Детализированная модель динамики структурированной популяции

Автор: Пойколайнен Вадим Вальтерович, Сиговцев Геннадий Сергеевич
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Физико-математические науки
Статья в выпуске: 6 (119), 2011 года.
Бесплатный доступ
Матричная модель структурированной популяции, проекционная матрица, задача оборота стада
Короткий адрес: https://sciup.org/14749977
IDR: 14749977
Текст статьи Детализированная модель динамики структурированной популяции
В агропромышленном секторе экономики одной из важных задач является планирование оборота стада сельскохозяйственных животных [10], [11]. Оборотом стада называется происходящая во времени совокупность процессов, включающая переход животных из одной стадии (категории) в другую, удаление животных разных стадий из стада и появление потомства.
В качестве основы для построения модели стада может быть использовано понятие структурированной популяции. В математической биологии под этим понимают множество проживающих в одной локации организмов одного вида, дискретно разбитых на некоторые классы по какому-либо признаку. Группы могут определяться полом, начальным и конечным возрастом и другими существенными для целей моделирования признаками. Поэтому в описании структурированных популяций используют понятие последовательных стадий жизни организма.
Состояние популяции, разделенной на некоторое число стадий, рассматривается в равноотстоящие дискретные моменты времени и описы-ваетсявектором-столбцом X ( t ) = ( x t д, xt 2 2 ,..., xt n ) T , где x t i - численность организмов, отнесенньтх к стадии i , n - количество стадий. Динамика популяции задается соотношением
X(t +1) = L • X ( t ), t = 0, 1, .„, (1) определяющим состояние популяции в момент t + 1 по состоянию в момент t посредством матрицы L t , которую называют проекционной. В общем случае элементы проекционной матрицы могут зависеть от времени.
РАЗВИТИЕ МОДЕЛЕЙ СТРУКТУРИРОВАННЫХ ПОПУЛЯЦИЙ
Простейшим вариантом модели (1) является модель Лесли [8], в которой стадиями являются © Пойколайнен В. В., Сиговцев Г. С., 2011
возрастные группы, а проекционная матрица имеет вид:
b l Ь 2 ... b n - 1 b n
|
S 1 0 ... 0 0 |
||
|
L = |
0 s 2 ... 0 0 |
, (2) |
0 0 ... S n - 1 0
0 < b i - коэффициент рождаемости для стадии i , 0 < s i < 1 — коэффициент дожития (перехода из стадии i в стадию i + 1).
Если для квадратной матрицы A порядка n и ориентированного графа G с n вершинами, пронумерованными числами от 1 до n , существует взаимно однозначное соответствие между ненулевыми элементами a . и дугами ( i , j ), то такой граф называется ассоци, ированным с матрицей и обозначается G(A). Граф, ассоциированный с проекционной матрицей структурированной популяции, называют графом жизненного цикла (ГЖЦ) организмов данной популяции.
Для модели Лесли ГЖЦ изображен на рис. 1.
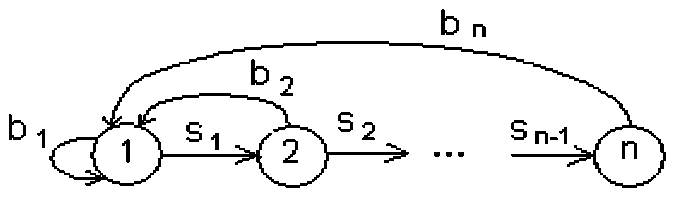
Рис. 1. ГЖЦ модели Лесли
В модели Лесли на каждом шаге происходит полное обновление состава каждой возрастной группы, что является достаточно жестким ограничением для практического использования этой модели. Более общей является модель Лефковича [8], в которой часть особей каждой стадии остается в той же стадии при переходе к следующему моменту времени. Проекционная матрица модели Лефковича имеет вид:
|
" b 1 + r |
b 2 . |
•• bn - 1 |
b n " |
||
|
s 1 |
Г 2 . |
.. 0 |
0 |
||
|
L = |
0 |
s 2 . |
.. 0 |
0 |
, 3 |
|
. 0 |
0. |
.. sn - 1 |
r n _ |
где г. - коэффициент задержки, 0 < s + r < 1.
