Демографические показатели ценопопуляций Helleborus caucasicus A. Br. на территории российской части Кавказа и Республики Абхазия

Автор: Карамурзова Марьят Мухамедовна, Шхагапсоев Сафарби Хасанбиевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 1, 2018 года.
Бесплатный доступ
На территории ряда регионов Кавказа (Ставропольский край, Республика Адыгея, Кабардино-Балкарская Республика, Абхазия) исследована возрастная структура ценопопуляций редкого вида H. caucasicus. Выявлены биологические свойства вида, определяющие характер базового возрастного спектра. Установлен характер реакции возрастных спектров ценопопуляций на антропогенное воздействие. Показана способность H. caucasicus захватывать незанятые участки, а также успешность реинтродукции вида в лесных фитоценозах.
Онтогенетический спектр, возрастная структура, эффективность самоподдержания
Короткий адрес: https://sciup.org/146279482
IDR: 146279482 | УДК: 581.5;
Demographic indicators of Helleborus caucasicus A. Br. cenopopulations within the Russian part of Caucasus and Abkhazia
The age structure of populations of the rare species H. caucasicus has been investigated within the territory of the Caucasus Regions (Stavropol Region, Adygea, Republic of Kabardino-Balkaria, Abkhazia). Biological properties of species, determining the character of base age spectrum have been established. Age population spectra change in response to anthropogenic influence. H. caucasicus is able to capture free territory. Species was successfully reintroduced in forest plant communities.
Текст научной статьи Демографические показатели ценопопуляций Helleborus caucasicus A. Br. на территории российской части Кавказа и Республики Абхазия
Введение. Проведение фундаментальных исследований экологобиологических особенностей дикорастущих видов актуально как одно из направлений в реализации концепций рационального природопользования и сохранения биоразнообразия. Поэтому целью нашей работы стало изучение возрастной структуры Helleborus caucasicus А. Вг. - редкого эндемичного ранневесеннего кавказского вида семейства Ranunculaceae Juss., занесенного в региональные Красные книги, обладающего ценными декоративными признаками и лекарственными свойствами.
Анализ исторических аспектов изученности вида Н. caucasicus позволяет сделать вывод о широкой представленности в литературе данных по его распространению (Зернов, 2006; Олексин, Рыбалко, 2015; и др.), морфологии (Гулиа, Челомбитько, 2003; Гулиа, Орловская, 2014; и др.), экологической и фитоценотической приуроченности (Хасуева, Астамирова, Теймуров, 2008; Тайсумов, 2012; и др.), практическому применению (Гусева, 2015; Олексин, Рыбалко, 2015; и др.). В гораздо меньшей степени (Загурная, Алексеева, 2010; Гулия, Орловская, 2015а,б) представлены результаты исследований структурно- функциональных особенностей ценопопуляций, в том числе возрастной структуры.
Методика. В ходе исследований нами проведен сравнительный анализ онтогенетической структуры 10 ценопопуляций (ЦП) Н. Caucasians, произрастающих в различных эколого-фитоценотических условиях на территории Кабардино-Балкарской Республики (ЦП1, ЦП2, ЦПЗ, ЦП4), Республики Адыгея (ЦП7), Абхазии (ЦП9), Ставропольского (ЦП11) и Краснодарского (ЦП 13, ЦП 14, ЦП 15) краев, в естественных и приближенных к естественным местообитаниях. Кроме того, в 2016 году изучена возрастная структура ЦП17 и ЦП18, реинтродуцированных нами в 2011 г. на территории Белореченского и Урванского лесничеств Кабардино-Балкарии (высадка генеративных растений), а также одичавшей ЦП16 в окр. с. Адиюх.
Прослежена также динамика возрастной структуры четырех ЦП вида (ЦП1, ЦП2, ЦПЗ, ЦП4) в условиях Кабардино-Балкарской Республики в течение пяти лет (с 2008 по 2012 гг.).
Описание возрастных состояний (проростки pl, имматурные im, виргинильные v, генеративные (gl, g2, g3), субсенильные ss и сенильные s) проводили по методике Т.А. Работнова (1992) и А.А. Уранова (1975). Возрастные типы ЦП выделяли по классификаци «А-со» Л. А. Животовского (2001), Онтогенетическую структуру ЦП анализировали с использованием ряда демографических показателей: индексы замещения 1з и восстановления 1в (Жукова, 1987), индекс старения 1с (Глотов, 1998), эффективная плотность Ме (Животовский, 2001). Первичный материал обработан при использовании программ BIOSTAT, EXCEL. Латинские названия таксонов приведены по С.К. Черепанову (1995).
