Демографические особенности ценопопуляции Linum flavum (Linaceae) в урочище Ухтинские степи (Пензенская область)
 в урочище Ухтинские степи (Пензенская область)")
Автор: Фатюнина Ю.А., Артемова С.Н., Щеглов А.Е.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 2 т.17, 2023 года.
Бесплатный доступ
В урочище Ухтинские степи Linum flavum входит в состав разнотравных настоящих лугов, длиннокорневищных остепненных лугов, разнотравных луговых степей, разнотравных настоящих степей. В слабо нарушенных сообществах вид играет роль ассектатора с проективным покрытием не более 2-4%. В нарушенных разреженных сообществах его присутствие возрастает до 10-20 абс. %. Онтогенез L. flavum включает 3 периода и 9 возрастных состояний; морфогенез состоит из фазы первичного побега и первичного куста. Ценопопуляция характеризуется высокой экологической плотностью, бимодальными спектрами с высоким участием ювенильных и/или имматурных первой подгруппы особей и раннегенеративных и зрелых генеративных особей, высокой способностью к самоподдержанию численности путем семенного размножения.
Онтогенез, онтогенетическая структура, ценопопуляция, редкий вид
Короткий адрес: https://sciup.org/148326699
IDR: 148326699 | УДК: [582.736:581.55]:470.57 | DOI: 10.24412/2072-8816-2023-17-2-165-177
Demographic peculiarities of the Linum flavum (Linaceae) cenopopulation in the Ukhtinsky steppe state (Penza region)
In the tract Ukhtinskie stepi Linum flavum is a part of forb real meadows, long-rhizomatous steppe meadows, forb meadow steppes, forb real steppes. In weakly disturbed communities, it plays the role of an assector with a projective coverage of no more than 2-4%. In disturbed sparse communities, its presence increases to 10-20 abs.%. The ontogeny of L. flavum includes 3 periods and 9 age states; morphogenesis consists of the phase of the primary shoot and the primary bush. The cenopopulation is characterized by high ecological density, bimodal spectra with a high participation of juvenile and / or immature first subgroup individuals and early generative and mature generative individuals, a high ability to self-maintain the population through seed reproduction.
Текст научной статьи Демографические особенности ценопопуляции Linum flavum (Linaceae) в урочище Ухтинские степи (Пензенская область)
Linum flavum L. – редкий вид флоры европейской части России (Gubanov et al., 2003). Включен в 16 региональных Красных книг, в том числе в четырех субъектах со статусом 1 «вид под угрозой исчезновения» (Voronkina, 2015; Oktyabryova, 2018; Sokolov, Sokolova, 2019; Senator, Gafurova, 2020); в пяти субъектах со статусом 2 «сокращающийся в численности вид» (Levin, 2003; Nedosekina, 2014; Semenishchenkov, 2016; Vorotnikov, 2017; Sheremeteva, 2020), в пяти субъектах со статусом 3 «редкий вид» (Agafonov, 2011; Zernov, 2013; Teymurov, 2017; Kazakova, 2021), в том числе и в Пензенской области (Leonova, Zaplatin, 2013); в Самарской области имеет статус 5 «восстанавливающийся в численности вид» (Mitroshenkova et al., 2017); в Белгородской области – статус 6 «особо ценный вид» (Kolchanov, 2007).
Уязвимость вида в современной биосфере связывают с особенностями его экологии. Ареал этого степного и лесостепного вида резко сократился в результате распашки, палов, чрезмерного выпаса (Zernov, 2013; Teymurov, 2017). Вместе с тем, неблагоприятно сказывается на состоянии популяций и полное отсутствие выпаса с последующим олу-говением, в результате чего вид вытесняется луговыми травами и кустарниками (Ka- zakova, 2021). В северной части ареала, где вид занимает преимущественно хорошо прогреваемые южные склоны на выходах карбонатных пород, местообитаниям угрожает добыча строительных материалов (Levin, 2003; Kolchanov, 2007). Из-за декоративности генеративных особей популяции страдают от сбора растений на букеты (Nedosekina, 2014).
В Пензенской области L. flavum приурочен к ковыльным и луговым степям, сухим и остепненным лугам, лесным полянам и опушкам, предпочитая черноземные карбонатные почвы, особенно меловые субстраты (Solyanov, 1998; Novikova, 2012; Vasjukov, Saksonov, 2020). Известно 13 местообитаний (Leonova, Zaplatin, 2013). Местообитание в урочище Ухтинские степи обнаружено относительно недавно и еще не изучено. В связи с этим, цель работы – выявить демографические особенности ценопопуляции L. flavum в урочище Ухтинские степи для оценки ее состояния.
Материалы и методы
Исследование проводилось в июле – сентябре 2022 г. Изучение жизненной формы осуществлялось по методикам И.Г. Серебрякова (Serebryakov, 1962; Serebryakov, 1964) и Т.И. Серебряковой (Serebryakova, 1972). Изучение онтогенеза и описание возрастных состояний проводилось на основе метода, разработанного Т.А. Работновым (Rabotnov, 1960); дополненного А.А. Урановым (Uranov, 1975). Всего изучено около 50 растений разных возрастных состояний.
