Динамика биомассы и численности фукусовых водорослей Кольского залива Баренцева моря

Автор: Малавенда Сергей Сергеевич
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Океанология
Статья в выпуске: 1-2 т.19, 2016 года.
Бесплатный доступ
Рассмотрена многолетняя динамика биомассы и численности поселений Fucus distichus и Fucus vesiculosus в южном и среднем коленах Кольского залива. Впервые выявлены обратные связи между этими параметрами и их изменения в течение нескольких лет. Смена преобладания биомассы и численности может рассматриваться как адаптация на популяционном уровне для сохранения стабильности поселений водорослей в условиях хронического загрязнения.
Кольский залив, биомасса, численность, многолетняя динамика
Короткий адрес: https://sciup.org/14294893
IDR: 14294893 | УДК: 574.34 | DOI: 10.21443/1560-9278-2016-1/2-296-301
Dynamics of biomass and population density of fucus algae of the Kola bay, Barents Sea
The long-term dynamics of biomass and population density of Fucus distichus and F. vesiculosus in the southern and middle knees of the Kola Bay have been analyzed. The feedbacks between these parameters and their changes over several years have been revealed for the first time. Changing the prevalence of biomass and population density can be considered as an adaptation at the population level to maintain the stability of algae communities in chronic pollution.
Текст научной статьи Динамика биомассы и численности фукусовых водорослей Кольского залива Баренцева моря
Исследование состояния сообществ фукусовых водорослей является обязательным при оценке воздействия на окружающую среду (ОВОС), подготовке различных проектов хозяйственной деятельности и других инженерных изысканиях согласно перечню видов флоры и фауны, являющихся индикаторами устойчивого состояния морских экосистем Арктической зоны РФ 1 , а также свода правил "Инженерные изыскания для строительства. Основные положения" 2 .
Интерпретация данных о численности и биомассе поселений фукоидов осложняется сезонными и многолетними изменениями этих параметров, о динамике которых известно крайне мало. Фукусовым водорослям, как и большинству макрофитов арктической и субарктической зон, свойственна сезонная динамика развития и эндогенная регуляция [1; 2]. Однако нельзя не учитывать возможные колебания популяционных характеристик в течение более длительного периода, обусловленные естественными процессами, характерными для всех долгоживущих организмов. Недооценка этих изменений может привести к ошибочным выводам, особенно при исследовании районов, испытывающих хроническое загрязнение, каким является, например, Кольский залив.
Цель данной работы – исследование межгодовой и сезонной динамики биомассы и численности Fucus distichus и Fucus vesiculosus в южном и среднем коленах Кольского залива.
Материалы и методы
Для исследования были использованы пробы Fucus. vesiculosus и Fucus distichus, отобранные на четырех участках Кольского залива (в южном колене – мыс Притыка, Абрам-мыс; в среднем – мыс Мишуков, бухта Белокаменная) (рис. 1). Характеристика участков отбора проб указана в табл. 1.
Пробы отбирали в периоды гидрологических сезонов в течение двух лет (2007–2009 гг.). На каждом участке литорали сбор проб проводили методом вертикальных трансект с использованием рамки площадью 0,25 м 2 [3] в 6-кратной повторности. При камеральной обработке проб определяли плотность зарослей (численность особей на м 2 ) и биомассу.
При статистической обработке использовали методы описательной статистики и корреляционного анализа по Спирмену. Анализ данных проводили в программном пакете MS Exel 2010 и PAST.