Результаты и обсуждение. Характерный (базовый) возрастной спектр Н. Caucasians, позволяющий выделить общие закономерности, повторяющиеся в возрастной структуре отдельных ЦП, широкий, двумодальный - с пиками на виргинильной (27,30%) и генеративной (29,87%) группах, низкой долей особей постгенеративного периода (2,57% субсенильных и 1,63% сенильных растений) (рис. 1).
Повышение в базовом спектре доли виргинильных особей во многом связано с биологическими особенностями вида - активным вегетативным размножением в форме юношеской партикуляции (в прегенеративном периоде) и зрелой партикуляции (в генеративном периоде) с омоложением рамет до виргинильного состояния.
Вероятно, значение имеет также замедленное развитие прегенеративных растений при взаимном угнетении в моноцентрических плотных партикулах («правило Сукачева» (Работнов, 1992)). Кроме того, задержка развития особей Н. Caucasians на этапе подготовки к формированию генеративного побега, как и у других многолетников, может быть связана со значительными «репродуктивными тратами». По мнению Т.А. Работного (1950), растения замедляют развитие, чтобы накопить ресурсы для развития генеративных органов.
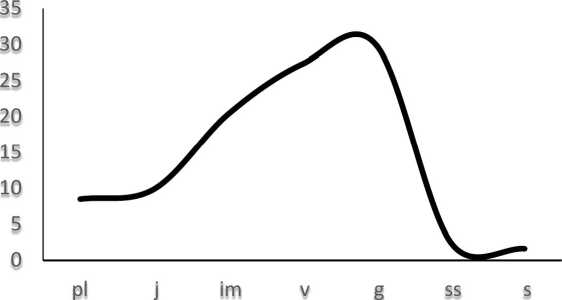
Рис. 1. Базовый возрастной спектр Н. caucasicus.
По оси ох - возрастные состояния (pl-s), по оси оу - доля возрастной группы в спектре.
Высокий процент генеративных растений в характерном возрастном спектре может определяться продолжительностью репродуктивного периода, а незначительная в целом доля особей постгенеративного периода, вероятно, связана с их быстрым отмиранием.
В то же время, исследования показали, что особенности возрастной структуры отдельных ЦП Н. caucasicus в значительной степени определяются степенью и характером антропогенной нагрузки.
Так, в условиях низкого (ЦП7, ЦП10, ЦП14) и среднего (ЦП11, ЦП15, ЦП16) уровня антропогенного давления (отсутствие или слабая выраженность переуплотнения почвы, относительная ненарушенность лесной подстилки, отсутствие следов выпаса скота в лесном массиве, редкие случаи рубки леса, обрыва цветоносов и т.п.) под пологом густого леса отмечено значительное увеличение доли особей генеративного периода (27,89-42,12%) (рис. 2).
Это обусловлено, вероятно, относительно быстрыми темпами развития прегенеративных (имматурных и виргинильных) растений в благоприятных эколого-фитоценотических условиях, а также отсутствием сбора генеративных растений на букеты. Увеличение в возрастных спектрах представленности генеративых особей при снижении доли имматурных (6,8 8-20,52%) и виргинильных (20,2428,48%) растений приводит к уменьшению показателей самоподдержания ЦП - индексов восстановления 1в и замещения 1з (1,35-4,12 и 1,02-3,68 соответственно) (табл. 1). Индексы восстановления 1в и замещения 1з показывают, соответственно, какую часть генеративных особей или генеративных и старых растений могут заменить молодые прегенеративные особи (Жукова, 1987).
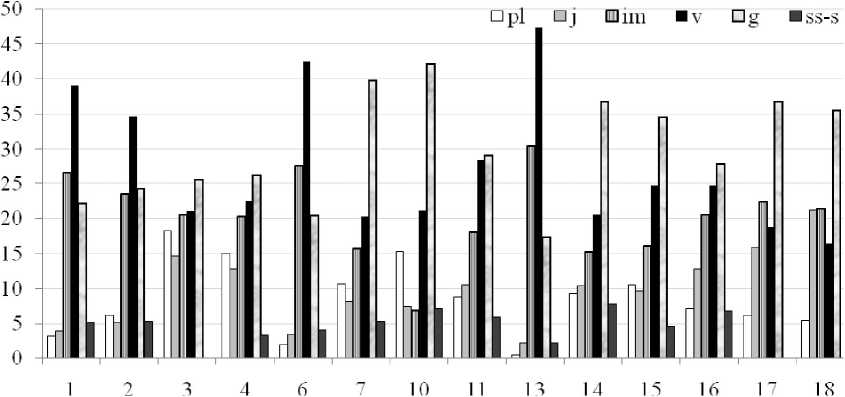
Рис. 2 . Возрастные спектры ценопопуляций Н. caucasicus. По оси ох -порядковые номера ЦП, по оси оу - доля возрастной группы в спектре, %.