Для изучения возрастного спектра ценопопуляции были заложены 13 учетных площадей (УП) размером 2×2 м (4 м2). На каждой из них посчитано количество особей ювенильного (j), имматурного (im1, im2), виргинильного (v1, v2) и генеративного (g1, g2, g3) возрастных состояний.
Для характеристики ценопуляции использовали следующие коэффициенты: индекс возрастности Δ по А.А. Уранову (Uranov, Smirnova, 1969) с уточнениями Л.А. Животовского (Zhivotovskiy, 2001), индекс эффективности ω по Л.А. Животовскому, индекс возобновляемости, индекс генеративности (Zlobin et al., 2013).
Для определения фитоценотической приуроченности на каждой УП было выполнено полное геоботаническое описание по традиционной методике: определено общее проективное покрытие (ОПП) и проективное покрытие отдельных видов (ПП). Принадлежность фитоценоза к растительной ассоциации определяли на основании эколого-фитоценотической классификации растительности на доминантной основе (Ipatov, Mirin, 2000). Латинские названия растений приводятся в соответствии со стандартами, принятыми в базе данных International Plant Names Index (IPNI).
Урочище Ухтинские степи расположено на западной окраине с. Ухтинка (Бессонов-ский район Пензенской области), на левобережном склоне долины р. Пензятка южной экспозиции. Участок включает Ухтинский овраг и примыкающую к нему с востока залежь общей площадью около 0.5 га. Овраг представляет собой результат длительного процесса эрозии – большой объем пролювиальных отложений в конусе выноса свидетельствует о не менее чем 300-летнем возрасте оврага. На современном этапе активные эрозионные процессы протекают в средней части оврага (7 крупных отвершков – результат боковой эрозии) и на склонах оврага (крутые стенки с обнажениями). Вершины отвершков растут как сверху (эрозионный тип), так и снизу в результате вымывания мелкозема подземными водами (ендовидный тип). На склонах происходят активные оползневые процессы. В устье оврага наблюдается процесс затухания эрозии и превращения его в балку. В днище оврага есть выход грунтовых вод. В верхней части происходит зарастание склонов. Овраг расположен на землях сельскохозяйственного назначения. В верховьях на пологом склоне водораздела водосборная площадь занята пашней. Склоны речной долины в основном залужены. Приняты активные меры по борьбе с ростом оврага: в верхней части – водорегулирующее сооружение (обваловка), на склонах – лесополоса, созданная, судя по возрасту деревьев, 15–20 лет назад. В настоящее время эрозионные процессы захватывают лесополосу (эффект «пьяного леса»).
Эрозионные процессы обусловлены особенностями геологического строения речной долины р. Пензятка. Коренные породы верхнего мела (иссинская толща) сложены в основном известковистыми глинами (карбонатными). Склоны водораздела в верховьях перекрыты четвертичными отложениями – древним делювием, представленным суглинками с лессоидами и прослоями песка и щебня, играющим роль материнских пород для распространенных здесь черноземов. Присутствие мелкодисперстных частиц (лесса) способствует вымыванию их подземными водами (суффозии), что ведет к росту оврага эндовидного типа. Известковистая глина, выступает в роли водоупора, набухает и является скользящей поверхностью, что способствует оползневым процессам. Кроме того, восточная часть Приволжской возвышенности испытывает медленные процессы поднятия, вызывая вторичные врезания оврага.
В период активного земледельческого использования окружающих территорий крутые склоны оврага стали убежищем для ряда степных и луговых видов, уничтоженных при распашке остальных участков; некоторые виды, среди которых и L. flavum сейчас восстанавливают свои позиции на залежи.
Результаты и обсуждение
Фитоценотическая приуроченность . L. flavum в Ухтинских степях наиболее часто обнаруживается в составе сообществ разнотравных луговых степей (табл. 1) с доминированием льна желтого (асс. шалфеевомутовчато-желтольновая – УП № 1, асс. типчаково-желтольновая – УП № 2), асс. желтольновая – УП № 2а, УП № 11, асс . обыкновеннорепешоково-желтольновая – УП №3, асс. узколистномятликовожелтольновая – УП № 9, УП № 10) и астры итальянской (асс. тырсо-береговокострецово-итальяноастровая – УП №5, асс. тырсово-итальяноастровая – УП № 8). Кроме того, L. flavum принимает участие в формировании дерновиннозлако-вых (асс. итальяноастрово-тырсовая – УП №4) и разнотравных настоящих степей (асс. обыкновеннорепешоково-мутовчатошалфеевая – УП № 7), а также разнотравных настоящих лугов (асс. узколистномятликово-узколистнопогремковая – УП № 6) и длиннокорневищных остепненных лугов (асс. желтольново-наземновейниковая – УП № 2б).