ГЖЦ для модели Лефковича изображен на рис. 2.
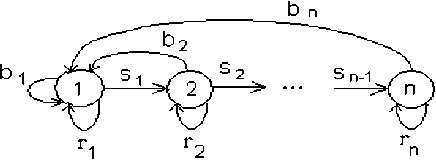
Рис. 2. ГЖЦ модели Лефковича
ГЖЦ на рис. 1 и 2 показаны для случая, когда все параметры проекционных матриц b i , r > 0. Если какой-то из этих параметров равен нулю, то соответствующая дуга ГЖЦ отсутствует.
С математической точки зрения модели Лесли и Лефковича достаточно хорошо изучены [6], [8]. Основные их свойства связаны с теоремой Перрона - Фробениуса для матриц с неотрицательными элементами [2; 130]. В соответствии с этой теоремой спектральный радиус неотрицательной неразложимой матрицы равен максимальному положительному собственному числу. Это число имеет кратность 1, ему соответствует положительный собственный вектор, и оно возрастает при увеличении любого элемента матрицы.
Неразложимость матрицы эквивалентна тому, что ассоциированный с ней орграф сильно связен (для любой пары вершин существует ориентированный путь из одной вершины в другую). В частности, проекционные матрицы моделей Лесли и Лефковича неразложимы, если b n > 0 (последняя стадия популяции является репродуктивной).
Таким образом, спектральный радиус проекционной матрицы р ( L ) = X max - положительное собственное число матрицы L кратности 1.
Асимптотика вектора X(t), определяемого соотношением (1), при t ^ го зависит от величины спектрального радиуса р ( L ) следующим образом:
Р (L ) > 1 x , , t ^ « ,
р ( L ) < 1 x. t ^ 0 , (4)
Р(L) = 1 xi,t ^ x,*, для каждой стадии г = 1, ..., n.
Последний случай означает устойчивость по -пуляции: ее численность в каждой стадии стре- мится к конечному пределу, не зависящему от начальной численности популяции. При этом из уравнений (1) для матрицы L вида (3) следует, что предельные численности стадий в модели Лефковича связаны следующими соотношениями:
x*, + 1 = 1 si • x*, , i = 1, ..., n - 1,
1 - r + 1
*
x 1
1 - b 1 - r
n
• I bx ‘.
= 2
Следствием соотношений (5) является условие:
b 1 + r 1 + I b • q , = i, где q , = П s j- . (6)
I = 2 /=1 1 - rj + 1
Так как вектор предельных численностей X является собственным вектором проекционной матрицы, соответствующим собственному числу X max = 1, то условие (6) - это условие стабильности популяции. Условие (6) соответствует приведенному в [3] результату об индикаторе потенциального роста популяции в модели Лефковича.
Собственный вектор X при условии, что n
I x *• = 1 , называется относительной стадийной г = 1
структурой популяции [6].
Из соотношений (5) следует, что вектор относительной структуры популяции в модели Лефковича имеет вид:
T n , n ,.-., n
1+ I q , 1+ I q, 1+ I q ,
\ i = 2 i = 2 i = 2 /
Условие bn > 0 не является принципиальным ограничением для применения описанных выше моделей [8; 58]. Наличие в модели популяции пострепродуктивных стадий не влияет на динамику стадий популяции с номерами от 1 до n (последняя репродуктивная стадия), которая по-прежнему определяется матрицей (3). Численность пострепродуктивных стадий опре- деляется соотношением:
x„ + . , t + 1 = s n + , - 1 • xn + , - 1, t + r + , • xn + i,t , i = 1, -, m , (7)
где m - количество таких стадий.
ГЖЦ модели с пострепродуктивными стадиями показано на рис. 3.
