Comparative Analysis of Seed Germination Under Salinity Stress Induced by Neutral and Alkaline Salts

Author: Sreelakshmi Rajesh, Bahira R.B., Renu Rajan
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 4 т.21, 2025.
Free access
Salinity stress is a critical abiotic factor limiting seed germination and early seedling development, particularly in glycophytic species. This study assessed the effects of neutral (NaCl) and alkaline (NaHCO₃) salt stress on seed germination parameters in two crop species, Vigna unguiculata (cowpea) and Setaria italica (foxtail millet). Seeds were exposed to varying concentrations of NaCl and NaHCO₃ to evaluate germination percentage, radicle and plumule emergence, and overall seedling vigor. Both species exhibited significant reductions in germination and seedling growth under increasing salt concentrations, with NaHCO₃ exerting a more severe inhibitory effect than NaCl. This heightened inhibition is attributed to the elevated pH caused by alkaline salts, which further disrupts physiological processes critical for germination. While both species are inherently sensitive to salinity, Vigna unguiculata demonstrated comparatively higher tolerance, maintaining better germination rates and seedling growth under stress, indicating the presence of adaptive mechanisms such as osmotic adjustment or ion compartmentalization. These findings highlight the need for selecting salt-tolerant species and implementing effective salinity management practices to improve crop establishment and productivity in salt-affected regions.
Abiotic stress, Seed germination, Salt tolerance
Short address: https://sciup.org/143185119
IDR: 143185119
Text of the scientific article Comparative Analysis of Seed Germination Under Salinity Stress Induced by Neutral and Alkaline Salts
Stress on plant germination refers to any environmental or biological factor that negatively affects the ability of seeds to sprout and develop into healthy seedlings. These stresses can delay germination, reduce germination rates, or even prevent seeds from germinating altogether ( i et al. , 2013).
There are several types of stress that can impact plant germination. Abiotic stress includes non-living factors such as drought, extreme temperatures (either too hot or too cold), salinity, and poor soil conditions, all of which can limit water uptake or damage seed structures (Yadav et al. , 2020). Biotic stress, on the other hand, involves living organisms like bacteria, fungi, insects, or other pests that can attack seeds or emerging seedlings. Both types of stress can significantly hinder plant growth and crop yield. Understanding and managing these stresses is essential for improving agricultural productivity and ensuring successful seed germination(Biswas & Das, 2024).
Salt stress is a problem that happens when there is too much salt in the soil. It can make it hard for plants to grow. When plants take in salty water, it can hurt their roots and stop them from getting the water and nutrients they need. This can cause plants to grow slowly, have yellow leaves, or even die(Atta et al. , 2023a). Salt stress can also affect the way plants make food through photosynthesis. In strong salt conditions, plants may produce fewer flowers and fruits. Overall, salt stress can greatly reduce the health and growth of plants(Perras et al. , 2023).
Soil salinization is an essential environmental stressor, threatening agricultural yield and ecological security world wide. Saline soil accumulate excessive soluble salts which are dertimental to most plants but limiting plant growth and productivity (Tarolli et al. , 2024).
Different types of salt in the soil can affect how well seeds grow into plants. Acidic salts, like aluminum or iron salts, can hurt the seed and make it hard for it to take in water, which can slow or stop germination (Hadjadj et al., 2022). Alkaline salts, such as sodium carbonate, can raise the pH of the soil and damage the seed or root tips, making it harder for seeds to grow properly. Neutral salts, like common table salt (sodium chloride), can also stop seeds from taking in water and cause stress (Wang et al., 2022a).
High salt concentrations in soil can hinder seed germination and impair the growth of healthy plants, with increasing salinity levels intensifying these negative effects. While most plants are sensitive to salinity, a few halophytes possess the ability to thrive under such stress (Nikolić et al. , 2023a). Excess sodium ions (Na ⁺ ) disrupt ionic balance by reducing potassium (K ⁺ ) levels, leading to altered cellular metabolism and stunted plant development. Moreover, salt-induced osmotic stress limits water uptake and triggers the overproduction of reactive oxygen species (ROS), which damage cellular membranes and proteins, disrupt electrochemical gradients, and interfere with nutrient absorption by membrane transporters (Waheed et al. , 2024).