Роль льна желтого в изученных сообществах сильно варьирует в зависимости от их строения. Число растений L. flavum на 1 м2 изменяется от 7 до 60, в среднем – 25. В слабо нарушенных выпасом луговых степях со Stipa tirsa Steven и Aster amellus L. (УП № 4, УП № 5) с ОПП 60–70%, лен желтый играет роль ассектатора, его доля не превышает 2–3%. В разнотравных настоящих степях с высоким участием Salvia verticillata L. и Agrimonia eupatoria L. (УП №7, ОПП 80%), доля L. flavum тоже низкая – 4%.
В сообществах, развивающихся на залежах, L. flavum увеличивает присутствие до 10–20%. Чем ниже сомкнутость растительного покрова, тем заметнее участие L. flavum . При уменьшении сомкнутости растительного покрова до 40% (УП № 2, УП № 3) – 20% (УП № 2а, УП № 11) доля льна желтого возрастает до трети и даже половины общего проективного покрытия.
Важно отметить, что L. flavum даже какое-то время может удерживаться в сообществах остепненных лугов с таким мощным конкурентом как Calamagrostis epigeios (L.) Roth (УП № 2б), выступая в роли содоминанта. Таким образом, на залежах разного возраста L. flavum ведет себя как содоминант и доминант, проявляя признаки эксплерент-ной стратегии.
Таблица 1. Характеристика ценопопуляции L. flavum в «Ухтинских степях»
Table 1. Characteristics of the L. flavum cenopopulation in the «Ukhtinskie stepi»
|
УП Аc cou nti ng are a |
Плот ность, шт/м2 Densit y, pcs/m 2 |
Название ассоциации – ОПП, % (доля L. flavum. , %) Association name – total projective cover, % (share of L. flavum , %) |
Абс. максимум, % Absolute maximum, % |
Δ |
ω |
Тип ЦП по клас-сифика-ции «Δ – ω» Type of cenopop-ulation by classification «Δ – ω» |
I воз, % I age % |
I ген, % I gen % |
|
1 |
24.5 |
Шалфеевомутовчато-желтольновая разнотравных луговых степей – 56 (20) |
im1 (28.6) g1 (16.3) |
0.15 |
0.33 |
Молодая |
76 |
25 |
|
2 |
31.8 |
Типчаково-желтольновая разнотравных луговых степей – 43 (14) |
j (78.7) |
0.04 |
0.13 |
Молодая |
96 |
4 |
|
2а |
55.0 |
Желтольновая разнотравных луговых степей 21 (9.5) |
j (84.5) g1 (4.0) |
0,04 |
0,14 |
Молодая |
93 |
7 |
|
2б |
59.8 |
Желтольново наземновейниковая длиннокорневищно-злаковых остепненных лугов – 45 (13) |
j (89.1) g2 (3.7) |
0.06 |
0.14 |
Молодая |
92 |
8 |
|
3 |
21.0 |
Обыкновеннорепешоково-желтольновая разнотравных луговых степей – 38 (20) |
j (58.3) g2 (23.8) |
0.15 |
0.36 |
Молодая |
69 |
31 |
|
4 |
7.0 |
Итальяноастрово-тырсовая дерновиннозлаковых луговых степей – 59 (2) |
j (46,4) |
0.09 |
0.22 |
Молодая |
89 |
11 |
|
5 |
15.0 |
Тырсо-береговокострецово-итальяноастровая разнотравных луговых степей 70 (3) |
j (88.0) g1 (10.0) |
0.07 |
0.19 |
Молодая |
87 |
13 |
|
6 |
11.5 |
Узколистномятликово- узколистнопогремковая разнотравных настоящих лугов – 40 (5) |
j (52.7) g2 (15.9) |
0.17 |
0.34 |
Молодая |
70 |
30 |
|
7 |
15.8 |
Обыкновеннорепешоково-мутовчатошалфеевая разнотравных настоящих степей – 80 (4) |
j (71.4) g2 (20.6) |
0.12 |
0.28 |
Молодая |
79 |
21 |
|
8 |
28.0 |
Тырсово-итальяноастровая разнотравных луговых степей – 55 (6) |
j (86.6) g2 (8.1) |
0.08 |
0.18 |
Молодая |
88 |
12 |
|
9 |
22.5 |
Узколистномятликовожелтольновая разнотравных луговых степей – 46 (20) |
j (74.4) g1 (8.8) |
0.08 |
0.21 |
Молодая |
84 |
16 |
|
10 |
13.8 |
Узколистномятликовожелтольновая разнотравных луговых степей 30 (7) |
j (72.7) g1 (3.6) |
0.06 |
0.15 |
Молодая |
93 |
7 |
|
11 |
18.5 |
Желтольновая разнотравных луговых степей – 25 (10) |
j (68.9) g1 (13.5) |
0.10 |
0.23 |
Молодая |
80 |
20 |
Особенности онтогенеза . L. flavum – это многолетнее стержнекорневое каудексное травянистое растение. В урочище Ухтинские степи было выделено 3 возрастных периода и 9 возрастных состояний (табл. 2).