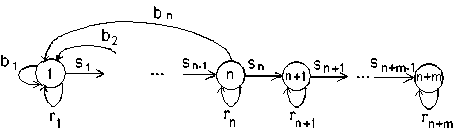
Рис. 3. ГЖЦ модели с пострепродуктивными стадиями
Данный граф не является сильно связным (отсутствуют пути из вершин n + 1, …, n + m в вершины 1, …, n ); соответственно, проекционная матрица L такой модели разложима. Она имеет вид:
L 0
SM
где L – матрица (3), M – m × n – матрица переходов для пострепродуктивных стадий
r n + 1
|
L = |
" b 1 + Г 1 s 21 s 31 |
b 2 r 2 s 3 2 |
- bn - 1 ... 0 ... 0 |
b 1 n 0 0 |
(10) |
|
[ s n 1 |
s n 2 |
... nnn - 1 n |
r n . |
||
|
при условии, что ..., n – 1. |
0 < r j |
+ Z s-^ 1 i = j + 1 |
для всех j = 1, |
||
M =
s n + 1
r n + 2
s , r n + m - 1 n + m
S – m × n –
матрица с единственным ненулевым элементом sn в правом верхнем углу, связывающим стадии n и n + 1.
Вектор состояния популяции X ( t ) = ( xt ,1 , xt ,2 , ..., xt , n ) T может быть разбит на два вектора X I ( t ) = = ( xt ,1 , xt ,2 , ..., xt , n ) и X ( t ) = ( xt , n +1 , xt , n +2 , ..., xt , n+m ) . Изменение численности популяции описывает-
ся системой уравнений:
[ X I ( t + 1) = L ■ X I ( t )
.
X " ( t + 1) = 5 ■ X I ( t ) + M ■ X ” ( t )
Асимптотика вектора X I ( t ), определяемого соотношением (9), при t → ∞ по-прежнему зависит от величины спектрального радиуса ρ ( L ) так, как это указано в (4).
Характер асимптотики вектора X II ( t ) однозначно определяется поведением вектора X I ( t ) при t ^ да. Если lim xti = x * , i = 1, ..., n, то по ин- ^ro ,
дукции доказывается, что существуют пределы xt,n + i для i = 1, ..., m при t → ∞.
Из (7) также следует, что если lim xti =» , i = 1,
^да ,
..., n, то lim xt n + i = да для всех j = 1, ..., m . t ^to ,
Вектор относительной структуры популяции имеет такой же вид, как и выше (с распространением суммирования по i от 2 до n + m ).
В работах Д. О. Логофета и других [5], [6], [7] рассмотрены некоторые обобщения моделей Лесли и Лефковича, связанные с необходимостью в некоторых случаях классифицировать особей популяции по нескольким признакам. Например, в [14] модель популяции северных оленей учитывает их возраст и статус («сильный», «средний», «слабый»).
Такого типа модель может быть записана в форме матричного уравнения (1), если матрицу двойной структуры популяции (строкам матрицы соответствует возраст, а столбцам – статус особи) представить в виде вектора, образованного конкатенацией строк [5]. Проекционная матрица такой модели, называемая обобщенной матрицей Лефковича или матрицей Логофета [4], в общем случае имеет вид:
Для матрицы (10) также существует выражаемый через элементы матрицы индикатор потенциального роста [6], обобщающий условие существования стабильной популяции (6).
В ряде работ указывается, что матричные модели не всегда удается должным образом применить для решения некоторых практических задач ввиду недостаточно явно представленной динамики популяции [3]. Например, в работах [13], [15] предлагается детализировать матричный подход к расчету динамики структурированной популяции, разложив его на «скрытые» субпроцессы. Из состояния популяции Xt в момент t рассчитывается численность выживших за шаг времени XS ( t ), затем взросление XA ( t ) и получение потомства XB ( t ), таким образом получается численность популяции в момент t + 1: X ( t ) ^ Xs ( t ) ^ X A ( t ) ^ X B ( t ) ^ X ( t + 1) . Такая детализация динамики в некоторой степени упрощает определение модельных параметров.