Salt can be neutral, alkaline or acidic based on its composition. Neutral (NaCl) and alkaline (NaHCO₃) salt stresses significantly affect plant germination and early growth, albeit through different mechanisms. Neutral salts primarily induces osmotic stress, reducing water uptake and delaying seed metabolic activation, which hampers germination and seedling development. At higher concentrations, NaCl can also cause ion toxicity, leading to membrane damage and oxidative stress. However, some plants exhibit partial tolerance to NaCl, especially at moderate concentrations. Alkaline salts combines osmotic stress with high pH-induced alkalinity, resulting in more severe effects than neutral salts. Alkaline conditions disrupt nutrient uptake, particularly potassium (K ⁺ ), and increase the production of reactive oxygen species (ROS), leading to oxidative damage. Studies have shown that NaHCO₃ stress more significantly suppresses seed germination and seedling growth compared to NaCl (Wang et al. , 2022; Guo et al. , 2010)
The present study was undertaken with the objective of assessing the effects of salt stress on seed germination in two plant species, Vigna unguiculata and Setaria italica. Specifically, the study aimed to evaluate the impact of both alkaline (NaHCO₃) and neutral (NaCl) salt stress at different concentrations on seed germination. It also sought to compare the rate of plumule and radicle emergence in these two species under the influence of varying salt levels. Furthermore, a comparative analysis of germination responses and salt tolerance between Vigna unguiculata and Setaria italica under NaHCO₃ and NaCl treatments was conducted to better understand species-specific responses to salinity and alkalinity stress during the early stages of plant development.
MATERIALS AND METHODS
Collection of seeds – Vigna ungiculata and Sataria italic a
Vigna ungiculata and Sataria italica seeds were bought from market. Fresh and healthy seeds were handpicked and used for further experimentation
Alkaline and neutral salts – NaCl and NaHCO3Preparation of salts of different concentrations
Two types of salts—sodium chloride (NaCl) and sodium bicarbonate (NaHCO 3 ) were prepared in different concentrations [millimolar (mM)] namely; 50 mM, 100 mM, 150 mM, and 200 mM. To calculate the amount of salt required for a specific molarity and volume, the equation used:
Mass (g) = Molarity (mol/ ) × Volume ( ) × Molar Mass (g/mol)
Molarity (mol/ )- Molarity, denoted as M, represents the concentration of a solution.
Volume ( )- This is the total volume of the solution you intend to prepare, expressed in litres ( ).
Molar Mass (g/mol)- Molar mass is the mass of one mole of a substance, expressed in grams per mole (g/mol).
A calibrated analytical balance for precise measurement of the salt was used. The salt was dissolved completely before adjusting to the final volume to ensure uniform concentration. volumetric flasks were used for accurate volume measurements. All solutions were labelled with concentration, date of preparation, and any other relevant information.
Germination test
Experiment was set up by labelling Petri dishes for each salt concentration. Place a piece of filter paper in each dish and moisten it with 10 m of the corresponding salt solution. Place 10 sterilized seeds evenly spaced on the moistened filter paper in each dish. Cover the dishes with lids or wrap them with transparent plastic to prevent evaporation. The dishes were incubated in a growth chamber set at optimal conditions in appropriate darkness and specific light conditions.
Monitor the seeds daily for 48 hours. A seed is considered germinated when the radicle (embryonic root) emerges at least 2 mm. Record the number of germinated seeds and note any abnormalities in seedling development. On reaching 48 hour, record the final germination count.
Germination Percentage = (Number of Germinated Seeds / Total Seeds) × 100
Salt Tolerance Index (STI) calculation
The Salt Tolerance Index (STI) were calculated to assess the impact of salt stress:
STI = (Germination Percentage under Salt Stress / Germination Percentage under Control) × 100
Observation of length of radicle and plumule
When ready to measure, the seedlings were carefully picked up using forceps to avoid damage. Place each seedling on a flat surface with a contrasting background to enhance visibility. Using a ruler or digital caliper with at least 0.1 mm precision, measure the length of the radicle from its point of emergence to its tip. Similarly, measure the plumule from its base to the tip of the shoot. For curved structures, a flexible measuring tool like a marked string can be used to follow the contour accurately. Record all measurements systematically. After collecting data from all samples, average lengths were calculated for each treatment group. These measurements can be used to assess seedling vigor and the effects of various treatments on early plant development.