Таблица 2. Онтогенез L. flavum в урочище «Ухтинские степи»
Table 2. Ontogeny of L. flavum in the tract «Ukhtinskie stepi»
|
Признак Feature |
j |
im1 |
im2 |
v |
g1 |
g2 |
g3 |
|
Высота вегетативного побега, см |
1.5 |
6.6 |
12.8 |
21.4 |
15.7 |
18.9 |
– |
|
Vegetative shoot height, cm |
±0.3 |
±0.8 |
±0.4 |
±0.4 |
±0.6 |
±1.6 |
|
|
Высота генеративного побега, см Height of generative shoot, cm |
— |
— |
37.7 ±1.7 |
48.2 ±1.6 |
33.6 ±1.2 |
||
|
Число вегетативных побегов, шт. |
1.0 |
1.0 |
2.3 |
1.6 |
2.5 |
1.7 |
— |
|
Number of vegetative shoots, pcs. |
±0.0 |
±0.0 |
±0.2 |
±0.4 |
±0.3 |
±0.6 |
|
|
Число генеративных побегов, шт. Number of generative shoots, pcs. |
— |
— |
— |
— |
1.2 ±0.2 |
7.9 ±2.9 |
1.8 ±0.4 |
|
Число листьев на побеге*, шт. |
3.5 |
9.6 |
12.6 |
17.6 |
25.7 |
27.2 |
26.4 |
|
Number of leaves on the shoot*, pcs. |
±0.5 |
±1.1 |
±0.6 |
±1.2 |
±0.8 |
±0.7 |
±1.0 |
|
Число листьев на растении*, шт. |
3.5 |
9.6± |
29.5 |
35.1 |
40.0 |
253.4 |
49.6 |
|
Number of leaves on the plant*, pcs. |
±0.5 |
1.1 |
±3.2 |
±6.5 |
±3.7 |
±77.7 |
±9.5 |
|
Длина листа, см |
0.6 |
1.4 |
2.7 |
3.8 |
3.7 |
3.3 |
2.9± |
|
Sheet length, cm |
±0.1 |
±0.1 |
±0.1 |
±0.2 |
±0,2 |
±0,1 |
0.1 |
|
Ширина листа, мм |
3.0 |
4.6 |
8.0± |
10.4 |
8.3 |
7.4 |
6.3 |
|
Sheet width, mm |
±0.1 |
±0.3 |
0.3 |
±0.3 |
±0.2 |
±0.2 |
±0.4 |
|
Число боковых корней, шт. |
10.0 |
12.0 |
13.0 |
13.0 |
15.0 |
17.0 |
14.0 |
|
Number of lateral roots, pcs. |
±1.0 |
±3.0 |
±3.0 |
±2.5 |
±1.2 |
±3.2 |
±1.1 |
|
Длина корня, см |
4.0 |
10.0 |
12.3 |
17.2 |
19.0 |
20.0 |
21.0 |
|
Root length, cm |
±0.8 |
±1.0 |
±0.8 |
±0.8 |
±2.1 |
±1.2 |
±1.9 |
|
Число цветков на побеге, шт. Number of flowers on the shoot, pcs. |
– |
– |
– |
– |
18.0 ±1.9 |
17.9 ±2.0 |
18.2 ±1.6 |
|
Число цветков на растении, шт. Number of flowers on the plant, pcs. |
— |
— |
— |
— |
23.1 ±4.2 |
183.9 ±38.5 |
35.3 ±3.2 |
-
* для генеративных особей – без учета вегетативных побегов
-
* for generative individuals – excluding vegetative shoots
Латентный период представлен семенем. Семена 0.3–0.4 см в длину, 0.1–0.2 см в ширину, яйцевидной форм, с блестящей поверхностью, гладкие; окраска темнокоричневая, к краям переходящая в светло-бурую или желтую.
Прегенеративный период . Для вида характерно надземное прорастание. В период наблюдений проростки отсутствовали, так как уже достигли следующего – ювенильного – возрастного состояния. J-особь представляет собой удлиненный побег высотой до 2 см, в том числе эпикотиль 4–5 мм, гипокотиль 15 мм. Семядоли 2–3 мм в длину и 1 мм в ширину; во вторую декаду июля желтые или бурые сухие. Настоящих листьев 3–5, до 6 мм в длину, до 3 мм в ширину. Подземные органы представлены стержневой корневой системой; главный корень длиной 5 см, диаметром до 0.5 мм. Боковые корни тонкие, в числе 9–12. Продолжительность j-состояния 1 год. На следующий год растения переходят к имматурному состоянию, либо отмирают.