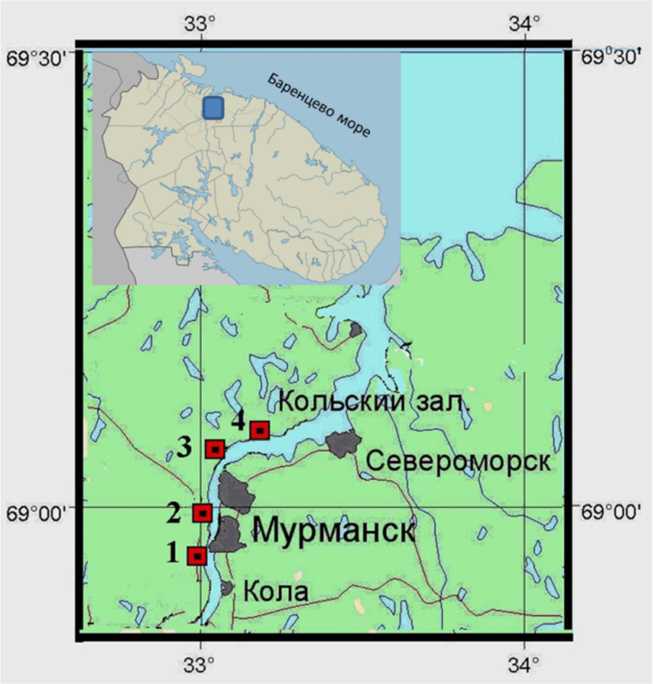
Рис. 1. Расположение пробных площадей на литорали Кольский залива: 1 – мыс Притыка;
2 – Абрам-мыс (южное колено); 3 – мыс Мишуков; 4 – бухта Белокаменная (среднее колено)
Таблица 1
Характеристика участков отбора проб [4]
|
Участок |
Интенсивность движения воды · 10 –3 , мгCaSO 4 /г·ч |
Прибойность, баллы [5] |
Соленость (прилив / отлив), ‰ |
Грунт |
|
Мыс Притыка |
4 |
IV |
11 / 2 |
Песок, ил |
|
Абрам-мыс |
8 |
IV |
30 / 2 |
Валуны |
|
Мыс Мишуков |
8 |
IV |
32 / 25 |
на песчаном основании |
|
Бухта Белокаменная |
3 |
IV |
33 / 25 |
Песчано-илистый и каменистый |
Результаты и обсуждение
Динамика численности и биомассы F. distichus южной части Кольского залива отражена на рис. 1. Колебания этих параметров идут в противофазе друг другу. Максимальная биомасса отмечена в первый год наблюдений, максимальная численность – во второй. Увеличение одного параметра всегда сопровождается уменьшением другого.
Численность и биомасса F. vesiculosus в течение всего периода наблюдений изменяется аналогичным образом (рис. 3).
Корреляционный анализ выявил значимую обратную связь между биомассой и численностью поселений F. distichus, обитающих в районе Абрам-мыса и бухты Белокаменная. Для F. vesiculosus значимая обратная корреляция для тех же параметров обнаружена в поселениях, обитающих на литорали мысов Притыка, Мишуков и бухты Белокаменная (табл. 2).
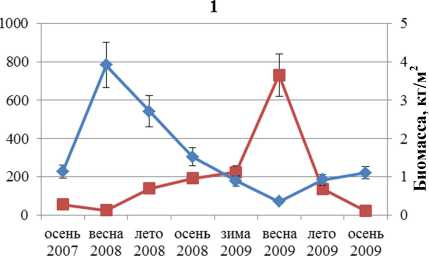
■ Численность ♦ Биомасса
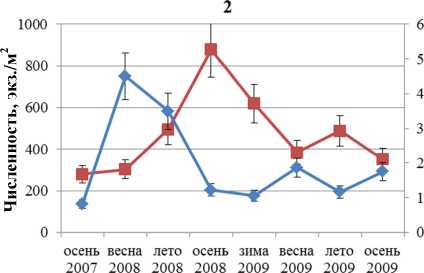
—■—Численность —♦—Биомасса
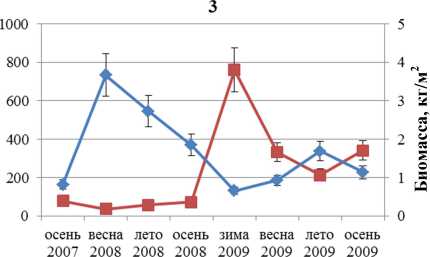
■ Численность ♦ Биомасса
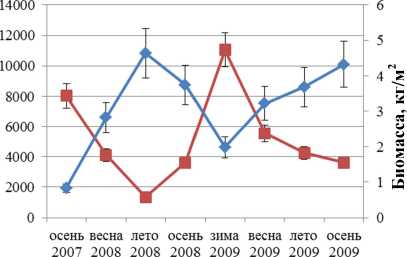
Рис. 2. Биомасса и численность поселений F. distichus литорали Кольского залива: 1 – Абрам-мыс; 2 – мыс Мишуков; 3 – бухта Белокаменная
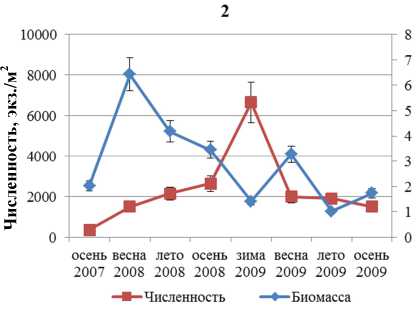
■ Численность ♦ Биомасса
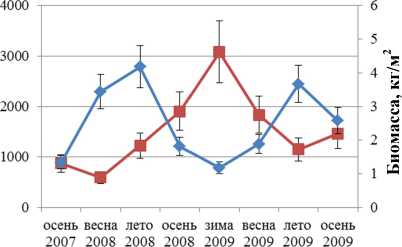
—•—Численность ♦ Биомасса
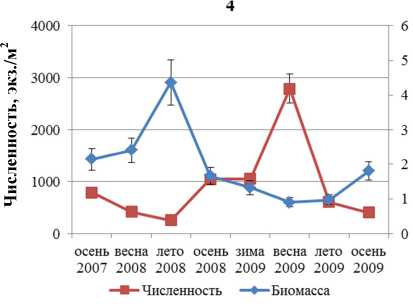
Рис. 3. Биомасса и численность поселений F. vesiculosus литорали Кольского залива: 1 – мыс Притыка; 2 – Абрам-мыс; 3 – мыс Мишуков; 4 – бухта Белокаменная
Биомасса, кг/м 2 Биомасса, кг/м 2 2
Биомасса, кг/м
Таблица 2
Значения коэффициентов корреляции между численностью и биомассой (по методу Спирмена)
|
Участок |
F. distichus |
F. vesiculosus |
||
|
r |
p |
r |
p |
|
|
Мыс Притыка |
– |
– |
–0,898 |
0,002 |
|
Абрам-мыс |
–0,524 |
0,183 |
–0,260 |
0,933 |
|
Мыс Мишуков |
–0,321 |
0,866 |
–0,583 |
0,207 |
|
Бухта Белокаменная |
–0,833 |
0,010 |
–0,762 |
