Dependence of guaiacol peroxidase activity on ph in officinal plant leaves

Author: Zhivetyev M.A., Graskova I.A., Voinikov V.K.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 1 т.9, 2013.
Free access
For the first time, the guaiacol peroxidase activity have been studied by different pH in lamina of Achillea asiatica Serg., Veronica chamaedrys L., Taraxacum officinale Wigg., Alchemilla subcrenata Buser.
Peroxidase, officinal plants
Short address: https://sciup.org/14323718
IDR: 14323718
Text of the scientific article Dependence of guaiacol peroxidase activity on ph in officinal plant leaves
Пероксидаза – фермент, обладающий выраженным полиморфизмом. Наличие множества изоформ этого фермента позволяет ему работать в разных условиях и выполнять разную функциональную нагрузку (Граскова и др., 2002). Изменения активности, изоферментного спектра и термостабильности пероксидазы отмечают при различных биологических, физических и химических воздействиях на растения (Андреева и др., 1979; Карпец и др., 2009).
Активность пероксидазы зависит от pH, температуры, концентраций субстрата и самого фермента (Kareska, 2009). Известно, что на перикарпе китайской сливы максимальная активность пероксидазы наблюдается при рН 6,5 и 70 ºС, а полифенолоксидазы при рН 6,5–6,75 и 20 ºС (Mizobutsi et al., 2010). Susan Kareska (2009) показано, что максимум активности пероксидазы приходится на рН 7, а минимум на 4. Zolfaghari с соавторами (2010) на буке (Fagus orientalis) показали увеличение активности пероксидазы от июля к сентябрю при отсутствии роста активности каталазы. В то же время считается общепринятой взаимосвязь резистентности растительных тканей к различным тепловым воздействиям и активностью ферментов, обладающих пероксидазной активностью (Müftlügil, 1985; Thongsook, Barrett, 2005).
В этой связи, целью наших исследований было изучить активность гваякол пероксидаз в листьях вероники дубравной, тысячелистника азиатского, одуванчика лекарственного и манжетки городковатой, произрастающих на побережье озера Байкал, в разные сроки вегетационного периода в зависимости от рН в реакционной смеси.
МАТЕРИАЛЫ И МЕТОДЫ
Материал для исследования. Исследовались листья тысячелистника азиатского Achillea asiatica Serg., вероники дубравной Veronica chamaedrys L., одуванчика лекарственного Taraxacum officinale Wigg., манжетки городковатой Alchemilla subcrenata Buser, собранных в течение вегетационного периода 2008 года на территории стационара «Речка Выдринная» в июле, августе и сентябре.
Выделение гваякол пероксидаз. Навеску (1 г) ткани помещали в 10 мл холодного цитратно-фосфатного буфера и растирали в фарфоровой ступке при 4 °С (Паду, 1995). Полученный гомогенат центрифугировали при при 3 тыс. об./мин. в течение 15 мин. Супернатант использовали для определения активности фермента.
Определение пероксидазной активности. Активность растворимых пероксидаз в листьях растений определяли по изменению оптической плотности (длина волны 580 нм) в реакционной смеси следующего состава: 0,5 мл 0,1 М цитратно-фосфатного буфера (рН от 4,0 до 7,0 с шагом 0,2), 0,5 мл 0,3 % перекиси водорода («Реахим», Россия), 0,5 мл 0,05 % гваякол (Sigma, США) и 0,5 мл пробы (1г пробы растирали в 10 мл Цитратно-фосфатного буфера рН 5,5).
Активность пероксидазы определяли при 25 °С сразу после выделения ферментов из образцов. Активность фермента рассчитывали по методу Бояркина (Бояркин, 1951) и выражали в условных единицах на мг сырого веса тканей по формуле: А = ∑ (α β γ)/d t, где ∑ - экстинция (0,125), α – отношение количества буфера, взятого для приготовления вытяжки в мл к весу сырой ткани, β – степень дополнительного разведения вытяжки в реакционной смеси, γ – степень постоянного разведения вытяжки в реакционной смеси, d – толщина поглощающего слоя кюветы, t – время реакции.
РЕЗУЛЬТАТЫ
Впервые была изучена активность гваякол пероксидаз при разных значениях рН в листовых пластинках тысячелистника азиатского Achillea asiatica Serg., вероники дубравной Veronica chamaedrys L., одуванчика лекарственного Taraxacum officinale Wigg., манжетки городковатой Alchemilla subcrenata Buser (рис. 1).
Для вероники дубравной (рис. 1-А) и манжетки городковатой (рис 1-Г) наблюдали уменьшение активности пероксидаз с июля по сентябрь, в то время как для тысячелистника азиатского (рис. 1-Б), наоборот, увеличение. Для одуванчика лекарственного (рис. 1-В) минимум нейтральных пероксидаз приходился на сентябрь, слабокислых на июль, а кислых на август.
