Диморфизм цветков Jasminum fruticans L. (Oleасеае)
")
Автор: Кузьмина Т.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2017 года.
Бесплатный доступ
Представлены результаты сравнительного анализа длинностолбчатых и короткостолбчатых цветков дистильного вида флоры Крыма Jasminum fruticans L. (Oleaceae). Выявлены различия размеров венчиков, пыльников, столбика, пыльцевых зерен и разница длин тычинок цветков разных морфологических форм. Показано, что у длинностолбчатых цветков тычинки и их пыльники, а также пыльцевые зерна значительно меньше, чем у цветков короткостолб-чатой формы. Цитоморфологическая оценка пыльцевых зерен цветков двух флоральных форм J. fruticans не выявила значимых различий между ними по долям морфологически нормальных пыльцевых зерен, которая составила более 80%, что свидетельствует об отсутствии серьезных аномалий в ходе развития мужского гаметофита и равных возможностях мужской генеративной сферы независимо от морфологической формы цветка.
Диморфизм, полиморфизм, гетеростилия, цветок, пыльцевые зерна
Короткий адрес: https://sciup.org/147204830
IDR: 147204830 | УДК: 582.916.6:581.46
The dimorphism of flowers of Jasminum fruticans L. (Oleaceae)
A comparative analysis of long-styled and short-styled flowers of the distylous species of the Crimean flora Jasminum fruticans L. {Oleaceae) is given. The differences of the corolla length, anthers and stigma heights, stigma-anther separation and pollen grains were found for two plants morphological forms. The stamens, anthers and pollen grains are larger in short-styled flowers. The cytomorphological analysis of pollen grains of J. fruticans flower two morph did not reveal the significant differences between the proportions of morphologically normal pollen grains (about 80%). The state of male gametophyte J. fruticans shows absence of lethal anomalies of its development. A cytomorphological status of pollen does not depend on the morphs of the flower. The functionality of male generative sphere of long-styled and short-styled flowers is equal.
Текст научной статьи Диморфизм цветков Jasminum fruticans L. (Oleасеае)
Согласно современным данным, гетеростилия, или разностолбчатость, встречается у представителей более 175 родов из 36 семейств покрытосеменных растений [Ganders, 1979; Агаджанян, 2000]. В их число входит род Jasminum L. (сем. Oleaceae), для которого типична дистилия, являющаяся частным случаем гетеростилии. При дистилии у представителей одного вида существуют особи с цветками двух морфологических форм – с длинными и короткими столбиками. В цветках длинностолбчатой формы тычинки располагаются в глубине трубки венчика, а рыльце столбика выступает из трубки венчика. В случае короткостолбчатой формы пыльники находятся в зеве цветка, а столбик пестика – в трубке венчика. Различия между фло-ральными формами также могут выражаться в размерах цветков, строении поверхности рылец пестика, в количестве цветков в соцветиях и ряде других признаков [Ganders, 1979; Жинкина, 2000; Демьянова, 2014].
Jasminum fruticans L. – кустарник, имеющий европейско-средиземноморско-переднеазиатский ареал. В Крыму этот вид произрастает в горном и южнобережном районах [Голубев, 1992].
Описание цветков J. fruticans с учетом ряда морфометрических параметров двух флоральных форм представлено в работах J.D. Thompson, B. Dommée [1993; 2000] и J. Guitiána с соавторами [1998]. Было показано, что пыльники короткостолбчатых цветков J. fruticans производят большее количество пыльцы, которая крупнее, чем у цветков длинностолбчатой формы [Guitián et al., 1998], также был выявлен изгиб столбика, свойственный длинностолбчатым цветкам [Thompson, Dommée, 2000].
Одним из наиболее распространенных объяснений морфологических различий у цветков гетеростильных растений является их функциональное приспособление к легитимному опылению между цветками различных морфологических форм [Фегри, Ван дер Пейл, 1982]. Подтверждением этому является характерная для ряда гетеростильных видов, в том числе и J. fruticans , как самонесовместимость, так и внутриморфная несовместимость [Dommée, Thompson, 1992; Guitián et al., 1998; Dommée, Thompson, 2000]. Однако у некоторых гетеростильных видов самонесовместимость четко не прослеживается [Фегри, Ван дер Пейл, 1982; Методические указания, 1986; Демьянова, 2014], что не позволяет ограничивать роль гетеростилии исключительно механизмом, гарантирующим ксе-ногамное опыление между разными флоральными формами.