Дискретные модели широко применяются для описания эксплуатируемых популяций, когда производится регулируемое изъятие некоторой части особей, принадлежащих определенным стадиям, а также для постановки и решения оптимизационных задач [9], [11].
ДЕТАЛИЗИРОВАННАЯ МОДЕЛЬ СТРУКТУРИРОВАННОЙ ПОПУЛЯЦИИ
Популяции типа стада сельскохозяйственных животных относят к эксплуатируемым популяциям, хотя для них характерны не только процессы изъятия части особей, но и добавление извне новых особей, а также другие целенаправленные воздействия управляющего субъекта, влияющие на состав и численность популяции. В этом случае более адекватным будет термин «управляемая популяция».
Использование дискретной модели динамики управляемой популяции для задачи планирования оборота стада сельскохозяйственных животных и рассмотрения на этой основе оптимизационных задач планирования работы агрохо-зяйственного предприятия требует более детального анализа динамики численности. В задачах планирования, как правило, расчеты производятся на определенный временной промежуток, разделяемый на несколько равных периодов длиной T.
Особи стада могут находиться в одной из n стадий, принадлежность к которым определяется возрастом особи тк , измеряемым в некоторых единицах дискретного модельного времени t = 0, 1 ..., m . Каждой стадии i соответствует свой возрастной промежуток [ т . 1 , т. 2 ]; промежутки разных стадий не пересекаются. Задан предельный возраст T m = m , при достижении которого все особи из популяции изымаются.
Будем считать величину периода T равной некоторому количеству единиц модельного времени. Обозначим через T = т. 2 - т. 1 продолжительность стадии i . В задачах планирования оборота стада обычно T > minT. [10], [11], поэтому за один период особь может пройти несколько стадий. Такая же возможность существует и для особи, родившейся в течение этого периода.
Основа детализации модели - в рассмотрении изменений численности различных стадий популяции за единицу модельного времени за счет изменения возраста особей с учетом изъятия части особей и появления особей-потомков.
Пусть заданы функция h ( т, t ) - доля изымаемых в момент времени t особей возраста T в общей численности особей этого возраста, и функция g ( т, t) - коэффициент репродуктивности особей возраста на момент времени t . Если обозначить Y ( t ) = ( yt 1 , yt 2,..., ytm ) T вектор, компонентами которого явлЯют , ся чиСленности особей каждого возраста в момент времени t , то для него будет справедливо соотношение вида (1) Y ( t + 1) = -Л t • Y ( t ) , где Л t имеет вид матрицы Лесли:
Л t -
е
e q
в т
0 0 ... <- 1
вк = g(тк, t), ^к = 1 — h(тк , t), q - номер начального возраста первой репродуктивной популяции.
Для вектора X ( t ) = ( x, ,1 , xt ,2 , ..., x, , n ) T численностей стадий популяции соотношение (1) будет иметь место с проекционной матрицей L , имеющей вид матрицы Лефковича
|
r t . |
.. b p . |
.. Ь П |
|
|
s 1 . |
.. 0 . |
.. 0 |
|
|
L - |
0. |
г1 |
.. 0 |
|
.. p . |
0 0 ... s n - 1 r
Элементы матрицы L t определяются через элементы матрицы Л t , p - номер первой репродуктивной популяции. y
Обозначим через Л - m t , к долю особей по-
X у, ,j j=1
пуляции возраста тк во всей численности попу- ляции, fkt =
fkt
X f t
- долю особей популяции воз- j е Ki раста тк, принадлежащего i-й стадии в численности этой стадии популяции (Ki - множество индексов, принадлежащих диапазону возрастов особей стадии i). Тогда для элементов матрицы Lt получаем :
si= <_2■ f_2, bi-Xe,-f,, ri- Xjf je к. je к.\{ i _2}
Структурированная численность популяции через плановый период времени T будет опреде -ляться для возрастной модели соотношением:
Y(T ) -Л( T ) ■ Y (0), (11)
Л(T) -Л1 ■Л.2 ■...■Лi, где l - количество единиц мо- дельного времени в периоде.
