Дегидрины в почках Betula pendula Roth: особенности сезонной динамики

Автор: Татаринова Татьяна Дмитриевна, Перк Александр Александрович, Бубякина Виктория Витальевна, Пономарев Анатолий Гаврильевич, Ветчинникова Лидия Васильевна, Васильева Ирина Вениаминовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 3-2 т.15, 2013 года.
Бесплатный доступ
В условиях Центральной Якутии и Карелии дегидрины в почках березы повислой ( Betula pendula Roth) представлены низко- и среднемолекулярными формами. У всех растений круглогодично выявлялись дегидрины с мол. м. 66 и 69 кДа со слабо выраженной сезонной динамикой. Напротив, низкомолекулярные дегидрины, преимущественно с мол. м. 17 кДа, имели более отчетливую сезонную динамику. У «карельских» берез они обнаруживались в сентябре, у «якутских» – в августе, сохранялись в течение всей зимы и исчезали только в начале (Карелия) или конце мая (Центральная Якутия). Предполагается непосредственное участие этих белков в формировании морозоустойчивости растений в разных климатических условиях.
Центральная якутия, карелия, суммарные белки, дегидрины, сезонная динамика
Короткий адрес: https://sciup.org/148201869
IDR: 148201869 | УДК: 574.24
Dehydrins of Betula pendula Roth bads: characteristics of seasonal dynamics
Dehydrins of buds of the Asian white birch ( Betula pendula Roth) are presented by low and middle molecular weight forms. Dehydrins with molecular wts of 66 and 69 kDa were detected the whole year round. They showed weak seasonal changes. Conversely, low molecular weight dehydrins, predominantly with molecular wt 17 kDa, were characterized by the largest seasonal changes. In Karelia they were detected in September, in Yakutia – in August, and were present in large amounts in winter and absent early (Karelia) or late in May (Central Yakutia). The results suggest that these proteins participate in the development of frost hardiness of plants in different climatic conditions.
Текст научной статьи Дегидрины в почках Betula pendula Roth: особенности сезонной динамики
Значительную часть территории Российской Федерации, находящейся под лиственными древесными породами, занимает береза повислая ( Betula pendula Roth). Данный вид обладает высокой экологической пластичностью, способностью адаптироваться к разным климатическим условиям. Так, он произрастает в Центральной Якутии, характеризующейся резко континентальным засушливым климатом с максимальными годовыми перепадами температур более 100°С и зимними минимумами -60°С. Также вид очень широко представлен в регионах с более мягким климатом, например, в Карелии. Здесь близость морей, интенсивная циклоническая деятельность обусловливают продолжительную, но не суровую зиму, относительно высокую влажность воздуха, значительное количество осадков и неустойчивые погодные условия в течение всех сезонов.
Приспособление древесных растений, имеющих многолетний цикл развития, к экстремальным факторам среды может осуществляться как в форме фенотипических изменений, так и генетических. Физиолого-биохимические показатели B. pendula относительно подробно исследованы для европейской части России [1]. Значительно в меньшей
Татаринова Татьяна Дмитриевна, кандидат биологических наук, старший научный сотрудник
Васильева Ирина Вениаминовна, аспирантка степени они изучены для Сибири и Дальнего Востока [2, 3]. Способность березы к формированию оптимального уровня морозоустойчивости в разных климатических условиях связана, наряду с другими причинами, экспрессией регулируемые холодом генов, кодирующих Cor-полипептиды. К таковым относятся водорастворимые белки-дегидрины, относящиеся ко II группе суперсемейства LEA-белков (late embryogenesis abundant – белки позднего эмбриогенеза) [4]. Они индуцируются обезвоживанием и задействованы в стабилизации макромолекул и мембран.
Цель работы: изучить сезонные изменения состава и содержания суммарных белков и дегидринов почек березы повислой ( Betula pendula Roth) при формировании морозоустойчивости в разных климатических условиях (на примере Центральной Якутии и Карелии).
Объекты и методы. Сбор почек и листьев березы B. pendula Roth проводили ежемесячно с февраля 2009 г. по февраль 2010 г. с 6-ти средневозрастных деревьев в лесопарковой зоне Ботанического сада ИБПК СО РАН (г. Якутск, 62° с.ш., 130° в.д.) и на территории экспериментальных участков ИЛ КНЦ РАН (г. Петрозаводск, 62° с.ш., 34° в.д.). Климатические показатели для Центральной Якутии (г. Якутск): среднегодовая температура воздуха – -10,2°С, среднемесячная температура воздуха в январе – -42,5°С, в июле – +19,0°С, годовое количество осадков – 215-265 мм. Климатические показатели для Карелии (г. Петрозаводск): среднегодовая температура воздуха – +2,6°С, среднемесячная температура воздуха в январе – -10,6°С, в июле – +16,2°С, годовое количество осадков –550-750 мм.