Существует также мнение, что гетеростилию можно рассматривать как эволюционное направление, ведущее к формированию однополых рас- тений [Baker ,1960; Фегри, Ван дер Пейн, 1982]. А поскольку известно, что формирование двудомности у покрытосеменных растений сопряжено с редукцией генеративных органов [Кордюм, Глущенко, 1976] то можно предположить, что полиморфизм генеративных элементов у гетеростильных растений сопряжен с редукцией структурных элементов мужской и женской генеративной сферы, в зависимости от морфологической формы цветка, что, в свою очередь, может оказывать влияние на состояние их гаметофитов.
Целью данной работы был сравнительный анализ морфометрических параметров с учетом оценки состояния мужского гаметофита длинностолбчатых и короткостолбчатых цветков J. fruticans , произрастающих на Южном берегу Крыма.
Объект и методы исследований
В ходе исследования было проанализировано по 100 цветков J. fruticans каждой морфологической формы, взятых с растений, произрастающих на территории Никитского ботанического сада (г. Ялта, Крым), в период массового цветения растений (II декада мая) в 2016 и 2017 гг. Измерения проводили по следующим параметрам: высота чашечки и трубки венчика; длина отгиба венчика, столбика с рыльцем, тычинок, пыльника; для каждого цветка вычисляли разницу длин столбика и тычинки и подсчитывали количество лепестков и тычинок (рис. 1).
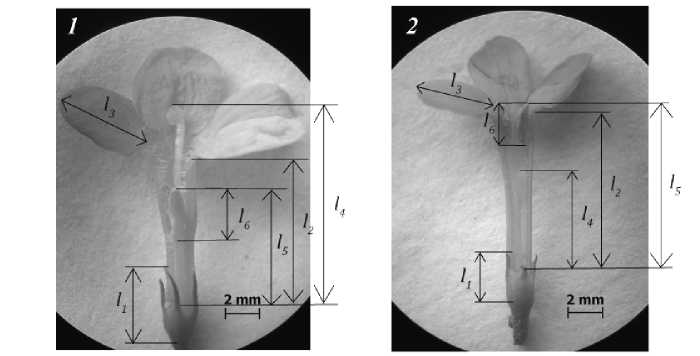
Рис. 1 . Морфометрические характеристики длинностолбчатых ( 1 ) и короткостолбчатых ( 2 ) цветков J. fruticans :
l 1 – высота чашечки; l 2 – высота трубки венчика; l 3 – длина отгиба венчика; l 4 – длина столбика с рыльцем; l 5 – длина тычинки; l 6 – длина пыльника
Для определения экваториальных диаметров пыльцевых зерен использовали постоянные препараты средних образцов пыльцы, которые окрашивали метилгрюнпиронином [Шевченко и др, 1986]. Анализ постоянных препаратов проводили методом светлопольной микроскопии на микроскопе
AxioScopeA.1 (Zeiss, Германия). Микрофотографии получены с помощью системы анализа изображения AxioCamERc5s и цифровой фотокамеры Olympus SP-350. Для морфометрических измерений использовали программное приложение Axio-VisionRel. 4.8.2., а для проведения цитоморфоло- гической оценки пыльцевых зерен – программу ImageJ 1.48v .
Статистическую обработку данных, включающую дескриптивную статистику, корреляционный и однофакторный дисперсионный анализы (ANOVA) проводили с использованием программного приложения Statistica 6.0. Среднее значение показателей приводятся в виде М±SE, где М – среднее, SE – стандартная ошибка, SD – стандартное отклонение; CV – коэффициент вариации. Статистически значимыми приняты различия при p<0.05. Оценку разности выборочных долей морфологически нормальных и стерильных пыльцевых зерен проводили методом преобразования долей в углы φ по критерию Фишера (F φ ) [Плохин-ский, 1970].
Результаты и их обсуждение
Цветок у J. fruticans зигоморфный, с яркожелтым трубчатым венчиком, рассеченным на 4–8 долей отгиба. При этом в среднем для цветков обоих типов характерен венчик с 5 долями. Однако у проанализированных нами цветков длинностолбчатой формы чаще встречается от 6 до 8 долей. В цветке J. fruticans , как правило, две тычинки, хотя встречаются цветки и с 3 тычинками. Доля таких цветков длинностолбчатой формы составляет 12%, в то время как у короткостолбчатых цветков формирование трех тычинок наблюдалось в 6% всех исследованных нами цветков данной флоральной формы.