Таблица 1
Демографические показатели ценопопуляций Н. caucasicus
|
№ ЦП |
1в |
1з |
1с |
м |
Ме |
А |
о |
Тип ЦП |
|
ЦП в условиях пониженного ант |
ропогенного давления |
|||||||
|
7 |
2,07 |
1,62 |
0,11 |
15,63 |
9,85 |
0,35 |
0,63 |
Зреющая |
|
10 |
1,35 |
1,02 |
0,15 |
22,07 |
14,34 |
0,37 |
0,65 |
Зрелая |
|
11 |
3,45 |
3,21 |
0,12 |
10,84 |
6,29 |
0,30 |
0,58 |
Молодая |
|
14 |
2,68 |
2,11 |
0,17 |
17,87 |
11,08 |
0,33 |
0,62 |
Зреющая |
|
15 |
3,02 |
2,89 |
0,09 |
11,85 |
7,11 |
0,31 |
0,60 |
Зреющая |
|
16 |
4,12 |
3,68 |
0,14 |
9,16 |
5,22 |
0,28 |
0,57 |
Молодая |
|
ЦП в условиях высокого антропогенного давления |
||||||||
|
1 |
6,35 |
5,62 |
0,10 |
5,87 |
3,17 |
0,24 |
0,54 |
Молодая |
|
2 |
5,62 |
5,02 |
0,07 |
8,34 |
4,59 |
0,26 |
0,55 |
Молодая |
|
6 |
7,34 |
6,78 |
0,05 |
5,56 |
2,89 |
0,23 |
0,52 |
Молодая |
|
13 |
9,34 |
8,79 |
0,02 |
3,76 |
1,88 |
0,21 |
0,50 |
Молодая |
|
ЦП на заброшенных лесных участках ботанических садов |
||||||||
|
3 |
6,48 |
6,48 |
- |
10,35 |
5,38 |
0,22 |
0,52 |
Молодая |
|
4 |
5,44 |
5,23 |
0,04 |
8,67 |
4,59 |
0,23 |
0,53 |
Молодая |
|
ЦП, реинтродуцированные в 2011 г. |
||||||||
|
17 |
8,54 |
8,54 |
- |
2,48 |
1,41 |
0,34 |
0,57 |
Молодая |
|
18 |
8,73 |
8,73 |
- |
1,34 |
0,75 |
0,32 |
0,56 |
Молодая |
Примечание: 1в, 1з, 1с - индексы восстановления, замещения, старения, М -физическая плотность, осб./м1, Ме - эффективная плотность, осб./м2; А - индекс возрастности, со - индекс эффективности, тип ЦП определен по классификации «Д-со».
В то же время, наличие собственных источников семян (генеративных растений) и высокая приживаемость проростков на ненарушенных и не переуплотненных почвах определяют высокую долю в спектрах как проростков (7,16-15,23%), так и ювенильных (7,43-
12,83%) растений, что свидетельствует об интенсивных процессах семенного возобновления ЦП. Наряду с активным вегетативным размножением и высокой долей генеративных растений это определяет максимальное повышение показателей физической (9,16-22,07 осб./м2) и эффективной (5,22-14,34 осб./м2) плотности растений, характеризующих интенсивность воздействия ЦП на ресурсы биотопа. Значительное участие в онтогенетических спектрах растений постгенеративного периода (4,58-7,17%) и относительно высокие показатели индекса старения 1с (0,09-0,17), вероятно, являются следствием пополнения субсенильной и сенильной групп за счет взаимного угнетения и быстрого старения генеративных растений в плотных клонах.
Таким образом, ЦП7, ЦП10, ЦП11, ЦП14, ЦП15, ЦП16 Н. caucasicus, развивающиеся в пределах ненарушенных лесных фитоценозов, полночленные (представлены все возрастные группы), генеративно-ориентированные (с абсолютным максимумом на группе генеративных растений). По классификации возрастных типов «дельтаомега», ЦП7, ЦП14, ЦП15 относятся к зреющим, ЦП10 - зрелая, ЦП11 и ЦП 16 — молодые.