Соответственно, в июле в листьях одуванчика преобладали нейтральные гваякол пероксидазы с оптимумом рН 6,6-6,2 и кислые с оптимумом рН 4,8-4,6. В августе у одуванчика лекарственного повышалась активность слабокислых пероксидаз с оптимумом рН 5,8. К сентябрю максимальная активность гваякол пероксидаз у этого растения сохранялась при рН 5,8, но этот пик активности значительно ниже, что характеризует понижение активности фермента и истощение адаптационных ресурсов осенью у этого вида.
Активность пероксидазы, Активность пероксидазы, Активность пероксидазы, Активность пероксидазы, усл, ед./г сырой массы усл. ед./г сырой массы усл ед/г сырой массы усл. ед./г сырой массы
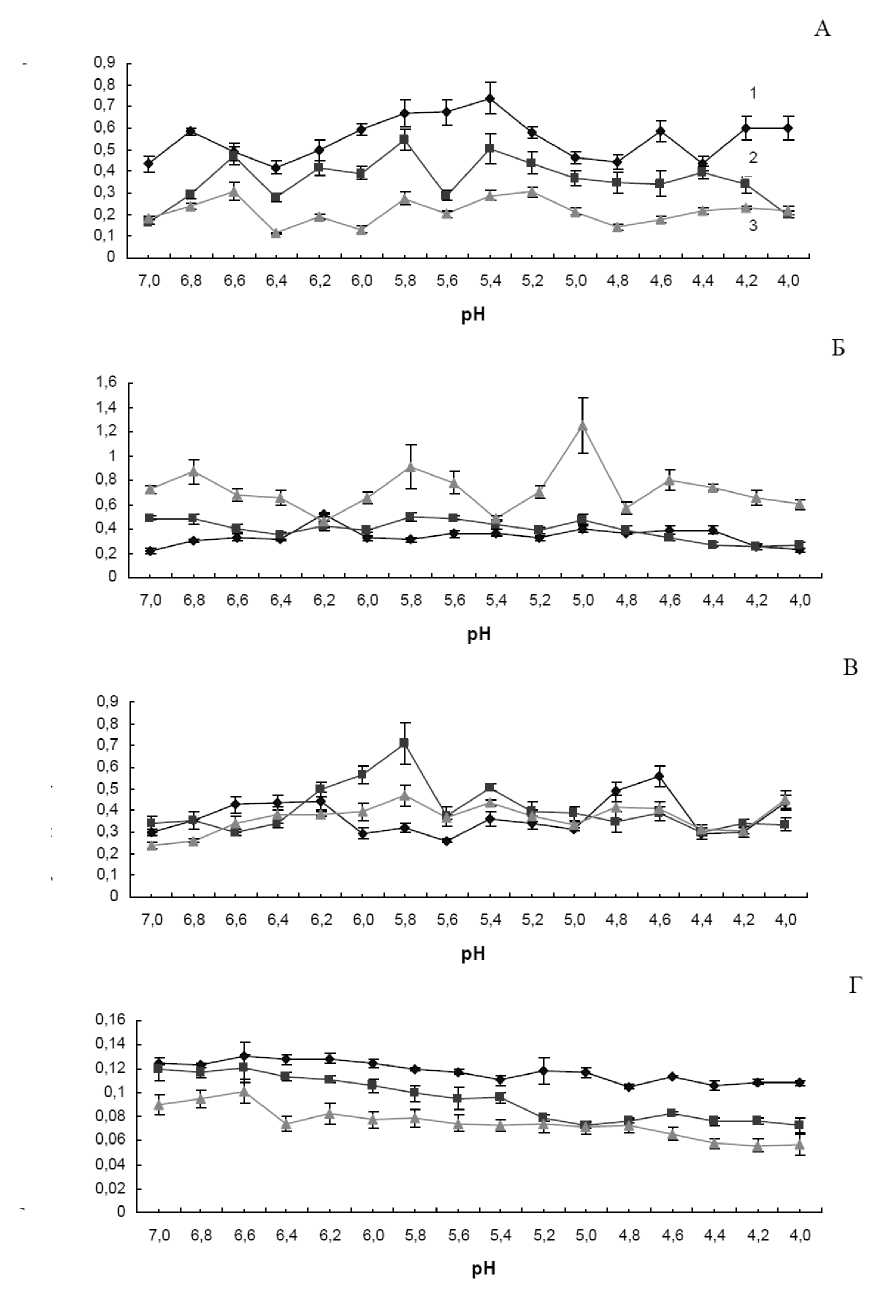
Рисунок 1. Активность общих кислых, слабокислых и нейтральных гваякол пероксидаз в листьях вероники дубравной (А), тысячелистника азиатского (Б), одуванчика лекарственного (В) и манжетки городковатой (Г) на побережье озера Байкал, стационар «речка Выдриная», 2008 г. 1 – июль, 2 – август, 3 – сентябрь
Таким образом, выраженное увеличение активности всех изученных пероксидаз с наступлением неблагоприятных условий произрастания в сентябре наблюдали только у тысячелистника азиатского, что может свидетельствовать о его холодоустойчивости и высоком адаптационном потенциале по отношению к гипотермии. Причем, если в июле у тысячелистника оптимум рН приходился на 6.2, с наступлением августа этот пик перекрывали пероксидазы с активностью при рН 7.0-6.8, 5.8 и 5.0, что может косвенно свидетельствовать о начавшейся экспрессии соответствующих форм фермента. К октябрю все три указанных пика еще сильнее возрастают и дополняются еще одним – 4,6. Минимум активности фермента в сентябре приходится на рН 6.2.
