Дифференцированная оценка антибиотиков на способность ограничивать скольжение Mycobacterium smegmatis

Автор: Цыганов Иван Вадимович, Нестерова Лариса Юрьевна, Ткаченко Александр Георгиевич
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 4, 2018 года.
Бесплатный доступ
Предложен метод дифференцированной оценки веществ, обладающих антибактериальным действием и воздействующих на способность подавлять скольжение Mycobacterium smegmatis за счет влияния на жизнеспособность клеток и/или ограничения подвижности. С помощью данной методики произведена оценка антибиотиков, обладающих антимикобактериальным действием. Показано, что рифампицин в сублетальных концентрациях ограничивает скольжение М. smegmatis, не оказывая действия на жизнеспособность бактерий. В свою очередь, тетрациклин и стрептомицин ингибировали процесс скольжения за счет бактерицидного или бактериостатического действия. Данный подход может быть использован для установления механизма действия новых антибиотиков, а также для изучения их эффекта на свойства клеточной поверхности микобактерий и других микроорганизмов, способных к скольжению.
Микобактерии, скольжение, антибиотики
Короткий адрес: https://sciup.org/147227048
IDR: 147227048 | УДК: 579.22 | DOI: 10.17072/1994-9952-2018-4-402-408
Differentiative evaluation of antibiotics for their ability to limit the sliding motility of Mycobacterium smegmatis
To evaluate some molecular species exhibiting antibacterial activity for their ability to limit the sliding motility of mycobacteria we developed differential method, including the measuring of sliding colony square (mm2) in combination with enumeration of cell number per 1 mm2 of the square. These parameters were used to differentiate inhibition of sliding motility from killing effect of antibacterials and were measured over a range of concentrations. The limiting effect on sliding motility was registered when the square of sliding colony decreased while the cell number per 1 mm2 of its square proportionally increased at subinhibitory concentrations. This effect was observed for rifampicin, however, tetracycline and streptomycin restricted the sliding colony size through their killing effects on Mycobacterium smegmatis cells.
Текст научной статьи Дифференцированная оценка антибиотиков на способность ограничивать скольжение Mycobacterium smegmatis
Изучение механизмов, лежащих в основе координированных действий микробных клеток, направленных на эффективное распространение по поверхности и формирование колониальных и биопленочных сообществ, приобретает все большую актуальность в связи с тем, что такие «поведенческие» реакции сопровождаются образованием толерантных к антибиотикам форм бактерий [Verstraeten et al., 2008; Thayil et al., 2011]. В случае с патогенными микроорганизмами, это сопровождается развитием не поддающихся лечению антибиотиками рецидивирующих хронических инфекций, в особенности вызванных Mycobacterium tuberculosis .
В отличие от жгутиковых микроорганизмов, микобактерии не способны к направленному дви- жению и роению и до недавнего времени считались абсолютно неподвижными. Однако в конце 90-х гг. прошлого века было показано, что микобактерии способны к скольжению (sliding) – особому типу пассивного движения по поверхности [Martinez, Torello, Kolter, 1999], который ранее был описан для других безжгутиковых форм [Henrichsen, 1972]. Распространение скользящей колонии реализуется за счет деления бактерий и возникающей при этом экспансивной силы, которая проталкивает клетки вперед [Daffé, Draper, 1997]. Для облегчения скольжения микроорганизмы могут продуцировать поверхностно активные вещества, уменьшающие трение о поверхность, например рамнолипиды у некоторых грамотрица-
тельных бактерий [Murray, Kazmierczak, 2008]. Хотя скольжение микобактерий не зависит от наличия специфических соединений в матриксе, тем не менее, установлено, что мутанты, у которых в клеточной стенке отсутствовали гликопептидо-липиды, теряли подвижность [Recht et al., 2000; Ghosh, Indi, Nagaraja, 2013]. О регуляции скольжения микобактерий известно немного, однако показано, что переход к скольжению у микобактерий контролируется при участии сигнальных нуклеотидов (p)ppGpp и c-di-GMP, которые не только оказывают влияние на поверхностные свойства клеток, в том числе содержание в их клеточных стенках гликопептидолипидов, но и ответственны за формирование толерантности к антибиотикам [Thayil et al., 2011; Gupta et al., 2016].
Особенности строения многокомпонентной клеточной стенки микобактерий придают им свойства повышенной устойчивости к различным неблагоприятным воздействиям окружающей среды, включая антибиотики и компоненты иммунной системы хозяина. От этого зависит также способность микобактериальных клеток к скольжению и биопленкообразованию, что позволяет им колонизировать субстраты и персистировать в окружающей среде [Etienne et al . , 2005]. В свою очередь, способность к скольжению часто используется для характеристики поверхностных свойств и физиологического состояния микобактерий.