Для выделения суммарных белков из почек березы (1-2 г) использовали методику [5]. Электрофорез проводили в 13% SDS-PAGE [6]. На треки наносили равные количества белка (15 мкг). Иммуноблоттинг проводили в соответствии с методикой [7]. Белки из ПААГ переносили на ПВДФ (поливинилиденфторид) мембрану («Bio-Rad», США). Идентификацию дегидринов выполняли с помощью поликлональных антител против их консервативного К-сегмента (EKKGIME/DKIKEKLPG) [4] («Agrisera», Швеция). Дегидрины визуализировали при помощи кроличьих антител, конъюгированных с щелочной фосфатазой («Sigma», США).
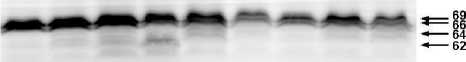
Результаты и обсуждение. Качественный спектр суммарных белков почек берез Центральной Якутии и Карелии оказался очень похожим, несмотря на то, что эти популяции обладают разными фенологическими особенностями, а также и продолжительностью глубокого и вынужденного покоя. В нем идентифицируется более 60 полипептидов с разными молекулярными массами (мол. м.). Наибольший интерес представляют белки с выраженной сезонной динамикой, представленные в летний период, а также белки, накопление которых наблюдается при подготовке растений к покою, что, по всей вероятности, связано с формированием их криорезистентности. Анализ сезонных изменений суммарных белков выявил различия между березами, произрастающими в исследуемых регионах. Так, деакклимация с повышением температуры воздуха у «якутских» берез наблюдалась в конце мая, а у «карельских» берез – в начале мая. Этот процесс сопровождался резким снижением количества белков с мол. м. 17, 26-27, 49 кДа. Накопление этих «зимних» белков в почках берез Якутии происходило в конце августа, а у «карельских» берез – только в сентябре. Вероятно, это связано с климатическими особенностями регионов – относительной большей продолжительностью вегетационного периода в Карелии по сравнению с Центральной Якутией. Вместе с тем, уровень белков в зимний период у изученных берез обеих популяций остается стабильно высоким в течение всей зимы. Дегидрины почек берез Якутии и Карелии, идентифицированные с помощью антител, в основном группируются в двух областях: низко и среднемолекулярной (рис. 1, 2).
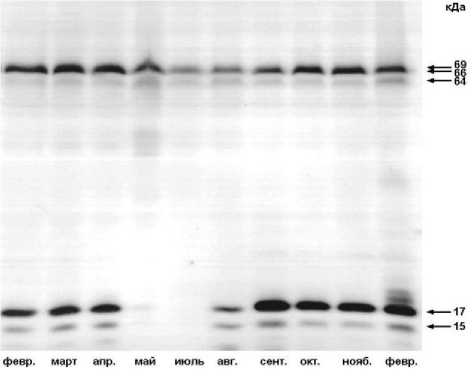
Рис. 1. Результаты иммуноблоттинга суммарных белков почек B. pendula Центральной Якутии с антителами на дегидрины (2009-2010 гг.). Справа – мол. м. дегидринов кДа
^--17 ---15
февр. март апр. май авг. сект. окт. нояб. февр.
Рис. 2. Результаты иммуноблоттинга суммарных белков почек B. pendula Карелии с антителами на дегидрины (2009-2010 гг.). Справа – мол. м. дегидринов
Анализ сезонных изменений дегидринов показал, что у всех изученных экземпляров берез круглогодично представлены белки с мол. м. 66 и 69 кДа. Низкомолекулярные дегидрины, преимущественно с мол. м. 17 кДа, имели более выраженную годовую динамику, чем среднемолекулярные. У «карельских» берез низкомолекулярные дегидрины в почках обнаруживались в сентябре, у «якутских» – в августе, сохранялись в течение всей зимы и исчезали только в начале (Карелия) или конце мая (Центральная Якутия). Количественное содержание низкомолекулярных дегидринов у берез Центральной Якутии было больше, чем у берез Карелии, а уровень среднемолекулярных дегидринов у обеих популяций был примерно одинаков.