Имматурные особи ясно дифференцируются на 2 подгруппы: im1 – однопобеговые растения высотой 4.5–9.5 см, с 8–14 листьями длиной 12–16 мм, шириной 4–5 мм. По- бег развивается из почки на гипокотиле, нарастание симподиальное. Начинает формироваться каудекс – главный корень сокращается и втягивает основание побега в почву, закладываются почки возобновления. Возраст особей ясно определяется по числу пере-вершиниваний. Продолжительность этого этапа онтогенеза – от 1–2 года до 4 лет в условиях затенения. С началом ветвления растения переходят к im2-возрастному состоянию. Ежегодно трогаются в рост 2–3 почки на каудексе, следовательно, число побегов увеличивается до 2–3. Продолжительность этого состояния определить труднее, видимо, от 1 до 2–3-х лет. Высота побегов увеличивается до 9.0–14.5 см. По числу листьев на каждом побеге (8–17) im2 -особи практически не отличаются от im1-растений, но листья крупнее – длиной 2.3–3.0 см, шириной 7–9 мм. Главный корень удлиняется до 15 см. Некоторые исследователи не видят оснований для дифференцировки имма-турного возрастного состояние на подгруппы (Klimachyeva, 2018), но, по нашему мнению, это оправдано.
Виргинильные растения формируют от 1 (редко, обусловлено пониженной жизненностью) до 4 побегов высотой до 17.5–23.0 см, количество листьев на побеге возрастает до 11–23, размеры листа увеличиваются до 3.0–5.0 см в длину и 9–12 мм в ширину. Продолжительность состояния от нескольких месяцев до нескольких лет. Таким образом, в наиболее благоприятных условиях лен желтый на третий год с момента прорастания семени может перейти к генеративному периоду онтогенеза.
Генеративные особи ясно дифференцируются на 3 подгруппы. У раннегенеративных растений 1–2 генеративных побега высотой 28–45 см и 1–4 вегетативных побега вирги-нильного облика высотой 9–17 см. Диаметр стебля составляет 1–2 мм. Каудекс небольшой, диаметром до 1.5–3.0 мм, с небольшим числом следов отмерших прошлогодних побегов. Число листьев в префлоральной части генеративных побегов составляет от 22 до 30. Длина листа варьирует от 3.3 до 4.4 см, ширина – от 7 до 10 мм. На генеративных побегах листья несколько мельче, чем на виргинильных. Число цветков на побеге колеблется от 8 до 25, на растении от 8 до 38.
У g2-особей вегетативные побеги встречаются намного реже, их число всегда намного меньше, чем генеративных побегов. Количество генеративных побегов варьирует от 4 до 25, высота – от 29 до 59 см. Диаметр стебля несколько увеличивается по сравнению с g1 до 1.5–3.0 мм. Диаметр каудекса – от 4.5 до 8.0 мм. По облиственности генеративного побега в префлоральной части g1- и g2-особи достоверно не отличаются, но из-за увеличения числа побегов облиственность особи возрастает до 106–712 листьев. Длина листа варьируют значительно больше, чем у g1-особей, но средние значения близки. По среднему числу цветков на побеге g1- и g2-особи достоверно не отличаются (при большем диапазоне изменчивости у g2-особей – от 5 до 72). Но из-за значительного увеличения числа побегов общее число цветков на растении существенно возрастает до 75–378 (в среднем почти в 8 раз). Это период максимального расцвета растения и его наибольшей семенной продуктивности. По мере старения, растение переходит к позднегенеративному состоянию.
П озднегенеративные особи обнаруживают сходство g1-особями по числу генеративных побегов (1–2), их высоте (22–39 см), диаметру стебля (1–2 мм), облиственности особи (25–109), по числу цветков у одной особи (10–30). Однако есть 2 существенных отличия, по которым их можно дифференцировать без выкапывания. У g3-особей отсутствуют вегетативные побеги. Кроме того, g3-особь имеет вид более рыхлого куста (в отличие от компактного у g1-особей) за счет того, что побеги на каудексе формируются только по периферии, а его центральная часть несет многочисленные следы отмерших побегов. Диаметр каудекса всегда больше 10 мм. Партикуляция, свойственная другим видам данной жизненной формы в этом возрасте, нами не была отмечена, видимо, из-за в целом небольшого диаметра каудекса и малочисленности побегов.
Растения на постгенеративном этапе развития, обычно свойственные каудексным поликарпикам (Zaugolnova, Smirnova, 1978; Zhukova, 1995), нами обнаружены не были, хотя самарские исследователи обнаружили в небольшом количестве субсенильные особи (Klimachyeva, 2018).
Онтогенетический анализ показал (рис. 1), что онтогенетические спектры L. flavum двувершинные. Первый максимум наблюдается на j- и, отчасти, на im1-особях. Число j-особей изменяется от 3 до 53 экз./м2 – (в среднем 18 экз./м2), что составляет от 26 до 89 % особей (в среднем 69 %). Это связано с большим количеством мелких семян, которые ежегодно образуются и прорастают, особенно укрытые степным войлоком. Число im1-особей уже заметно ниже – 0–7, в среднем 1,8 экз./м2, или 0.0–15.9 %, (за исключением УП №1 и УП №4, где участие im1-особей остается значительным – до 28.6– 39.2%). Таким образом, в среднем на долю im1-растений приходится 10.3%.