В последнее время активно ведутся поиски эффективных ингибиторов межклеточных взаимодействий, нарушающих функционирование бактериальной системы quorum sensing (QS), агрегацию, адгезию и движение клеток по поверхности субстратов, то есть процессов, играющих основную роль в биопленко-образовании и персистенции. Ранее показано, что некоторые группы соединений могут оказывать значительное влияние на способность микобактерий к скольжению (ПАВ, синтетические гликолипиды, регуляторы метаболизма, биогенные полиамины, антибиотики) [Gopalaswamy et al., 2008; Naresh et al . , 2010; Syal et al . , 2016; Нестерова, Цыганов, Ткаченко, 2017]. Причинами уменьшения диаметра скользящей колонии при их действии может быть не только ингибирование процесса скольжения, но и снижение жизнеспособности бактериальных клеток в результате токсического действия. Это ставит задачу дифференцировать два вида активности при изучении механизма действия исследуемых соединений, что составляет цель данной работы.
Материалы и методы исследования
Объектом исследования служил штамм Mycobacterium smegmatis mc2 155 из коллекции лаборатории адаптации микроорганизмов
Института экологии и генетики микроорганизмов УрО РАН.
Скользящие колонии выращивали в полистироловых чашках Петри (Thermo scientific, 35 мм). В жидкую среду Middlebrook 7H9 (Difco, Франция) добавляли агарозу (Хеликон, Россия) в концентрации 0.3%. Антибиотики вносили в остывшую до температуры 47оС питательную среду, которую разливали по чашкам и подсушивали в течение 40 мин. Культуру M. smegmatis выращивали на питательной среде Middlebrook 7H9 до оптической плотности (ОП) 1.3 (600 нм), разводили до ОП 0.4 (600 нм), наносили на агар в центр чашки Петри каплей (2 мкл) и оставляли в термостате (37оС) на 48 ч.
Подсчет клеток в колонии проводили под микроскопом в камере Горяева. Колонию вырезали вместе с полужидким агаром и помещали в микропробирку, куда вносили физиологический раствор с добавлением tween 80 (0.05%) до конечного объема 1 мл и стеклянные бусы (2 мм, Hofmann Glastechnik, Германия). Микропробирку встряхивали на вортексе (1400 об/мин.; 1ч.). После этого производили подсчет клеток в камере Горяева в 60 малых квадратах по диагонали. Количество клеток на мм2 колонии рассчитывали по формуле
N = (mx4000xs/q)x1000, где N – число клеток в 1 мл раствора; m – сумма посчитанных клеток; s – степень разведения; q – число малых квадратов сетки камеры Горяева, в которых считали клетки.
Измерение площади колонии проводили после фотографирования чашки со скользящей колонией с помощью камеры Olympus C-3040 ZOOM (Япония) с использованием пробной версии программы Adobe Photoshop CC 2015.5, определяя количество пикселей и их площадь.
Минимальную подавляющую концентрацию (МПК) антибиотиков определяли методом двукратных серийных разведений в 96-луночных полистироловых планшетах на среде Middlebrook 7H9 с добавлением tween 80 (0.05%). Приготовление инокулята: культуру M. smegmatis выращивали на среде Middlebrook 7H9 до ОП 1.0 (600нм), затем доводили до ОП 0.1 и разводили ещё в 10 раз. В лунки планшета, содержащие 100 мкл питательной среды с антибиотиком, вносили 100 мкл инокулята. Культивировали 48 ч. в термошейкере Biosan PST-60HL (Латвия) при 37 ° С и 300 об/мин. МПК считали минимальную концентрацию антибиотика, подавляющую видимый рост микроорганизмов.
Для того чтобы определить, сопровождается ли уменьшение размера скользящей колонии уменьшением количества клеток, подсчитали количество бактерий на 1 мм2 скользящей колонии. Эффект ограничения скольжения без бактерицидного действия фиксировали в том случае, если хотя бы при одной из концентраций, достоверно уменьшающих размер скользящей колонии, наблюдалось статистически значимое увеличение количества клеток на мм2 колонии по сравнению с контролем.