RESULTS
The germination percentage of Vigna unguiculata was assessed under varying concentrations of NaCl and NaHCO₃. For NaCl treatment, the seeds exhibited 100% germination at both 50 mM and 100 mM concentrations, which then decreased to 90% at 150 mM and 80% at
200 mM. Similarly, under NaHCO₃ treatment, 100% germination was observed at 50 mM, which decreased to 90% at 100 mM, 80% at 150 mM, and 70% at 200 mM. These results indicate a gradual reduction in germination percentage with increasing salt concentration for Vigna unguiculata (Table 1).
For Setaria italica , the germination percentage under NaCl treatment followed a similar trend, with 100% germination at 50 mM, decreasing to 90% at 100 mM, 80% at 150 mM, and 70% at 200 mM. In the case of NaHCO₃ treatment, the seeds showed 100% germination at 50 mM, which decreased to 80% at 100 mM, 70% at 150 mM, and 70% at 200 mM. ike Vigna unguiculata , the germination of Setaria italica was negatively impacted by increasing concentrations of both salts, with NaCl and NaHCO₃ showing similar effects [Table 2].
Both Vigna unguiculata and Setaria italica demonstrated a decrease in germination percentage as the salt concentrations of NaCl and NaHCO₃ increased. However, Vigna unguiculata exhibited a slightly more consistent germination at higher concentrations of both salts, with a 10% higher germination rate at 200 mM NaCl and NaHCO₃ compared to Setaria italica , which had a greater reduction in germination at 200 mM. Both species show a trend of salt sensitivity, with NaCl and NaHCO₃ inhibiting germination more at higher concentrations.
The salt tolerance index of Vigna unguiculata was measured under varying concentrations of NaCl and NaHCO₃. For NaCl treatment, the salt tolerance index remained at 100% at 50 mM, decreased to 90% at 100 mM, stayed at 90% at 150 mM, and decreased further to 80% at 200 mM. Similarly, under NaHCO₃ treatment, the salt tolerance index was 100% at 50 mM, decreased to 90% at 100 mM, further declined to 80% at 150 mM, and dropped to 70% at 200 mM. These results indicate that the salt tolerance of Vigna unguiculata decreased progressively as salt concentrations increased [Table:3].
For Setaria italica , the salt tolerance index under NaCl treatment followed a similar pattern, with 100% at 50 mM, 90% at 100 mM, 80% at 150 mM, and 70% at 200 mM. Under NaHCO₃ treatment, the salt tolerance index remained at 100% at 50 mM, decreased to 90% at
100 mM, dropped to 80% at 150 mM, and remained at 70% at 200 mM. The trend for both NaCl and NaHCO₃ treatments showed a gradual decline in the salt tolerance index with increasing salt concentration [Table:4].
Both Vigna unguiculata and Setaria italica exhibited a similar trend in their salt tolerance indices, with both species maintaining a high tolerance at lower salt concentrations (50 mM and 100 mM) and showing a decrease in tolerance as the salt concentration increased. However, Setaria italica had a more consistent decline in its salt tolerance index, with a steady drop from 100% to 70% at 200 mM for both NaCl and NaHCO₃. On the other hand, Vigna unguiculata maintained a higher salt tolerance index at 200 mM compared to Setaria italica, especially under NaCl treatment (80% vs. 70%). Both species are sensitive to higher salt concentrations, but Vigna unguiculata demonstrates relatively better salt tolerance.
The length of the plumule was measured after 48 hours of NaCl salt stress induction for both Vigna unguiculata and Setaria italica . For Vigna unguiculata , the control group showed a plumule length of 2.72 ± 0.75 mm. At increasing NaCl concentrations, the plumule length decreased progressively: 1.253 ± 0.435 mm at 50 mM, 2.72 ± 0.75 mm at 100 mM (similar to the control), 1.74 ± 0.21 mm at 150 mM, and 1.09 ± 0.39 mm at 200 mM. In contrast, for Setaria italica , the control group had a plumule length of 1.17 ± 0.29 mm. The plumule length decreased significantly at increasing concentrations of NaCl: 0.75 ± 0.22 mm at 50 mM, 0.39 ± 0.09 mm at 100 mM, and 0.06 ± 0.05 mm at 200 mM [Table:5].
