белка. Более того вакуоли аккумулируя бактериальные токсины снижают их патогенный эффект (например, токсины E. coli Stx2 и SubAB, токсин M. pneumoniae CARDS и некоторые другие [Shubin, Demidyuk, Komissarov, 2016]). Можно предположить, что такая ситуация будет способствовать внутриклеточной персистенции бактерий-продуцентов полиаминов.
Заключение
Таким образом, в ходе проведенных исследований установлено, что кадаверин и путресцин, продуцируемые преимущественно прокариотическими клетками, оказывают влияние как на численность, так и на цитологические характеристики иммунокомпетентных клеток.
Список литературы Цитологические особенности мононуклеарных лейкоцитов, культивированных в присутствии полиаминов
- Бархина Т.Г., Гущин М.Ю., Алексеев Ю.В. Ультраструктурный анализ различных популяций клеток крови при аллергопатологии и воздействии светокислородного и фотодинамического эффектов // Вестник новых медицинских технологий. 2018. Т. 12, № 4. С. 218–224.
- Бухарин О.В. Адаптивные стратегии взаимодействия возбудителя и хозяина при инфекции // Вестник Российской академии наук. 2018. Т. 88, № 7. С. 637–643.
- Годовалов А.П., Карпунина Т.И. Влияние полиаминов бактериального происхождения на продукцию ключевых цитокинов в культуре мононуклеарных лейкоцитов человека // Медицинская иммунология. 2022. Т. 24, № 2. С. 257–262.
- Годовалов А.П. и др. Полиамины как рецептор-независимые факторы агрессии условно-патогенных микроорганизмов // Иммунопатология, аллергология, инфектология. 2019. № 3. С. 91–94.
- Годовалов А.П. и др. Люминолзависимая хемилюминесценция как средство выявления маркеров окислительного стресса // Высокие технологии, определяющие качество жизни: материалы II Междунар. науч. конф. Пермь, 2018. С. 201–203.
- Морозов И.А., Карпунина Т.И., Годовалов А.П. Кадаверин как регулятор активности про- и эукариотических клеток // Аллергология и иммунология. 2018. Т. 19, № 3. С. 149–150.
- Нестерова Л.Ю., Негорелова Е.В., Ткаченко А.Г. Биогенные полиамины как модуляторы активности Quorum sensing системы и биопленкообразования Vibrio harveyi // Вестник Пермского университета. Сер. Биология. 2019. Вып. 3. C. 300–308.
- Сидельникова А.А., Начева Л.В. Морфологические изменения периферической крови в динамическом наблюдении при остром описторхозе у экспериментальных животных // Современные проблемы науки и образования. 2016. № 6. С. 504–512.
- Силачев Д.Н., Плотников Е.Ю., Горюнов К.В. Роль полиаминов в жизнедеятельности клеток репродуктивной системы // Цитология. 2018. Т. 60, № 3. С. 164–172.
- Carriche G.M. et al. Regulating T-cell differentiation through the polyamine spermidine // The journal of allergy and clinical Immunology. 2021. Vol. 147, № 1. Р. 335–348.
- Equi A.M. et al. Oxidation of putrescine and cadaverine derivatives by diamine oxidases // Tetrahedron. 1991. Vol. 47, № 3. Р. 507–518.
- Fujiswa S., Kadoma Y. Kinetic evaluation of polyamines as radical scavengers // Anticancer Research. 2005. Vol. 25, № 2A. Р. 965–970.
- Hesterberg R.S., Cleveland J.L., Epling-Burnette P.K. Role of polyamines in immune cell functions // Medical sciences (Basel, Switzerland). 2018. Vol. 6, № 1. 22. doi: 10.3390/medsci6010022.
- Kurihara S. Polyamine metabolism and transport in gut microbes // Bioscience, biotechnology, and bio-chemistry. 2022. Vol. 86, № 8. P. 957–966.
- Lavizzari T. et al. Histamine, cadaverine, and putrescine produced in vitro by enterobacteriaceae and pseudomonadaceae isolated from spinach // Journal of food protection. 2010. Vol. 73, № 2. Р. 385–389.
- Liu G. et al. Putrescine enhances intestinal immune function and regulates intestinal bacteria in weaning piglets // Food Function. 2019. Vol. 10. Р. 4134–4142.
- Mei Y. et al. A sequential injection analysis chemiluminescent plant tissue-based biosensor system for the determination of diamine // Biosens. Bioelectron. 2007. Vol. 22, № 6. Р. 871–876.
- Puleston D.J. et al. Polyamine metabolism is a central determinant of helper T cell lineage fidelity // Cell Vol. 2021. Vol. 184. Р. 4186–4202.
- Shah P. et al. Polyamine biosynthesis and transport mechanisms are crucial for fitness and pathogenesis of Streptococcus pneumoniae // Microbiology. 2011. Vol. 57, № 2. Р. 504–515.
- Shah P., Swiatlo E. A multifaceted role for polyamines in bacterial pathogens // Molecular microbiology. 2008. Vol. 68, № 1. р. 4–16.
- Shubin A.V., Demidyuk I.V., Komissarov A.A. Cytoplasmic vacuolization in cell death and survival // Oncotarget. 2016. Vol. 7, № 34. р. 55863–55889.
- Tabor C.W., Tabor H. Polyamines in microorganisms // Microbiological Reviews. 1985. Vol. 49, № 1. Р. 81–99.