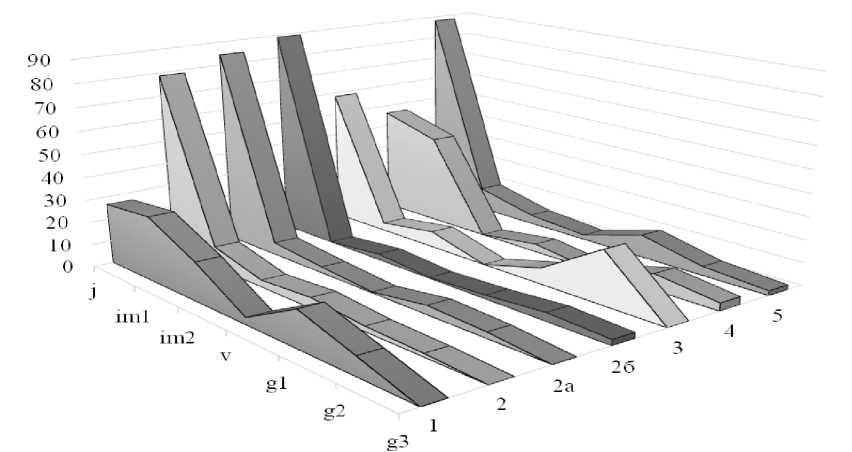
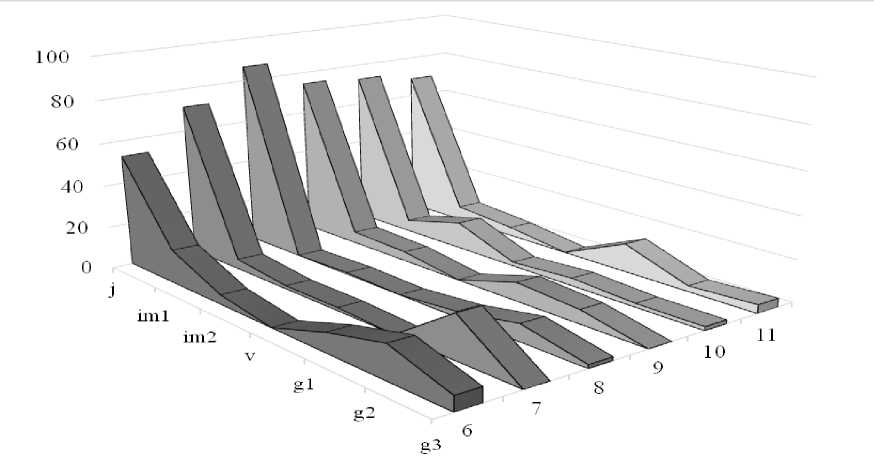
Рис. Онтогенетические спектры L. flavum в «Ухтинских степях», %.
Fig. Ontogenetic spectra of L. flavum in the «Ukhtinskie stepi»,%
Число im2-особей также снижается до 0.0–4.3 экз./м2 (в среднем 1,1 экз./м2) – то есть до 0–17 % (в среднем 5%). Наименее многочисленна группа v-растений – до 0.0– 1.5 экз./м2 (в среднем 0,3 экз./м2); v-особи не обнаружены на 5 площадках из 13-ти изученных. Их вклад в структуру популяции – от 0 до 6% (в среднем 1.3 %).
Второй максимум наблюдается на g1- (5 учетных площадей) или g2-особях (8 учетных площадей). Это связано с увеличением продолжительности пребывания в этом состоянии по сравнению с особями прегенеративного периода.
Число g1-особей изменяется от 0 до 4 экз./м2 (в среднем 1.4 экз./м2), или 0.0–16.3 % (в среднем 6.1 %). Число g2-особей – от 0.3 до 5 экз./м2 (в среднем 1,6 экз./м2), их доля варьирует в диапазоне от 1.5 до 23.8 % (в среднем 8.2 %).
Анализ онтогенетических спектров позволяет оценить состояние популяции (Zhukova, 1967; Zaugolnova, 1976; Smirnova et al., 1976; Populyatsionnaya …, 1994; Smirnova, 2004; Osmanova, Zhivotovskiy, 2020). Сопоставляя полученные нами результаты с данными самарских экологов, можно прийти к выводу, что онтогенетические спектры льна желтого в Ухтинских степях кардинально отличаются от таковых в «Самарской Луке», где наблюдаются одновершинные спектры с максимумом на g2-особях (31.8 %), при значительном участии g3- и g1-особей (25.6 и 18.3 % соответственно), заметной ролью v-растений (17.7%) и очень скромным вкладом в структуру популяции проростков, ювенильных и имматурных особей (1.3, 1.0 и 2.4 % соответственно) (Klimachyeva, 2018). Так как авторы больше не приводят никаких характеристик популяции (плотности популяции, фитоценотической приуроченности), то интерпретировать эти различия трудно. Возможно, низкая доля проростков и ювенильных особей связана с высокой сомкнутостью растительных сообществ, значительной конкуренцией, отсутствием нарушений по сравнению с «Ухтинскими степями», где постоянно образующиеся нарушения почвенно-растительного покрова снижают конкуренцию и создают условия для приживаемости семян. Также благоприятны и зоогенные нарушения, создаваемые муравьями. Хотя это не объясняет большой доли ювенильных особей на ненарушенных хорошо задернованных участках Ухтинских степей (УП№4, УП№5). Еще одной возможной причиной может быть флуктуационная динамика, поэтому требуются дальнейшие наблюдения.