For the length of the radicle, Vigna unguiculata had a control length of 2.88 ± 0.43 mm. At 50 mM NaCl, the radicle length decreased slightly to 2.83 ± 0.26 mm, and further reduced to 2.72 ± 0.75 mm at 100 mM. A sharp decline was observed at 150 mM (1.09 ± 0.39 mm), and a significant reduction occurred at 200 mM (0.16 ± 0.05 mm). On the other hand, Setaria italica exhibited a radicle length of 1.66 ± 0.37 mm in the control group. As NaCl concentration increased, the radicle length decreased progressively: 1.41 ± 0.39 mm at 50 mM, 1.01 ± 0.45 mm at 100 mM, and 0.59 ± 0.14 mm at 200 mM [Table:6].
Both species exhibited a reduction in the length of the plumule and radicle with increasing NaCl salt concentrations. Vigna unguiculata maintained a relatively higher growth in both plumule and radicle compared to Setaria italica , particularly at higher NaCl concentrations. Setaria italica showed more pronounced inhibition, especially in the plumule growth, where its length almost diminished entirely at 200 mM NaCl. Vigna unguiculata exhibited a less severe reduction in both plumule and radicle growth, demonstrating relatively better tolerance to salt stress.
The length of the plumule was measured after 48 hours of NaHCO₃ salt stress induction for both Vigna unguiculata and Setaria italica . For Vigna unguiculata , the control group showed a plumule length of 2.72 ± 0.75 mm. At increasing concentrations of NaHCO₃, the plumule length decreased: 2.19 ± 0.10 mm at 50 mM, 1.79 ± 0.04 mm at 100 mM, 1.253 ± 0.435 mm at 150 mM, and 0.93 ± 0.04 mm at 200 mM. In contrast, for Setaria italica, the control group had a plumule length of 2.06 ± 0.05 mm. The plumule length decreased progressively at increasing concentrations of NaHCO₃: 1.51 ± 0.05 mm at 50 mM, 1.00 ± 0.50 mm at 100 mM, and 0.81 ± 0.07 mm at 150 mM, with no measurement at 200 mM [Table:7].
For the radicle length, Vigna unguiculata had a control length of 2.88 ± 0.43 mm. At 50 mM NaHCO₃, the radicle length decreased to 1.77 ± 0.07 mm, further reduced to 1.253 ± 0.435 mm at 100 mM, and 1.04 ± 0.06 mm at 150 mM. A significant reduction was observed at 200 mM, with a radicle length of 0.56 ± 0.07 mm. On the other hand, Setaria italica exhibited a control radicle length of 1.63 ± 0.10 mm. As NaHCO₃ concentration increased, the radicle length decreased: 1.12 ± 0.06 mm at 50 mM, 0.92 ± 0.49 mm at 100 mM, and 0.44 ± 0.12 mm at 150 mM, with no measurement at 200 mM [Table:8].
Both species exhibited a reduction in the length of the plumule and radicle as NaHCO₃ salt stress increased. Vigna unguiculata showed a gradual decrease in both plumule and radicle length, but the species still maintained higher growth compared to Setaria italica , particularly at higher NaHCO₃ concentrations. For instance, at 200 mM NaHCO₃, Vigna unguiculata had a measurable radicle length (0.56 ± 0.07 mm), while Setaria italica showed no measurement. Overall, Setaria italica demonstrated greater sensitivity to NaHCO₃ stress, with both plumule and radicle growth severely inhibited at higher concentrations, whereas Vigna unguiculata maintained relatively better growth.