Статистическую обработку результатов проводили с использованием пакета стандартных программ Statistica 7.0 (StatSoft, Inc., 2006). На графиках отражены средние значения (4–8 экспериментов), вертикальными отрезками обозначены величины стандартной ошибки среднего. Оценка статистической значимости различий произведена с использованием t-критерия Стьюдента. Различия считали значимыми при р≤0.05.
Результаты
В качестве соединений, способных оказать влияние, как на скольжение, так и на численность клеток в скользящих колониях М. smegmatis , в работе использованы антибиотики, обладающие антимикобактериальной активностью: рифампицин, стрептомицин и тетрациклин.
Действие рифампицина связано с подавлением активности ДНК-зависимой РНК-полимеразы [Се-лизарова, 2003], в то время как стрептомицин и тетрациклин блокируют синтез белка, связываясь с 30S субъединицей рибосомы и нарушая образование комплекса рибосомы с т-РНК соответственно [Epe, Woolley, Hornig, 1987; Егоров, 2004].
Антибиотики добавляли в полужидкую питательную среду, которая использовалась для воспроизведения скольжения микобактерий на чашках Петри (см. раздел «Материалы и методы»). Концентрации антибиотика были подобраны таким образом, чтобы отследить постепенное уменьшение размеров скользящей колонии. Для того, чтобы сравнить между собой эффекты антибиотиков, обладающих разным механизмом действия, их концентрацию выражали в условных единицах, кратных значению минимальной подавляющей концентрации. Для рифампицина она была равна 9.7, стрептомицина – 0.04, а тетрациклина – 0.57.
Результаты исследования показали, что присутствие даже минимальных концентраций рифампицина и тетрациклина в среде (0.06 МПК) приводило к снижению площади колонии по сравнению с контрольным значением приблизительно в 2 и 1.5 раза соответственно (рис. 1, 2). Однако содержание клеток в колонии, выросшей в присутствии этих антибиотиков, существенно различалось. При этом численность клеток на единицу площади колонии в присутствии рифампицина возрастала обратно пропорционально изменению ее площади. В то же время снижение площади колонии в ответ на добавку 0.06 МПК тетрациклина не сопровождалось возрастанием количества клеток на единицу площади, которое оставалось на уровне контрольного. Это свидетельствует о том, что рифампицин в данной концентрации практически не влиял на размножение клеток, но существенно ингибировал их скольжение. В отличие от этого эффект тетрациклина, по-видимому, был вызван ингибировани- ем роста и размножения клеток.
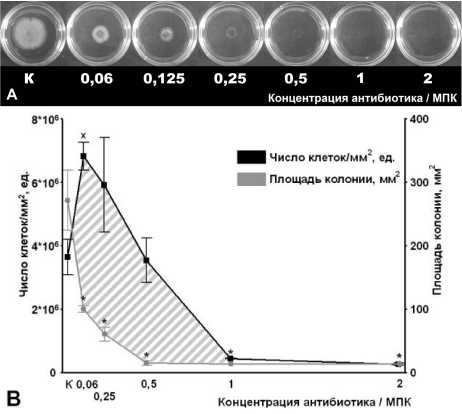
Рис. 1 . Влияние рифампицина на скольжение Mycobacterium smegmatis mc2 155:
МПК – минимальная подавляющая концентрация; К – контроль; А – фото скользящих колоний; В – изменение площади колоний и числа клеток на 1 мм2 колонии; * – статистически значимое отличие площади колонии от контрольной группы (с использованием t-критерия Стьюдента, р≤0.05); х – статистически значимое отличие числа клеток на 1 мм2 от контрольной группы в большую сторону (с использованием t-критерия Стьюдента, р≤0.05)
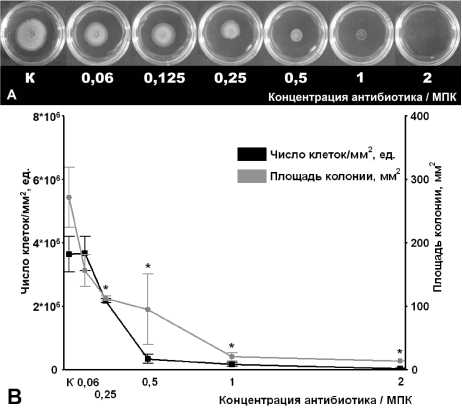
Рис. 2 . Влияние тетрациклина на скольжение Mycobacterium smegmatis mc2 155:
МПК – минимальная подавляющая концентрация; К – контроль; А – фото скользящих колоний; В – изменение площади колоний и числа клеток на 1 мм2 колонии; * -статистически значимое отличие площади колонии от контрольной группы (с использованием t-критерия Стьюдента, р≤0.05
Дальнейшее повышение концентрации рифампицина и тетрациклина подтвердило тенденцию в изменении исследуемых параметров, выявленную при воздействии относительно малых концентраций антибиотиков (рис. 1, 2). При этом повышение концентрации рифампицина вызывало более выраженное ингибирование скольжения по сравнению с изменением количества клеток на единицу площади (рис. 1 – заштрихованная область).