Интересна активность разных групп пероксидаз у манжетки. У нее во все сроки исследований наблюдается общая закономерность: активность кислых пероксидаз была ниже нейтральных и слабокислых гваякол пероксидаз.
Для вероники на фоне общего уменьшения активности пероксидаз с наступлением осени выделяются стабильные пики при рН 6.8-6.6, 5.8, 5.4-5.2, 4.6-4.2, что может свидетельствовать о стабильности изоферментного состава пероксидаз у этого вида в течение всего исследуемого периода вегетации. Для вероники наблюдается совпадающее с литературными данными уменьшение активности фермента в ряду рН от 7 к 4 (Mizobutsi et al., 2010; Kareska, 2009).
ЗАКЛЮЧЕНИЕ
В листьях тысячелистника азиатского, вероники дубравной, одуванчика лекарственного и манжетки городковатой впервые была изучена активность пероксидаз в зависимомти от рН.
Для вероники дубравной и манжетки городковатой наблюдали уменьшение активности фермента с июля по сентябрь, а для тысячелистника азиатского, наоборот, увеличение. Для одуванчика лекарственного минимум нейтральных пероксидаз приходился на сентябрь, слабокислых на июль, а кислых на август. Для вероники наблюдали уменьшение активности фермента в ряду рН от 7 к 4.
References Dependence of guaiacol peroxidase activity on ph in officinal plant leaves
- Андреева В.А., Воронова В.А., Угарова Н.Н. (1979) Активность, изоферментный спектр, термостабильность и молекулярный вес пероксидазы, выделенной из здоровых и зараженных вирусами растений табака. Биохимия. 44(3), 394-399.
- Бояркин А.Н. (1951) Быстрый метод определения активности пероксидазы. Биохимия. 16. 352.
- Граскова И.А., Боровский Г.Б., Владимирова С.В., Романенко А.С., Войников В.К. (2002) Изоферментные спектры пероксидазы картофеля при патогенезе кольцевой гнили. Докл. РАН. 384(6), 844-847.
- Карпец Ю.В., Ястреб Т.О., Обозный А.И., Колупаев Ю.Е. (2009) Активность и термостабильность антиоксидантных ферментов корней проростков пшеницы после воздействия экзогенного пероксида водорода. Вестник Харьковского национального аграрного университета. Серия Биология. 17(2). 62-70.
- Паду Э.Х. (1995) Свойства пероксидазы и фенилаланин-аммиак-лиазы при образовании и лигнификации клеточных стенок стебля пшеницы. Физиология растений. 42. 408-415.
- Kareska S. (2009) Factors affecting hydrogen peroxidase activity. ESSAI. 7(27). 82-85.
- Mizobutsi G.P., Finger F.L., Ribeiro R.A., Puschmann R., de Melo Neves L.L., da Mota W.F. (2010) Effect of pH and temperature on peroxidase and polyphenoloxidase activities of litchi pericarp. Sci. Agric. (Piracicaba, Braz.). 67(2). 213-217.
- Müftügil N. (1985) The peroxidase enzyme activity of some vegetables and its resistance to heat. J. Sci. Food Agric. 36. 877-880.
- Thongsook T., Barrett D.M. (2005) Heat Inactivation and Reactivation of Broccoli Peroxidase. J. Agric. Food Chem. 53. 3215-3222.
- Zolfaghari R., Hosseini S.M., Korori S.A.A. (2010) Relationship between peroxidase and catalase with metabolism and environmental factors in Beech (Fagus orientalis Lipsky) in three different elevations. International J. of Environmental Sciences. 1(2). 243-252.