Ранее сезонная динамика дегидринов с использованием антител против засухоспецифического полипептида dsp 16 из Craterostigma plantagineum была изучена в почках у другого вида берез (B. pubescens Ehrh.) в Финляндии [8]. У этих берез в зимний период отмечались мажорные дегидрины с мол. м. 24, 30 и 33 кДа. В Финляндии процесс накопления дегидринов завершался в ноябре, в Якутии и Карелии – в октябре. Наиболее близким к дегидрину с мол. м. 24 кДа березы B. pubescens в Фенноскандии [8] являлся дегидрин с мол. м. 21 кДа, встречающийся только у отдельных «якутских» берез. Есть также данные по исследованию двух генов дегидринов B. pubescens в Фен-носкандии. Экспрессия BpuDhn1 происходила в осенний период, а BpuDhn2 – в самые холодные месяцы зимы [9]. В контролируемых условиях при замораживании-оттаивании саженцев «финской» березы B. pendula в состоянии покоя показано, что во время оттаивания запускался синтез транскриптов дегидрина BpLTI36 для обеспечения надлежащего уровня морозоустойчивости в период частых колебаний температур [10]. Известно также о наличии кислого 36 кД-дегидрина, индуцируемого низкой температурой в листьях финской березы B. pendula [11], который в почках наших растений не обнаруживался. Наблюдаемые различия в молекулярных массах могут быть связаны с тканевой и видовой спецификой растений, а также с особенностями использовавшихся антител. Следует подчеркнуть, что в наших исследованиях применялись высокоспецифичные поликлональные антитела против консервативного К-сегмента дегидринов.
Выводы: выявлены различия в сезонной динамике суммарных белков и дегидринов березы повислой B. pendula Roth, произрастающей в Карелии и Центральной Якутии. Стабильно высокое содержание отдельных групп суммарных белков и дегидринов в зимний период может свидетельствовать об их участии в общих механизмах формирования морозоустойчивости растений в разных климатических условиях.
Работа выполнена при поддержке гранта РФФИ №09-04-98556-р_восток_а.
Список литературы Дегидрины в почках Betula pendula Roth: особенности сезонной динамики
- Ветчинникова, Л.В. Береза: вопросы изменчивости (морфо-физиологические и биохимические аспекты). -М.: Наука, 2004. 183 с.
- Бубякина, В.В. Особенности сезонной динамики дегидринов Betula platyphylla Sukacz., ассоциированные с формированием морозоустойчивости в условиях криолитозоны/В.В. Бубякина, Т.Д. Татаринова, А.Г. Пономарев и др.//Доклады Академии наук. 2011. Т. 439, № 6. С. 844-877.
- Петров, К.А. Древесные растения Якутии и низкотемпературный стресс/К.А. Петров, В.Е. Софронова, В.В. Бубякина и др.//Физиология растений. 2011. Т. 439. С. 866-874.
- Kosova, K. Role of dehydrins in plant stress response/K. Kosova, I.T. Prašil, P. Vitamvas//Handbook of plant and crop stress. -Tucson: CRC Press, 2010. P. 239-285.
- Korotaeva, N.E. Variations in the content of stress proteins in the needles of common pine (Pinus sylvestris L.) within an annual cycle/N.E. Korotaeva, M.V. Oskorbina, L.D. Kopytova et al.//Journal of Forest Research. 2011. DOI 10.1007/s10310-011-0260-y.
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of Bacteriophage T4//Nature. 1970. Vol. 227. P. 680-685.
- Timmons, T.M. Protein blotting and immunodetection/T.M. Timmons, B.S. Dunbar//Methods Enzymol. 1990. Vol. 182. P. 679-701.
- Rinne, P. Onset of freezing tolerance in birch (Betula pubescens Ehrh.) involves LEA proteins and osmoregulation and is impaired in an ABA-deficient genotype/P. Rinne, A. Welling, P. Kaikuranta//Plant Cell Environ. 1998. V. 21. P. 601-611.
- Welling, A. Photoperiod and temperature differentially regulate the expression of two dehydrin genes during overwintering of birch (Betula pubescens Ehrh.)/A. Welling, P. Rinne, A. Vihera-Aarnio et al.//J. Exp. Bot. 2004. V. 55. P. 507-516.
- Welling, A. Involvement of CBF transcription factors in winter hardiness in birch/A. Welling, E.T. Palva//Plant Physiol. 2008. V. 147. P. 1199-1211.
- Puhakainen, T. Short day potentiation of low temperature-induced gene expression of a C-repeat-binding factor-controlled gene during cold acclimation in silver birch/T. Puhakainen, Ch. Li, M. Boije-Malm et al.//Plant Physiol. 2004. V. 136. P. 4299-4307.