Table 1: Germination percentage of Vigna unguiculata
|
S No |
Salt |
Number of seeds germinated |
||||
|
control |
50 |
100 |
150 |
200 |
||
|
1. |
NaCl |
100 |
100 |
90 |
90 |
80 |
|
2. |
NaHCO 3 |
100 |
100 |
90 |
80 |
70 |
Table 2: Germination percentage of Setaria italica
|
S No |
Salt |
Number of seeds germinated |
||||
|
control |
50 |
100 |
150 |
200 |
||
|
1. |
NaCl |
100 |
100 |
90 |
80 |
70 |
|
2. |
NaHCO 3 |
100 |
100 |
80 |
70 |
70 |
Table 3: Salt Tolerance index of Vigna unguiculata
|
S No |
Salt |
Salt Tolerance Index (in%) |
|||
|
50 |
100 |
150 |
200 |
||
|
1. |
NaCl |
100 |
90 |
90 |
80 |
|
2. |
NaHCO 3 |
100 |
90 |
80 |
70 |
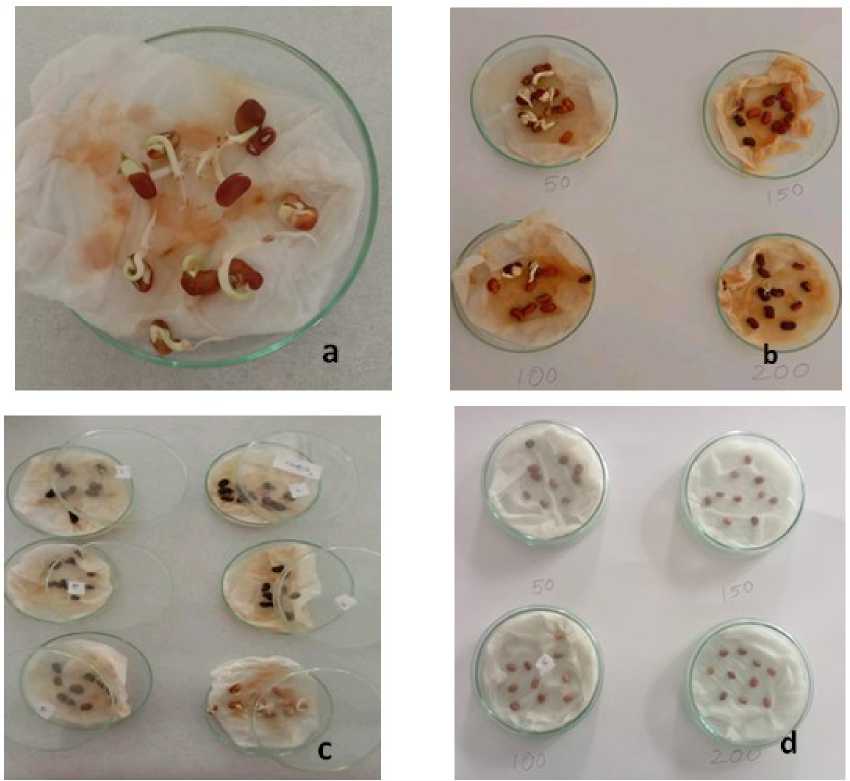
Figure 1: Fig. (a): control plate with full germinated seeds; Fig (b) Vigna unguiculata treated with 50 mmol, 100 mmol, 150 mmol and 200 mmol NaCl; Fig. (c): Vigna unguiculata seeds treated with 50 mmol, 100 mmol, 150 mmol and 200 mmol concentrations of NaHCO 3 ; Fig (d): Germinated Vigna unguiculata seeds on NaHCO 3 after 48 hrs in concentrations of 50 mmol, 100 mmol, 150 mmol and 200 mmol.
Table 4: Salt Tolerance index of Setaria italica
|
S No |
Salt |
Salt Tolerance Index (in%) |
||||
|
control |
50 |
100 |
150 |
200 |
||
|
1. |
NaCl |
100 |
100 |
90 |
80 |
70 |
|
2. |
NaHCO 3 |
100 |
100 |
80 |
70 |
70 |
Table 5: engthof plumule observed (in mm) after 48 hours of NaCl salt stress induction
|
S.No |
Seed |
control |
50 |
100 |
150 |
200 |
|
1 . |
Vigna unguiculata |
2.72 ± 0.75 |
1.253 ± 0.435 |
2.72 ± 0.75 |
1.74 ± 0.21 |
1.09 ± 0.39 |
|
2. |
Setaria italica |
1.17 ± 0.29 |
0.75 ± 0.22 |
0.39 ± 0.09 |
0.06 ± 0.05 |
Table 6: engthof radicle observed (in mm) after 48 hours of NaCl salt stress induction
|
S. No |
control |
50 |
100 |
150 |
200 |
|
|
1. |
Vigna unguiculata |
2.88 ± 0.43 |
2.83 ± 0.26 |
2.72 ± 0.75 |
1.09 ± 0.39 |
0.16 ± 0.05 |
|
2. |
Setaria italica |
1.66 ± 0.37 |
1.41 ± 0.39 |
1.01 ± 0.45 |
0.59 ± 0.14 |

Figure 2: Fig (a): Sateria italica seeds treated with NaHCO 3 concentrations of 50 mmol, 100 mmol, 150 mmol and 200 mmol along with control plate; Fig (b): Sateria italica seeds on NaCl after 48 hrs in concentrations of 50 mmol, 100 mmol, 150 mmol and 200 mmol along with control plate; Fig (c): Vigna unguiculata seeds; Fig (d) Sateria italica seeds.