У цветков сравниваемых морф прослеживаются различия по форме рыльца: у длинностолбчатых цветков рыльце головчатой формы, а у цветков короткостолбчатого типа оно плоское, расчлененное на две лопасти (рис. 2).
Морфометрические параметры длинно- и короткостолбчатых цветков J. fruticans представлены в табл. 1. Установлены статистически значимые различия между высотой трубки венчика, длинами столбика, тычинок, пыльников, а также разницы длин столбика и пыльников у цветков сравниваемых морфологических форм. Так, у длинностолбчатых цветков, как правило, высота трубки венчика больше, по сравнению с короткостолбчатыми цветками (F=33,73>Fst=3.9 при p<0.05). В соответствии с морфологическим типом цветки различаются по длине столбика (F=1211,62>Fst=3.9 при p<0.05) и тычинок (F=289,41>Fst=3.9 при p<0.05). По длине пыльников также выявлены различия: у цветков короткостолбчатого типа пыльники в среднем на 0.6 мм длиннее пыльников длинностолбчатых цветков (F=43,46>Fst=3.9 при p<0.05). Сравнение значений коэффициентов вариации (CV), характеризующих степень изменчивости признаков, показало, что у короткостолбчатых цветков J. fruticans длина столбика обладает большей изменчивостью, чем у длинностолбчатых цветков, что свидетельствует о большой стабильности данного признака у последнего и согласуется с литературными данными как для J. fruticans, так и для других гетеростильных видов [Thompson, Dommée, 2000]. В то же время, для длинностолбчатых цветков характерна большая вариабельность длин тычинок, чем у цветков короткостолбчатой формы. Таким образом, у длинностолбчатых цветков более стабильным признаком является длина столбика, а у короткостолбчатых – длина тычинок. При этом проведенный нами корреляционный анализ выявил, что у длинностолбчатых цветков J. fruticans длина столбика и тычинок находятся в тесной положительной взаимосвязи с высотой трубки венчика (r=0.83; r=0.86, соответственно), у короткостолбчатых цветков зависимость высоты трубки венчика и длин генеративных элементов оценивается как значительная (r=0.54; r=0.60). При этом у исследуемых нами цветков длинностолбчатых растений высота трубки венчика была больше, чем у короткостолбчатых, а по длине отгиба статистическое различие между флоральными формами не установлено.
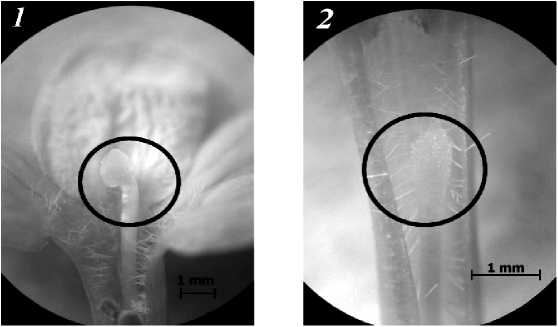
Рис. 2. Рыльце длинно- (1) и короткостолбчатого (2) цветков J. fruticans
Таблица 1
Морфометрические параметры длинно- и короткостолбчатых цветков Jasminum fruticans
|
Признак |
Длинностолбчатые цветки |
(n=100) |
Короткостолбчатые цветки |
(n=100) |
||||
|
M |
SD |
SE |
CV |
M |
SD |
SE |
CV |
|
|
Высота чашечки с зубцами, мм ( l 1 ) |
4.11 |
0.62 |
0.06 |
15.09 |
4.03 |
0.75 |
0.08 |
18.73 |
|
Высота трубки венчика, мм ( l 2 ) |
9.82 |
1.57 |
0.16 |
16.02 |
8.79 |
0.82 |
0.08 |
9.32 |
|
Длина отгиба венчика, мм ( l 3 ) |
5.96 |
1.06 |
0.11 |
17.84 |
5.82 |
0.87 |
0.09 |
14.99 |
|
Длина столбика, мм ( l 4 ) |
11.74 |
1.59 |
0.16 |
13.57 |
5.15 |
1.02 |
0.10 |
19.87 |
|
Длина тычинки, мм ( l 5 ) |
7.56 |
1.23 |
0.12 |
16.28 |
10.22 |
0.97 |
0.10 |
9.53 |
|
Длина пыльника, мм ( l 6 ) |
2.56 |
0.57 |
0.06 |
22.40 |
3.12 |
0.63 |
0.06 |
20.11 |
|
Разница длины столбика и тычинки, мм |
4.17 |
0.89 |
0.09 |
21.24 |
5.07 |
1.13 |
22.32 |
0.11 |
Выявленные нами различия расходятся с данными, представленными в ряде работ, анализирующих строение цветков различных видов гете-ростильных растений [Pailler, Thompson, 1997; Faivre, McDave, 2001; Naiki, Nagamasu, 2003]. Согласно мнению J.D. Thompson и B. Dommée [2000], увеличение трубки венчика у короткостолбчатых цветков способствует улучшению взаимодействия генеративных органов при фиксированном положении тычиночной нити в трубке венчика и наблюдается у ряда гетеростильных видов различных семейств. Однако не у всех гетеро-стильных видов отмечают такую взаимосвязь, в частности, у Anchusa officinalis L. ( Boraginaceae ) значимых различий в размерах венчика у морфологических форм цветков не выявлено [Philipp, Schou, 1981]. Таким образом, утверждение о четкой зависимости размеров цветка от его морфологической формы не является однозначным, хотя и прослеживается взаимосвязь размеров венчика и морфологической формы цветка.