По классификации «Δ – ω» ценопопуляция L. flavum в урочище Ухтинские степи принадлежит к группе молодых (показатель Δ варьирует от 0.04 до 0.17, в среднем 0.09; показатель ω – от 0.13 до 0.36, в среднем 0.22). Об этом же свидетельствуют и высокие индексы возобновляемости (69–96%, в среднем 84%) в сочетании с низкими индексами генеративности (4–31%, в среднем 16%) (табл. 1).
Заключение
Таким образом, онтогенез L. flavum в Ухтинских степях включает 3 периода и 9 возрастных состояний; постгенеративный период онтогенеза не выражен. Морфогенез L. flavum как каудексного поликарпика состоит из двух фаз: первичного побега (онтогенетические состояния: p, j, im1) и первичного куста (онтогенетические состояния: im2, v, g1, g2, g3). Вид имеет моноцентрическую биоморфу, так как размножается только семенами, вегетативно неподвижен.
L. flavum в Ухтинских степях входит в состав разнообразных сообществ от разнотравных настоящих лугов, длиннокорневищных остепненных лугов до разнотравных луговых и настоящих степей. В слабо нарушенных сообществах он играет роль ассек-татора с ПП не более 2–4%. В нарушенных разреженных сообществах его присутствие возрастает до 10–20 абс.%, L. flavum играет роль содоминанта и доминанта, демонстрируя, таким образом, признаки эксплерентной стратегии.
Ценопопуляция L. flavum в Ухтинских степях характеризуется высокой экологической плотностью. Анализ онтогенетической структуры (бимодальные спектры с высоким участием j- и/или im1-особей с одной стороны и g1- и g2- особей с другой стороны) и показателей возрастности, эффективности, возобновляемости и генеративности позволяет говорить о высокой способности к самоподдержанию численности ценопопуля-ции, о ее стабильном состоянии. Геолого-геоморфологические условия (карбонатные почвогрунты, высокая эрозионная расчлененность) благоприятны для сохранения ареала распространения вида. Антропогенная нагрузка умеренная: территория не используется для выпаса; сенокошение практически отсутствует; отмечена несанкционированная свалка бытовых отходов; заметна небольшая рекреационная нагрузка; осуществляется сбор плодов клубники, сбор растений на букеты, но в целом явной угрозы существованию ценопопуляции в настоящий момент не обнаружено. Однако вид нуждается в постоянном мониторинге, так как ценопопуляция расположена в непосредственной близости от активно расстраивающегося населенного пункта.
Авторы благодарят доктора биологических наук, профессора кафедры общей биологии и биохимии Педагогического института имени В.Г. Белинского Любовь Александровну Новикову, а также биолога-любителя, энтузиаста в изучении флоры Пензенской области Сергея Шитова.
Список литературы Демографические особенности ценопопуляции Linum flavum (Linaceae) в урочище Ухтинские степи (Пензенская область)
- [Agafonov] Агафонов В.А. 2011. Лен желтый. — В кн.: Красная книга Воронежской области. Воронеж. С. 162-163.
- [Gubanov et al.] Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. 2003. Иллюстрированный определитель растений Средней России. Том 2: Покрытосеменные (двудольные раздельнолепестные). М. С. 513.
- [Ipatov, Mirin] Ипатов В.С., Мирин Д.М. 2000. Описание фитоценоза: методические рекомендации. СПб. 55 с.
- [Kazakova] Казакова М.В. 2021. Лен желтый. — В кн.: Красная книга Рязанской области. Ижевск. С. 377.
- [Klimachyeva] Климачева Е.А. 2018. Онтогенетическая структура ценотических популяций некоторых редких растений Яблоневой геосистемы (Самарская Лука). — В кн.: Самарская Лука: проблемы региональной и глобальной экологии. Т. 27. № 4(1). С.266-270.
- [Kolchanov] Колчанов Р.А. 2007. Лен желтый. — В кн.: Красная книга Белгородской области. Белгород. С. 172.
- [Levin] Левин В.К. 2003. Лен желтый. — В кн.: Красная книга Республики Мордовия. Редкие виды растений, лишайников и грибов. Саранск. С. 152.
- [Leonova, Zaplatin] Леонова Н.А. Заплатин П.И. 2013. Лен желтый. — В кн.: Красная книга Пензенской области. Грибы, лишайники, мхи и сосудистые растения. Пенза. Т.3. С. 181.
- [Mitroshenkova et al.] Митрошенкова А.Е., Ильина В.Н., Сенатор С.А. 2017. Лен желтый. — В кн.: Красная книга Самарской области. Т. 1. Редкие виды растений и грибов. Самара. С. 156.
- [Nedosekina] Недосекина Т.В. 2014. Лен желтый. — В кн.: Красная книга Липецкой области. Растения, грибы, лишайники. Липецк. С. 368-369.