В природных ЦП, произрастающих в условиях повышенного антропогенного давления в форме вырубки леса, вытаптывания, выпаса скота, сбора на букеты, заготовки корневищ (ЦП1, ЦП2, ЦП6, ЦП 13), наблюдается массовая гибель проростков (всего 0,46-6,17%), наименее устойчивых (наряду со старыми растениями) к переуплотнению почвы и механическим повреждениям. Это определяет, в свою очередь, низкую долю ювенильных особей в возрастных спектрах (2,24-5,15%). Медленные темпы прохождения ранних этапов онтогенеза в прегенеративном периоде и выраженная задержка в развитии на этапе формирования генеративных органов способствуют накоплению в спектрах имматурных (23,52-30,42%) и виргинильных (34,61-47,32%) особей, снижению процента генеративных растений (17,34-24,24%). В то же время, несоответствие условий произрастания биологическим требованиям вида, вероятно, способствует быстрому отмиранию генеративных, субсенильных и сенильных растений, что определяет уменьшение доли особей постгенеративного периода (2,22-5,35%) и индекса старения 1с (0,02-0,10). Слабое семенное возобновление, низкая интенсивность вегетативного размножения (в условиях переуплотнения почвы), наряду со сбором растений, приводят к резкому сокращению физической плотности особей (3,76-8,34 осб./м2). ЦП1, ЦП2, ЦП6, ЦП 13 являются полночленными, вегетативно ориентированными, молодыми.
Возрастная структура ЦП, произрастающих на хорошо освещенных заброшенных делянках ботанического сада КБГУ (ЦПЗ) и совхоза «Декоративные культуры» (ЦП4) отличается значительным повышением доли особей ранних этапов онтогенеза - проростков (15,04-18,17%), ювенильных (12,76-14,65%) и имматурных (20,2420,55%) растений. Вероятно, в условиях пониженной межвидовой конкуренции (обработка почвы, прополка в предшествующие годы) высокая приживаемость подроста определяет наличие в ЦП волн возобновления, определяющей способность к захвату новых территорий и инвазионность возрастных спектров.
Характер возрастной структуры ЦП 17 и ЦП 18, реинтродуцированных нами в 2011 г. на территории Белореченского и Урванского лесничеств Кабардино-Балкарии, свидетельствует об успешной адаптации растений в условиях пониженного уровня антропогенной нагрузки под пологом густого букового леса. Семенное размножение высаженных генеративных растений в течение предшествующих четырех лет, определило наличие в 2016 г. в ценопопуляциях обособленных особей прегенеративного периода, среди которых особенно велика доля ювенильных (15,85-21,15%) и имматурных (21,35-22,40%) растений. Неполночленность спектров (отсутствие особей постгснсративного периода) и низкая доля виргинильных особей (16,45-18,76%) обусловлены молодостью ЦП17 и ЦП18. Число растений, пересаженных на данную территорию, таким образом, возросло с 50 и 65 до 274 и 315 особей соответственно, имеет место тенденция к расширению площади распространения.
Пятилетние наблюдения за изменениями возрастной структуры ЦП проводили в естественных местообитаниях в окр. с. Кашхатау и пос. Герпегеж (ЦП1 и ЦП2), подверженных высокому антропогенному давлению, а также в приближенных к естественным условиях заброшенных лесных участков республиканского ботанического сада «Декоративные культуры» (ЦПЗ) и ботанического сада КабардиноБалкарского государственного университета (ЦП4).
Исследования позволили установить наличие выраженных динамических процессов во всех изученных ЦП, характер которых значительно отличается в разных условиях. В ЦП1 и ЦП2, характеризующихся усилением антропогенной нагрузки в ряду лет, наблюдается выраженная тенденция к сокращению доли генеративных особей в возрастных спектрах, в то время как процент проростков и ювенильных растений остается стабильно низким (рис. 3).
Как сказано выше, большое значение здесь имеет сбор цветущих растений на букеты и выкапывание корневищ, а также медленное развитие растений при нарушении водного режима и аэрации переуплотненных почв. Стабильный, хотя и низкий, уровень семенного возобновления обеспечивается, вероятно, не только за счет семенного размножения, но и за счет прорастания семян почвенного банка.