В целом же выявленные различия длинно- и короткостолбчатых цветков растений J. fruticans , произрастающих на Южном береге Крыма, имеют типичные черты, свойственные представителям рода Jasminum и других гетеростильных родов и семейств покрытосеменных растений [Guanian et all., 1998], которые выражаются в различии длин столбиков, тычинок и их пыльников у цветков разных флоральных форм. Цитометрический анализ пыльцевых зерен цветков J. fruticans показал наличие статистически значимых различий по экваториальному диаметру пыльцы длинностолбчатых и короткостолбчатых цветков. При этом пыльцевые зерна короткостолбчатых цветков крупнее, чем у длинностолбчатых цветков и составляют 46.58±0.25 μm и 36.49±0.25μm, соответственно (рис. 3; рис. 4). Аналогичная закономерность установлена для ряда других гетеростильных видов как рода Jasminum [Olesen et al., 2003], так и других таксонов, в частности, Gaertnera vaginata Lam. (Rubiaceae) [Pailler, Thompson, 1997] и Anchusa officinalis L. (Boraginaceae) [Philipp, Schou, 1981].
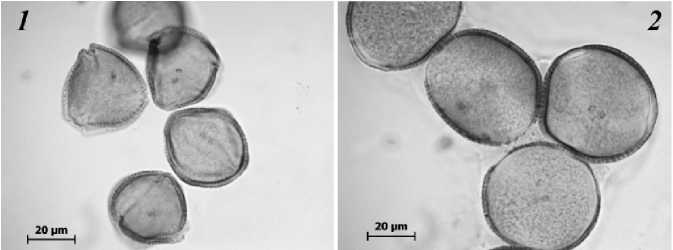
Рис. 3. Пыльцевые зерна длинно- ( 1 ) и короткостолбчатых ( 2 ) цветков Jasminum fruticans
Цитоморфологический анализ пыльцевых зерен длинно- и короткостолбчатых цветков J. fruticans показал, что независимо от флоральной формы цветка в средних образцах пыльцы преобладают морфологически нормальные пыльцевые зерна, доля которых превышает 80%, (Fφ =2.61 нова, 2014], что может свидетельствовать об от- Таблица 2 Цитоморфологическая характеристика пыльцевых зерен J. fruticans Количество Морфологически нор- Морфологическая Стерильные пыльце- проанализированных мальные форма цветка вые зерна, % пыльцевых зерен, шт. пыльцевые зерна, % Fφ Длинностолбчатая 623 87.80 12.20 2.61 Короткостолбчатая 432 84.26 15.74 | 44 I 42 s 40 | 38 321------------------■-------------------------------------■------------------ 1 2 □ Среднее , , □ Среднее±Ст.ош. Морфологическая форма цветка J Среднее±Ст.откл. Рис. 4. Экваториальный диаметр пыльцевых зерен длинностолбчатых (1) и короткостолбчатых (2) цветков J. fruticans Следовательно, выявленные при дистилии J. fruticans различия длинно- и короткостолбчатых цветков, выражающиеся в пространственной разобщенности генеративных элементов, варьировании формы рыльца, уменьшении размеров пыльников и пыльцевых зерен у цветков длинностолбчатого типа, не отражаются на потенциальных возможностях эффективного функционирования мужской генеративной сферы у цветков данной флоральной формы. Подтверждением этому может служить установленное J. D. Thompson, B. Dom-mée [1993] отсутствие различий в показателях семенной продуктивности у длинно- и короткостолбчатых растений J. fruticans. Выводы Диморфизм J. fruticans связан с морфометрическими и морфологическими различиями элементов длинно- и короткостолбчатых цветков. У короткостолбчатых цветков пыльники и пыльцевые зерна крупнее, чем у цветков длинностолбчатой формы. При этом у флоральных форм отсутствуют статистически значимые различия по долям морфологически нормальных и стерильных пыльцевых зерен, что свидетельствует об их потенциаль- но равнозначном участии в опылении и половом процессе. Морфологические различия цветков J. fruticans обусловлены особенностями строения рыльца в зависимости от их флоральной формы: у длинностолбчатых цветков оно головчатой формы, а у цветков короткостолбчатой формы рыльце уплощенное и рассечено на две лопасти.