В том частном случае, когда проекционные матрицы детализированной модели не зависят от времени ( Л , -Л ) , имеем л ( t ) -Л 1 и для спектрального радиуса р ( Л T ) ) = [ р ( Л )] 1 . В этом случае каждый из вариантов условий (4) для матриц детализированной и недетализированной моделей выполняется одновременно, и для выяснения устойчивости популяции достаточно проверки условия (6), которое для матрицы Л имеет вид:
m i - 1
X в . чп ^ -) = ! • i - q j - 1
При моделировании популяции типа стада крупного рогатого скота более реалистичны нестационарные модели, когда зависимость параметров детализированной возрастной (а следовательно, и стадийной) модели от времени носит сезонно-циклический характер. Но если l кратно длине цикла изменения параметров, то матрица Л ( T ) от времени не зависит. При этом возможно выполнение условия устойчивости популяции р (Л ( T ) ) - 1 в ситуации, когда спектральные радиусы л , в разные моменты модельного времени принимают значения как больше, так и меньше единицы.
Структура проекционной матрицы планового периода T Л ( T ) при T < q показана на рис. 4.
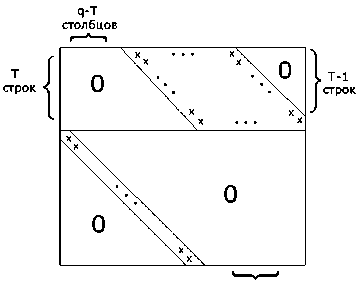
Т-1 столбцов
Рис. 4. Структура матрицы Л ( T )
Элементы, отмеченные значком х, выражаются через значения величин в, о матриц-сомножителей; остальные элементы равны нулю.
Элементы поддиагонали с номером T имеют вид:
j + T - 1
a, j = 1 l " k - j + 1 , j = 1 -’ m - T , i = T + j , (12)
r = j
а элементы параллелограмма в верхней части
матрицы имеют вид:
a
' i, j
g T - i + 1 P t - i+j
' T - i
П<- 1 , к = 1
i - 1
П
к = 1
к - i + T + 1
" к
основана на идее степенного метода нахождения наибольшего по модулю собственного числа матрицы [12].
Если квадратная матрица A порядка n - матрица простой структуры (то есть имеет n линейно независимых собственных векторов) и zk = Ak • z0, к = 1, 2 ..., то для всех значений индекса i = 1,..., n limlz-!!)^ = х, (15) к^” (zk )i
i = 1, ..., T, j = q - T + i,..., m - T + i . (13)
Если в (13) в одном из произведений верхняя граница индекса равна нулю, то произведение считается равным единице.
Для стадийной модели проекционная матрица планового периода L T ) может быть получена путем агрегации элементов матрицы Л T ) , относящихся к одной стадии. Для этого на матрицу Л T ) накладывается прямоугольная сетка с ячейками K , i , j = 1,..., n . K принадлежат элементы матрицы Л T ) a u v , где u g K i . Ячейке K i ^ соответствует элемент d i j матрицы L ( T ) .
Так как xT , = Z yT , l , из (11) получаем:
l G K i
где X - наибольшее по модулю простое собственное число матрицы A . (15) имеет место для произвольного вектора z 0 , имеющего ненулевой коэффициент C при соответствующем X собственном векторе x в разложении по базису из собственных векторов. При этом
m
m
/
X T , i = Z | Z a , и У 0, u l = Z Z a , u l У 0,u
l G K i V u = 1
u = 1 V l G K i
n
/
/
= Z Z Z a i,u
j = 1
V u G K j V l G K i
^ ^
У 0, u
7 7
zk ^ ХкСх при k ^ ^ . (16)
В [1; 242] показано, что равенство (15) остается в силе и для матрицы, не обладающей простой структурой, при условии, что X - единственное максимальное по модулю собственное значение единичной кратности. (Матрица, имеющая единственное собственное число с максимальным модулем, называется примитивной.)