Прямо противоположная тенденция характерна для тетрациклина (рис. 2) и стрептомицина (рис. 3), при действии которых оба параметра менялись сходным образом, что свидетельствует о преобладании бактерицидного или бактериостатического эффекта, вносящего наибольший вклад в снижение диаметра колонии.
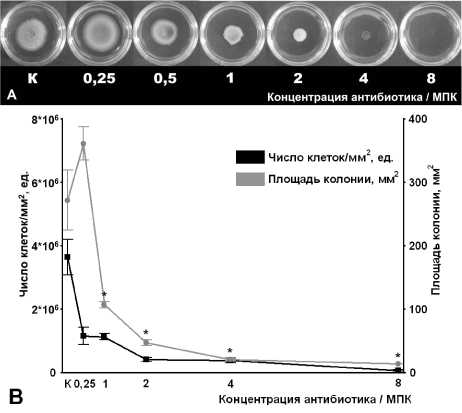
Рис. 3 . Влияние стрептомицина на скольжение Mycobacterium smegmatis mc2 155:
МПК – минимальная подавляющая концентрация; К – контроль; А – фото скользящих колоний; В – изменение площади колоний и числа клеток на 1 мм2 колонии; * – статистически значимое отличие площади от контрольной группы (с использованием t-критерия Стьюдента, р≤0.05)
Обсуждение
Особенности клеток Mycobacterium smegmatis, включая их способность к агрегации во время культивирования, затрудняют определение количества живых организмов в популяции. Использование с этой целью высева образцов культуры на твердую питательную среду для подсчета КОЕ может давать ошибочные результаты. Как показали результаты наших микроскопических исследований, в процессе ресуспендирования скользящей колонии, даже после интенсивного встряхивания с ПАВ, не все клетки микобактерий расходились, частично оставаясь в виде скоплений. Однако используемый нами подсчет в камере Горяева давал возможность точно учитывать не только отдельно расположенные, но и клетки в составе скоплений. Данный метод обеспечивал высокую воспроизводимость результатов. Использование программы Adobe Photoshop, в свою очередь, сделало возможным быстрое и точное определение площади колонии с учетом неровностей её края.
Ранее нами показано, что площадь скользящей колонии может уменьшаться не только в результате торможения деления или бактерицидного действия антибактериальных соединений, но и за счет ингибирующего эффекта не токсичных для бактерий веществ на процесс скольжения [Нестерова, Цыганов, Ткаченко, 2017]. В этом случае действующие вещества могут, проникая в клетку, оказывать регуляторное действие на процессы, контролирующие скольжение, а также взаимодействовать с бактериальной поверхностью, изменяя ее свойства, что сказывается как на характере взаимодействия с субстратом, так и на межклеточных контактах.
Сходное действие могут, по-видимому, оказывать также соединения, обладающие противо-микробной активностью, что может быть выявлено при их использовании в субингибиторных концентрациях. Дифференцировать противомик-робный эффект от влияния на уровне межклеточных коммуникаций или взаимодействия между клетками и средой важно для установления механизма действия вновь открываемых антибактериальных соединений. Это становится особенно актуально в связи с предпринимаемыми в последнее время усилиями по поиску антибиотиков, действие которых направлено на подавление процессов, находящихся под регуляторным контролем QS-системы, включая формирование биопленок, вирулентность, координированное перемещение по твердой поверхности, персистенцию и другие [Garrison et al., 2015; Gutiérrez-Barranquero et al., 2015; Нестерова, Ткаченко, Писцова, 2016].
Таким образом, предложенный нами дифференцированный подход к оценке различных соединений по способности воздействовать на бактериальное скольжение с учетом их возможной антибактериальной активности может быть использован для установления механизма действия новых антибиотиков, а также для изучения их эффекта на свойства клеточной поверхности микобактерий и других микроорганизмов, способных к скольжению.
Работа выполнена в рамках госзадания, номер госрегистрации темы 01201353249 и при финансовой поддержке РФФИ и Администрации Пермского края в рамках научного проекта р_а 16-44590279.