Table 7: engthof plumule observed (in mm) after 48 hours of NaHCO 3 salt stress induction
|
.No |
seed |
control |
50 |
100 |
150 |
200 |
|
1 . |
Vigna unguiculata |
2.72 ± 0.75 |
2.19±0.10 |
1.79±0.04 |
1.253 ± 0.435 |
0.93±0.04 |
|
2. |
Setaria italica |
2.06±0.05 |
1.51±0.05 |
1.00 ± 0.50 |
0.81±0.07 |
Table 8: engthof radicle observed (in mm) after 48 hours of NaHCO 3 salt stress induction
|
S.No |
seed |
control |
50 |
100 |
150 |
200 |
|
1 . |
Vigna unguiculata |
2.88 ± 0.43 |
1.77±0.07 |
1.253 ± 0.435 |
1.04±0.06 |
0.56±0.07 |
|
2. |
Setaria italica |
1.63±0.10 |
1.12±0.06 |
0.92 ± 0.49 |
0.44±0.12 |
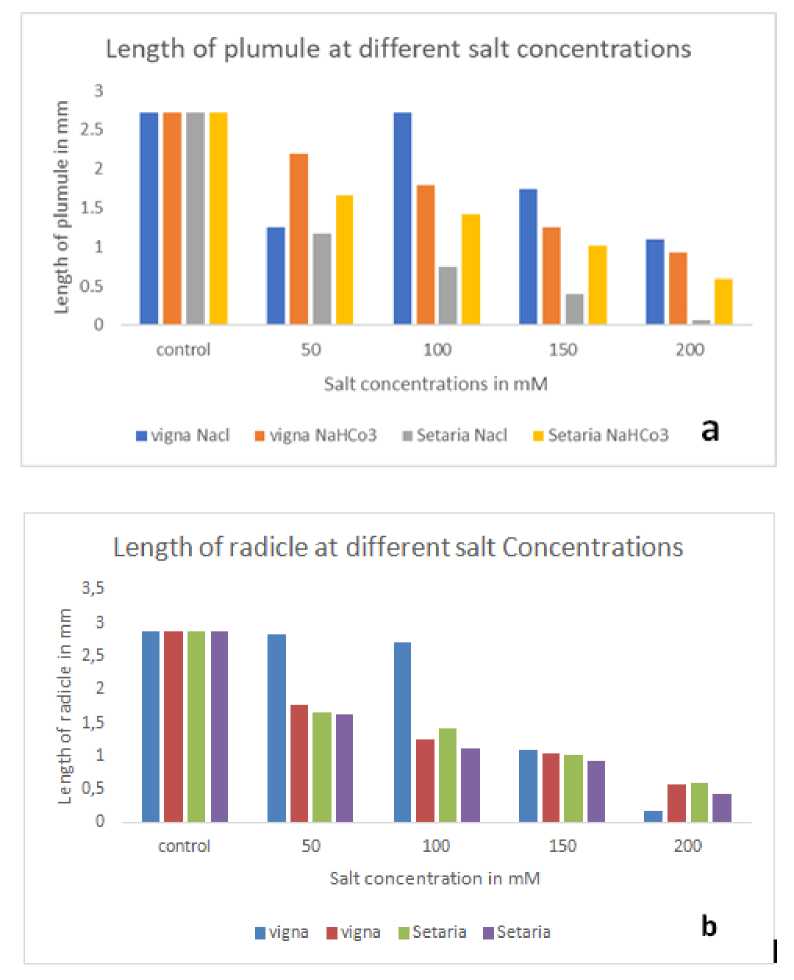
Figure 3: (a) showing length of plumule in mm at different salt concentrations; (b): showing length of radicle in mm at different salt concentrations.
DISCUSSION
Vigna unguiculata maintained a relatively higher salt tolerance index at 200 mM NaCl, which suggests that the species may have better mechanisms for ion regulation and osmotic balance. The higher tolerance observed in Vigna unguiculata , particularly at 200 mM NaCl, could be attributed to its ability to compartmentalize sodium ions in vacuoles or its ability to exclude sodium from sensitive tissues, a characteristic feature of salt-tolerant plants (Munns & Tester, 2008).
On the other hand, Setaria italica showed a more consistent decline in salt tolerance as salinity increased, which is typical of salt-sensitive species. Plants like Setaria italica , which have limited mechanisms to exclude or compartmentalize sodium, experience more severe disruptions in cellular function under saline conditions. This corresponds with the sharp drop in the salt tolerance index of Setaria italica observed in my study, particularly at the higher concentrations of NaCl and NaHCO₃.