Для проекционных матриц рассматриваемых моделей справедлива теорема Перрона -Фробениуса, по которой кратность максимального по модулю собственного числа равна единице. Как показано в [8; 64], матрицы Л t и Lt будут примитивными, если какие-то два соседних элемента их первых строк отличны от нуля. Для матрицы Lt , кроме того, для примитивности достаточно отличия от нуля хотя бы одного элемента r i [2; 132]. Примитивность матриц Л ( T ) и L ( T ) следует из того, что они являются произве-
Соотношение X(T ) = L ( T ) выполняется, если
Г ^
Z d-,j x o , j = Z Z a >.u y o , u l , где a i,u = Z a iu .
j=1 j=1 uGK, ) l GK, ji
Отсюда получаем: d i ., j =
Za v l,uy o,u uGKj
Z y ou u G K j
Разделив числитель и знаменатель на начальную численность популяции, получаем формулу для вычисления элементов матрицы L(T ) :
d i, = Z a , u f u . (14)
u G K j
В общем случае получение явных выражений для элементов матриц Л(T) и L(T), необходимых для проверки условий типа (6) или вычисления индикатора потенциального роста [6], весьма трудоемко. Эти проверки можно заменить вычислительной процедурой, результаты которой могут быть использованы как «эмпирический» индикатор потенциального роста. Процедура дениями примитивных матриц.
Таким образом, для получаемых с помощью проекционных матриц итерационных последовательностей векторов будут справедливы соот-( Zh+1)- к ношения (15) и (16). Обозначим------через w^.
( z- ) i i
В соответствии с (15), если X > 1, то для любой координаты i начиная с некоторой итерации wk > 1, и наоборот, неравенство wk > 1 свидетельствует, что X > 1. Аналогично, если начиная с некоторой итерации wk < 1, то X < 1. При этом из (16) следует, что |( zk ) i | ^» или |( zk ) i | ^ 0
соответственно.
Пусть теперь X = 1. В этом случае wk ^ 1 при к ^ го . Характер стремления wk к 1 может быть как монотонным, так и немонотонным.
О степени близости отношения wk к величине X на практике можно судить по совпадению (с некоторой точностью) значений этого отношения для разных значений индекса i [12; 331].
Таким образом, величина отношения wik , полученная после некоторого количества итераций, может играть роль численного индикатора потенциального роста. Необходимое количество итераций определяется в каждом конкретном случае потребностью различить, какой именно из трех возможных вариантов для величины λ (больше, меньше, равно 1) имеет место.
В качестве примера рассмотрим упрощенный вариант детализированной модели, соответствующей модели оборота стада крупного рогатого скота, предложенной А. А. Трофимовым [11].
Исходные данные: предельный возраст особей m = 12, функции h , g зависят только от возраста особей и имеют вид: h ( τ ) = {0,2, 0,05, 0,05, 0,05, 0,03, 0,03, 0,03, 0,03, 0,04, 0,04, 1}, g( τ ) = {0, 0, 0, 0, 0, 0, 0, 0,3, 0,4, 0,6, 0,4, 0,2}. Количество стадий n = 3, возрастные диапазоны стадий: K 1 = {1, 2, 3}, K 2 = {4, 5, 6, 7}, K 3 = {8, 9, 10, 111, 12}. Возрастная структура популяции f ( τ ) = {0,15, 0,12, 0,1, 0,08, 0,08, 0,08, 0,06, 0,1, 0,08, 0,05, 0,05, 0,05}, T = 6.