Список литературы Дифференцированная оценка антибиотиков на способность ограничивать скольжение Mycobacterium smegmatis
- Егоров Н.С. Основы учения об антибиотиках. М.: Наука, 2004. 528 с.
- Нестерова Л.Ю., Цыганов И.В., Ткаченко А.Г. Роль биогенных полиаминов в регуляции скольжения у микобактерий // Вестник Пермского университета. Сер. Биология. 2017. Вып. 3. С. 304-311.
- Нестерова Л.Ю., Ткаченко А.Г., Писцова О.Н. Изучение механизмов противомикробного действия соединения ( )-erogorgiane // Вестник Пермского университета. Сер. Биология. 2016. Вып. 4. С. 351-355.
- Селизарова Н.О. Антибиотики, нарушающие синтез макромолекул // Обзоры по клинической фармакологии и лекарственной терапии. 2003. № 1 С. 70-78.
- Epe B., Woolley P., Hornig H. Competition between tetracycline and tRNA at both P and A sites of the ribosome of Escherichia coli // FEBS Letters. 1987. Vol. 213. P. 443-447
- Etienne G. et al. The cell envelope structure and properties of Mycobacterium smegmatis mc2155: is there a clue for the unique transformability of the strain? // Microbiology. 2005. Vol. 151. № 6. Р. 2075-2086.
- Daffé M., Draper P. The envelope layers of mycobacteria with reference to their pathogenicity // Advances in microbial physiology. 1997. Vol. 39. P. 131-203.
- Garrison A.T. et al. Halogenated phenazines that potently eradicate biofilms, MRSA persister cells in non-biofilm cultures, and mycobacterium tuberculosis // Angewandte Chemie. 2015. Т. 127, № 49. Р. 15032-15036.
- Ghosh S., Indi S.S., Nagaraja V. Regulation of lipid biosynthesis, sliding motility, and biofilm formation by a membrane-anchored nucleoid-associated protein of Mycobacterium tuberculosis // Journal of bacteriology. 2013. Vol. 195. № 8. P. 1769- 1778.
- Gopalaswamy R.R. et al. Mycobacterium smegmatis biofilm formation and sliding motility are affected by the serine/threonine protein kinase PknF // FEMS microbiology letters. 2008. Vol. 278, № 1. Р. 121-127.
- Gupta K.R. et al. Regulation of growth, cell shape, cell division and gene expression by second messengers (p) ppGpp and c-di-GMP in Mycobacterium smegmatis // Journal of bacteriology. 2016. Vol. 198, № 9. Р. 1414-1422.
- Gutiérrez-Barranquero J.A.et al. Deciphering the role of coumarin as a novel quorum sensing inhibitor suppressing virulence phenotypes in bacterial pathogens // Applied microbiology and biotechnology. 2015. Т. 99, № 7. Р. 3303-3316.
- Henrichsen J. Bacterial surface translocation: a survey and a classification // Bacteriological reviews. 1972. Vol. 36, № 4. P. 478.
- Martínez A., Torello S., Kolter R. Sliding motility in mycobacteria // Journal of bacteriology. 1999. Vol. 181, № 23. P. 7331-7338.
- Murray T.S., Kazmierczak B.I. Pseudomonas aeruginosa exhibits sliding motility in the absence of type IV pili and flagella // Journal of bacteriology. 2008. Vol. 190, № 8. P. 2700-2708.
- Naresh K. et al. Synthetic arabinomannan glycolipids and their effects on growth and motility of the Mycobacterium smegmatis // Organic & biomolecular chemistry. 2010. Vol. 8, № 3. Р. 592- 599.
- Recht J. et al. Genetic Analysis of Sliding Motility in Mycobacterium smegmatis // Journal of bacteriology. 2000. Vol. 182, № 15. P. 4348-4351.
- Reen F.J. et al. The sound of silence: activating silent biosynthetic gene clusters in marine microorganisms // Marine drugs. 2015. Vol. 13, № 8. Р. 4754-4783.
- Syal K. et al. Synthetic arabinomannan glycolipids impede mycobacterial growth, sliding motility and biofilm structure // Glycoconjugate journal. 2016. Vol. 33, № 5. Р. 763-777.
- Thayil S.M. et al. The role of the novel exopolyphosphatase MT0516 in Mycobacterium tuberculosis drug tolerance and persistence // PloS one. 2011. Vol. 6, № 11. Р. e28076.
- Verstraeten N. et al. Living on a surface: swarming and biofilm formation // Trends in microbiology. 2008. Vol. 16, № 10. P. 496-506.