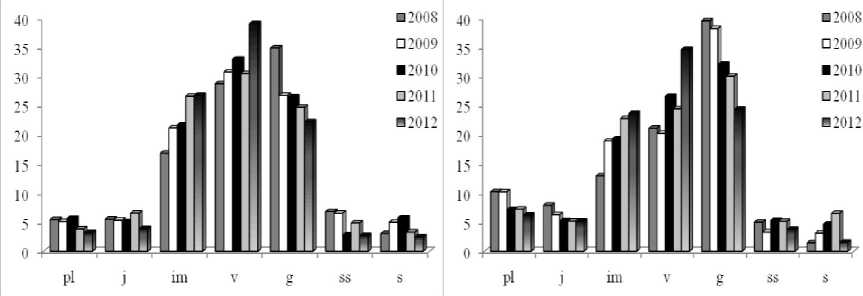
Рис. 3 . Динамика возрастной структуры ЦШ (слева) спектры и ЦП2 (справа) в ряду лет (2008-2012 гг.). По оси ох - возрастные группы pl-s, по оси оу - доля возрастной группы в спектре, %.
Динамика возрастной структуры ЦПЗ и ЦП4, произрастающих на заброшенных участках ботанических садов в условиях пониженной межвидовой конкуренции и среднего уровня антропогенной нагрузки, проявляется, в основном, в направленном увеличении в ряду лет доли особей прегенеративного периода семенного происхождения (рис. 4).
а 2008
□2009
□ 2008
□ 2009
■ 2010
□ 2011
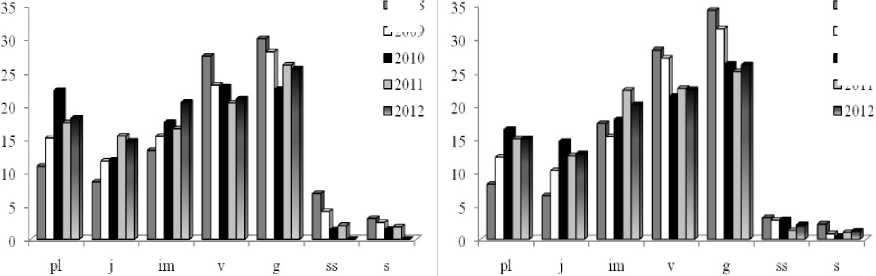
Рис. 4 . Динамика возрастной структуры ЦПЗ (слева) спектры и ЦП4 (справа) в ряду лет (2008-2012 гг.). По оси ох - возрастные группы pl-s, по оси оу - доля возрастной группы в спектре, %.
Подобные результаты свидетельствуют о протекании в ЦП волны возобновления, затрагивающей левую часть онтогенетических спектров. Вероятно, благодаря семенному размножению и высокой приживаемости проростков на ненарушенных плодородных почвах при низкой конкуренции со стороны других видов Н. caucasicus способен захватывать свободные территории.
Выводы. Таким образом, для Н. caucasicus в целом характерна высокая представленность в онтогенетической структуре ЦП виргинильной и генеративной возрастных групп, образующих локальные и абсолютные пики, что во многом обусловлено биологическими особенностями вида - способностью растений к юношеской и зрелой партикуляции, а также длительным пребыванием растений в генеративном периоде.
Соотношение возрастных групп в конкретных ЦП во многом зависит от характера и степени антропогенной нагрузки. Вырубка леса, вытаптывание, выпас скота в лесных массивах, сбор растений на букеты и в качестве лекарственного сырья приводят к снижению в спектрах доли генеративных и ювенильных растений, проростков, к быстрому отмиранию особей постгенеративного периода, выраженному сокращению физической и эффективной плотности, падению активности вегетативного размножения. Все это негативно сказывается на способности ЦП к самоподдержанию и их устойчивости.
На плодородных ненарушенных почвах при низком уровне межвидовой конкуренции, например, на заброшенных участках ботанических садов, где в предыдущие годы велась прополка, Н. caucasicus способен быстро осваивать свободные территории благодаря активному семенному возобновлению, о котором свидетельствует инвазионный характер возрастных спектров.
При реинтродукции в лесные фитоценозы с низким уровнем антропогенной нагрузки генеративные особи Н. caucasicus способны проходить полный цикл сезонного развития и образовывать полноценные семена, о чем свидетельствует наличие в ЦП молодых растений семенного происхождения.
Выражаем искреннюю благодарность к.б.н. Крапивиной Е.А., а также к.б.н. Чадаевой В.А. за помощь и консультации, оказанные при сборе и обработке полевого материала.