The observed reduction in plumule and radicle lengths in both Vigna unguiculata and Setaria italica under increasing NaCl and NaHCO₃ concentrations aligns with established patterns of salt stress effects on early plant development. Notably, Vigna unguiculata maintained relatively higher growth metrics compared to Setaria italica , indicating a greater tolerance to salinity stress.
Previous studies have demonstrated that Vigna unguiculata exhibits moderate salt tolerance, maintaining seedling growth under saline conditions. For instance, research has shown that cowpea seedlings can sustain growth parameters such as shoot and root lengths under NaCl stress, although reductions are observed at higher concentrations. This resilience is attributed to mechanisms like osmotic adjustment and ion compartmentalization, which help mitigate the detrimental effects of salt stress (Rathinapriya et al. , 2020).
In contrast, Setaria italica has been reported to be more sensitive to salinity. Studies indicate that foxtail millet experiences significant reductions in germination and seedling growth under NaCl stress, with higher concentrations leading to pronounced inhibition of root and shoot development . This sensitivity is likely due to less efficient salt tolerance mechanisms, making Setaria italica more susceptible to the adverse effects of salinity(Terfa et al. , 2025).
The differential responses observed between the two species in the current study are consistent with these findings. Vigna unguiculata 's ability to maintain measurable plumule and radicle lengths at higher salt concentrations suggests a more robust tolerance mechanism, while the severe inhibition observed in Setaria italica underscores its sensitivity to saline conditions. These insights are crucial for agricultural practices, especially in regions prone to soil salinization.
Selecting and cultivating salt-tolerant species like Vigna unguiculata can enhance crop productivity and sustainability in such challenging environments.
The experimental findings indicate that both Vigna unguiculata and Setaria italica are glycophytic species, inherently sensitive to salinity stress. Salinity adversely affects seed germination primarily through osmotic stress, which impedes water uptake, and ion toxicity, resulting from the accumulation of harmful ions like Na ⁺ and Cl ⁻ . Additionally, alkaline salts such as NaHCO₃ exacerbate stress by increasing the pH of the germination medium, further inhibiting seed germination and early seedling growth. The data demonstrate that elevated concentrations of both NaCl and NaHCO₃ significantly reduce germination rates and impede seedling development in both species. Notably, NaHCO₃ exhibited a more pronounced inhibitory effect compared to NaCl, likely due to its alkalinity, which can disrupt cellular processes and enzyme activities essential for germination.
While both species are affected by salinity, Vigna unguiculata displayed relatively better tolerance under certain conditions, maintaining higher germination percentages and longer plumule and radicle lengths at elevated salt concentrations. This suggests that Vigna unguiculata may possess more effective mechanisms to cope with salt-induced stress, such as efficient ion compartmentalization or osmotic adjustment..
CONCLUSIONS
These findings underscore the importance of selecting salt-tolerant crops for cultivation in saline-prone areas. Implementing strategies such as soil salinity management, use of salt-tolerant cultivars, and agronomic practices that mitigate salt stress can enhance germination success and ensure sustainable agricultural productivity in affected regions.
Future research on the impact of salinity stress on seed germination in Vigna unguiculata and Setaria italica can be directed toward several key areas to enhance understanding and improve crop resilience. Molecular and genetic studies, such as transcriptomic analyses, could uncover specific genes and regulatory pathways involved in salinity tolerance during seed germination. Investigating the role of microRNAs (miRNAs) in regulating stress-responsive genes may also offer novel insights into plant adaptation mechanisms under salt stress. Additionally, the application of beneficial elements like selenium (Se) has shown potential in mitigating salt-induced damage, as observed in related studies on Setaria italica. Breeding programs aimed at identifying and developing salt-tolerant genotypes, especially in cowpea, could play a significant role in sustainable agriculture. astly, extending the scope of research beyond germination to include seedling, vegetative, and reproductive stages will provide a comprehensive understanding of salinity effects across the entire plant life cycle. These future directions are vital for addressing the challenges posed by soil salinization and for supporting crop productivity in salt-affected regions.
ACKNOWLEGEMENT
The authors gratefully acknowledge, Union Christian college for providing necessary facilities for the project.
CONFLICTS OF INTEREST
The author declare that he has no potential conflicts of interest.