В рассматриваемом примере матрица Лt детализированной модели не зависит от времени, Л(т) = Л6. Вид этих матриц не приводится ввиду их громоздкости. Элементы проекционных матриц стадийной модели вычисляются по форму- лам (11), (14), и эти матрицы имеют вид соответственно:
|
0,63 |
0 |
0,37 |
0,07 |
0,86 |
0,15 |
|
|
0,26 |
0,78 |
0 |
, L т ) = |
0,26 |
0,15 |
0,69 |
|
0 |
0,19 |
0,82 |
0,47 |
0,64 |
0 |
Максимальные по модулю собственные числа: X (Л) = 1,010, X ( Л T ) ) = 1,064, X ( L ) = 1,014, X (Л( T ) ) = 1,097. Применение степенного метода дает следующие результаты: первые три знача- ( z )
щие цифры отношения p+1 i и максимального (zp)i по модулю собственного числа совпадают для Л(T) начиная с пятнадцатой итерации, а для L(T) – начиная со второй. (Начальные вектора z0 имели единичные компоненты.)
Предложенные в данной статье матричные модели структурированных популяций применительно к задаче оборота стада сельскохозяйственных животных позволяют анализировать последствия управляющих воздействий на популяцию. Воздействия в виде изъятия части особей учитываются через изменение функции изъятия h ( τ, t ), а добавление особей в стадо учитывается через изменение функции f ( τ, k ), описывающей возрастную структуру популяции.
Список литературы Детализированная модель динамики структурированной популяции
- Березин И. С., Жидков Н. П. Методы вычислений. Т. 2. М.: Гос. изд-во физ.-мат. лит., 1960. 620 с.
- Воеводин В. В., Кузнецов Ю. А. Матрицы и вычисления. М.: Наука, 1984. 320 с.
- Динамическая теория биологических популяций/Под ред. Р. А. Полуэктова. М.: Наука, 1974. 455 с.
- Клочкова И. Н. Обобщение теоремы о репродуктивном потенциале для матриц Логофета//Вестник Московского университета. Сер. 1. Математика, механика. 2004. № 3. С. 45-48.
- Логофет Д. О. Три источника и три составные части формализма популяции с дискретной стадийной и возрастной структурами//Математическое моделирование. 2002. Т. 14. № 12. С. 11-22.
- Логофет Д. О., Белова И. Н. Неотрицательные матрицы как инструмент моделирования динамики популяций: классические модели и современные обобщения//Фундаментальная и прикладная математика. 2007. Т. 13. № 4. С. 145-164.
- Логофет Д. О., Клочкова И. Н. Математика модели Лефковича: репродуктивный потенциал и асимптотические циклы//Математическое моделирование. 2002. Т. 14. № 10. С. 116-126.
- Свирежев Ю. М., Логофет Д. О. Устойчивость биологических сообществ. М.: Наука, 1978. 352 с.
- Скалецкая Е. И., Фрисман Е. Я., Шапиро А. П. Дискретные модели динамики численности популяций и оптимизация промысла. М.: Наука, 1979. 165 с.
- Терновых К. С. Планирование на предприятии АПК. М.: Колос, 2006. 79 с.
- Трофимов А. А., Чугин И. В. Моделирование оборота стада крупного рогатого скота и оптимальное планирование производства в агрохозяйстве//Проблемы информационной экономики. Вып. VI. Моделирование инновационных процессов и экономической динамики: Сб. науч. тр./Под ред. Р. М. Нижегородцева. М.: Ленанд, 2006. С. 212-225 [Электронный ресурс]. Режим доступа: http://www.econ.asu.ru/lib/sborn/model2006/pdf/14.pdf
- Фаддеев Д. К., Фаддеева В. Н. Вычислительные методы линейной алгебры. М.: Гос. изд-во физ.-мат. лит., 1960. 656 с.
- Buckland S. T., Newman K. B., Thoma s L., Koesters N. B. State-space models for the dynamics of wild animal populations//Ecological Modelling. 2004. Vol. 171. P. 157-175.
- Csetenyi A. I., Logofet D. O. Leslie model revisited: some generalizations for block structures//Ecological Modelling. 1989. Vol. 48. P. 277-290.
- Newman K. B., Buckland S. T., Lindley S. T., Thomas L., Fernandez C. Hidden process models for animal population dynamics//Invited Paper for Special Feature of Ecological Applications. 2006. Vol. 16. P. 74-86.