Об авторах:
КАРАМУРЗОВА Марьят Мухамедовна - учитель биологии МОУ СОШ №1 с.п. Атажукино Минобрнауки КБР, 360051, КабардиноБалкарская Республика, с.п. Атажукино, ул. Катанчиева, ПО, e-mail: karamurzo va8 7 @mail ,ru
ШХАГАПСОЕВ Сафарби Хасанбиевич - доктор биологических наук, профессор, заместитель председателя Комитета Парламента Кабардино-Балкарской Республики по аграрной политике, экологии, природопользованию и земельным отношениям, 360051, КабардиноБалкарская Республика, г. Нальчик, пр. им. В. И. Ленина, 55
Карамурзова М.М. Демографические показатели ценопопуляций Helleborus caucasicus X. Вг. На территории российской части Кавказа и Республики Абхазия / М.М. Карамурзова, С.Х. Шхагапсоев // Вести. ТвГУ. Сер. Биология и экология. 2018. № 1.С. 113-122.
Список литературы Демографические показатели ценопопуляций Helleborus caucasicus A. Br. на территории российской части Кавказа и Республики Абхазия
- Глотов Н.В. 1998. Об оценке параметров возрастной структуры популяций растений//Жизнь популяций в гетерогенной среде: материалы Всероссийского популяц. семинара. Йошкар-Ола: Периодика Марий Эл. С. 146-149.
- Гулиа В.О., Орловская Т.В. 2014. Морфолого-анатомическое исследование генеративных органов Helleborus caucasicus и Helleborus abchasicus//Современные проблемы науки и образования. № 6. С. 70-77.
- Гулиа В.О., Челомбитько В.А. 2003. Сравнительное анатомическое изучение Helleborus caucasicus A. Br. и Helleborus abchasicus A. Br. флоры Кавказа//Разработка, исследование и маркетинг новой фармацевтической продукции: сб. науч. трудов. Вып. 58. С. 27-29.
- Гулия В.О., Орловская Т.В. 2015а. Демографические характеристики ценопопуляций видов рода Helleborus L. в условиях Республики Абхазия//Научные ведомости. Серия Естественные науки. № 15 (212). Вып. 32. С. 5-12.
- Гулия В.О., Орловская Т.В. 2015б. Оценка виталитетного состава ценопопуляций Helleborus caucasicus A. Br. и Helleborus abchasicus A. Br. в условиях Абхазии//Известия Самарского научного центра Российской академии наук. Т.17. №4(4). С. 641-647.
- Гусева И.Н. 2015.Флора лесов Центрального Предкавказья и её анализ: автореферат дис.. кандидата географических наук. Ставрополь. 23 с.
- Животовский Л.А. 2001. Онтогенетические состояния, эффективная плотность и классификация популяций//Экология. №1. С. 3-7.
- Жукова Л.А. 1987. Динамика ценопопуляций луговых растений в естественных фитоценозах//Динамика ценопопуляций травянистых растений: сб. тр. Киев: Наукова думка. С. 9-19.
- Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола: РИИК «Ланар». 224 с.
- Загурная Ю.С., Алексеева А.И. 2010. Редкие виды сосудистых растений предгорной части Республики Адыгея//Экологические проблемы современности. Рациональное природопользование и сохранение биоразнообразия: мат. XVI международ. научно-практич. конф. Майкоп: Изд-во МГТУ. С. 132-136.
- Зернов А.С. 2006. Флора Северо-Западного Кавказа. М.: Товарищество научн. Изданий КМК. 664с.
- Олексин Ю.Р., Рыбалко А.Е. 2015. Введение в культуру in vitro редкого лекарственного растения Helleborus сaucasicus A. Brown//Биотехнология: состояние и перспективы развития: мат. VIII Московского международ. конгресса. М.: ЗАО «Экспо-биохим-технологии». С. 62-63.
- Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах//Труды БИН АН СССР. Серия Геоботаника. Вып. 6. С. 7-204 с.
- Работнов Т.А. 1992. Фитоценология: Уч. пос. -3-е изд., перераб. и доп. М.: Изд. МГУ. 352 с.
- Тайсумов М.А. 2012. Общий анализ флоры лекарственных растений Чеченской Республики//Вестник Академии наук Чеченской Республики. № 2(17). С. 83-90.
- Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов//Научные доклады ВШ. Биол. наук. № 2. С. 7-34.
- Хасуева Б.А., Астамирова М.А., Теймуров А.А. 2008. Третичные реликты в лесах Чечни и Ингушетии//Юг России: экология, развитие. № 2. С. 76-79.
- Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья. 990 с.