Список литературы Диморфизм цветков Jasminum fruticans L. (Oleасеае)
- Агаджанян A.M. Распространённость и распределение гетеростилии в системе покрытосеменных растений//Успехи современной биологии. 2000. Т. 120, № 4. С. 348-360
- Голубее В.Н. Биологическая флора Крыма. Ялта: НБС-ННЦ, 1996. 126 с
- Демьянова Е.И. О полиморфизме некоторых гетеростильных растений лесостепного Зауралья//Вестник Пермского университета. Серия Биология. 2014. Вып. 2. С. 10-17
- Жинкина Н.А. Гетеростилия//Эмбриология цветковых растений. Терминология и концепции. СПб., 2000. Т. 3. С. 82-83
- Кордюм Е.Л., Глущенко Г. И. Цитоэмбриологиче-ские аспекты проблемы пола покрытосеменных. Киев: Наукова думка, 1976. 200 с
- Методические рекомендации по изучению антэко-логических особенностей цветковых растений. Морфологическое описание репродуктивной структуры. Ялта, 1986. 43 с
- Плохинский Н.А. Биометрия. М.: Изд-во МГУ, 1970. 367 с
- Фегри К., Ван дер Пейл Л. Основы экологии опыления. М.: Мир, 1982. 379 с
- Шевченко С.В., Ругузов И.А., Ефремова Л.М. Методика окраски постоянных препаратов метиловым зеленым и пиронином//Бюллетень Государственного Никитского ботанического сада. 1986. Вып. 60. С. 99-101
- Baker Н. G. Reproductive methods as factors in speci-ation in flowering plants//Cold Spring Harbor symp. Quant. Boil. 1960. Vol. 24. P. 177-191
- Faivre A.E., McDade L.A. Population-level variation in the expression of heterostyly in three species of Rubiaceae: does reciprocal placement of anthers and stigmas characterize heterostyly?//American Journal of Botany. 2001. Vol. 88, № 5. P. 841-853.
- Ganders F.R. The biology of heterostyly//New Zealand Journal of Botany. 1979. Vol. 17. P. 607-635
- Guitian J., Guitian P., Medrano M. Floral biology of the distylous Mediteranean shrub Jasminum fruticans (Oleaceae)//Nordic Journal of Botany. 1998. Vol. 18. P. 195-201
- Naiki A,. Nagamasu H. Distyly and pollen dimorphism in Damnacanthus (Rubiaceae)//Journal of Plant Research. 2003. Vol. 116. P. 105-113. doi 10.1007/S10265-002-0075-1
- Olesen J.M. et al. Heterostyly in the Canarian endemic Jasminum odoratissimum (Oleaceae)//Nordic Journal of Botany. 2003. Vol. 23, № 5. P. 537-539
- Pailler Th., Thompson J.D. Distyly and variation in heteromorphic incompatibility in Gaertnera vaginata (Rubiaceae) endemic to La Reunion Island//American Journal of Botany. 1997. Vol. 84, № 3. P. 315-327
- Philipp M., Schou O. An unusual heteromorphic incompatibility system. Distyly, self-incompatibility, pollen load and fecundity in Anchusa officinalis (Boraginaceae)//New Phytol. 1981. Vol. 89. P. 693-703
- Thompson J.D., Dommee B. Sequential variation in the components of reproductive success in the distylous Jasminum fruticans (Oleaceae)//Oecologia. 1993. Vol. 94, № 4. P. 480-487 DOI: 10.1007/BF00566962
- Tompson J.D., Dommee B. Morph-specific patterns of variation in stigma height in natural populations of distylous Jasminum fruticans II New Phytol. 2000. Vol. 148. P. 303-314