- [Novikova] Новикова Л.А. 2012. Структура и динамика травяной растительности лесостепной зоны на западных склонах Приволжской возвышенности и пути ее оптимизации. Дис. ... доктора биол. наук. Пенза. 537 с.
- [Oktyabryova] Октябрева Н.Б. 2018. Лен желтый. — В кн.: Красная книга Московской области. Московская область. С. 593.
- [Osmanova, Zhivotovskiy] Османова Г.О., Животовский Л.А. 2020. Онтогенетический спектр как индикатор состояния ценопопуляций растений. — Известия РАН. Серия Биологическая. 2:144-152.
- [Populyatsionnaya Ekologia rasteniy] Популяционная экология растений. 1994. Йошкар-Ола. 88 с.
- [Rabotnov] Работнов Т.А. 1960. Методы определения возраста и длительности жизни у травянистых растений. — В кн.: Полевая геоботаника. М.-Л. Т. 2. С. 141-149.
- [Semenishchenkov] Семенищенков Ю.А., Панасенко Н.Н. 2016. Лен желтый. — В кн.: Красная книга Брянской области. Брянск. С. 102.
- [Senator, Gafurova] Сенатор С.А., Гафурова М.М. 2020. Лен желтый. — В кн.: Красная книга Чувашской Республики. Том 1. Часть 1. Редкие виды растений и грибов. М. С. 112.
- [Serebryakov] Серебряков И.Г. 1962. Экологическая морфология растений. М. 378 с.
- [Serebryakov] Серебряков И.Г. 1964. Жизненные формы высших растений и их изучение. — В кн.: Полевая геоботаника. Т. 3. Л. С. 146-205.
- [Serebryakova] Серебрякова Т.И. 1972. Учение о жизненных формах растений на современном этапе. — В кн.: Итоги науки и техники. Ботаника. Т.1. С. 84-169.
- [Sheremeteva] Шереметьева И.С. 2020. Лен желтый. — В кн.: Красная книга Тульской области. Растения и грибы. Тула. С. 125-126.
- [Smirnova] Смирнова О.В. 2004. Оценка состояния популяции по типу онтогенетического спектра. — В кн.: Восточноевропейские леса: история в голоцене и современность. М. Т. 1. С. 159-161.
- [Smirnova et al.] Смирнова О.В., Заугольнова Л.Б., Ермакова И.М. 1976. Ценопопу-ляции растений (основные понятия и структура). М. 217 с.
- [Sokolov, Sokolova] Соколов А.С., Соколова Л.А. 2019. Лен желтый. — В кн.: Красная книга Тамбовской области: мхи, сосудистые растения, грибы, лишайники. Тамбов. С.146-147
- [Solyanov] Солянов А.А. 2001. Флора Пензенской области. Пенза. С. 80.
- [Teymurov] Теймуров А.А. 2017. Лен желтый. — В кн.: Красная книга Курской области: редкие и исчезающие виды животных, растений и грибов. Калининград; Курск. С. 218.
- [Uranov] Уранов А.А. 1975. Возрастной спектр ценопопуляций как функция времени и энергетических волновых процессов. — Биол. науки. 2: 7-34.
- [Uranov, Smirnova] Уранов А.А., Смирнова О.В. 1969. Классификация и основные черты развития популяций многолетних растений. — Бюлл. МОИП. Отд. биол. 79(1): 119-135.
- [Vasjukov, Saksonov] Васюков В.М., Саксонов С.В. 2020. Конспект флоры Пензенской области. Флора Волжского бассейна. Т. IV. Тольятти. С. 128.
- [Voronkina] Воронкина Н.В. 2015. Лен желтый. — В кн.: Красная книга Калужской области. Т. 1. Растительный мир. Калуга. С. 342.
- [Vorotnikov] Воротников В.П. 2017. Лен желтый. — В кн.: Красная книга Нижегородской области. Т. II. Сосудистые растения, моховидные, водоросли, лишайники и грибы. Калининград. С. 93.
- [Zaugolnova] Заугольнова Л.Б. 1976. Неоднородность строения ценопопуляций во времени и пространстве. — Бот. журн. 61(2): 187-196.
- [Zaugolnova, Smirnova] Заугольнова Л.Б., Смирнова О.В. 1978. Возрастная структура ценопопуляций многолетних растений и ее динамика. — Журнал общей биологии. 39(6): 849-857.
- [Zernov] Зернов А.С. 2013. Лен желтый. — В кн.: Красная книга Карачаево-Черкесской Республики. Черкесск. С. 102.
- [Zlobin et al.] Злобин Ю.А., Скляр В.Г., Клименко А.А. 2013. Популяции редких видов растений: теоретические основы и методика изучения. Сумы. 439 с.
- [Zhivotovskiy] Животовский Л.А. 2001.Онтогенетические состояния, эффективная плотность и классификация популяций растений. — Экология. 21: 3-7.
- Жукова Л.А. 1967. Изменение возрастного состава популяций луговика дернистого на окских лугах при различной продолжительности выпаса. — Биологические науки. 7: 67-72.
- Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола. 224 с.