0,028 |
Высокая биомасса при низкой численности поселений фукусовых водорослей обеспечивается за счет преобладания фертильных растений средневозрастных групп, которые являются наиболее адаптированными к постоянному воздействию неблагоприятных факторов среды [6]. Однако затем наблюдается преобладание численности над биомассой, что указывает на упрощение размерно-возрастной структуры. Подобная закономерность была отмечена во многих работах и рассматривается как адаптация популяции фукусовых на эвтрофирование [7–10]. В целом значения биомассы и численности F. vesiculosus и F. distichus на литорали Кольского залива согласуются с исследованиями авторов работ [11; 12; 6; 13], однако большинство данных в цитируемых работах представлены на основании исследований, проведенных в летний период. Как показывает данное исследование, биомасса водорослей зимой снижается в 2–4 раза, при этом численность резко возрастает из-за отмирания взрослых растений, обусловленного образованием ледового припая на литорали. Сходные закономерности отмечались на Белом море [14], однако восстановление биомассы в течение следующего года не происходит, что связано с низкой скоростью роста и длительным жизненным циклом фукусовых водорослей. Таким образом, восстановление биомассы происходит только через два года.
Длительные наблюдения показывают, что смена стратегий существования фукоидов в течение нескольких лет может являться как адаптивным механизмом к антропогенному воздействию, так и естественной динамикой популяции, что обеспечивает стабильность существования. Обратно зависимые колебания биомассы и численности скорее свидетельствуют о стабильности популяции.
Несмотря на высокую степень загрязнения прибрежных вод южного колена Кольского залива 3 , среднемноголетние значения биомассы и численности F. distichus в исследованных районах практически одинаковы (табл. 3), следовательно, градиентного влияния антропогенного загрязнения на этот вид нет. Данный вид водорослей не обитает в кутовой части залива (мыс Притыка), что связано только с его солеустойчивостью [15].
Таблица 3
Среднемноголетние значения биомассы и численности водорослей
|
Участок |
F. distichus |
F. vesiculosus |
||
|
Биомасса, кг/м 2 |
Численность, экз./м 2 |
Биомасса кг/м 2 |
Численность экз./м 2 |
|
|
Мыс Притыка |
– |
– |
3,2 ± 0,4 |
5194 ± 1052 |
|
Абрам-мыс |
1,7 ± 0,4 |
190 ± 80 |
2,9 ± 0,6 |
2333 ± 649 |
|
Мыс Мишуков |
2 ± 0,5 |
475 ± 67 |
2,5 ± 0,4 |
1513 ± 268 |
|
Бухта Белокаменная |
1,7 ± 0,4 |
233 ± 84 |
1,9 ± 0,4 |
918 ± 282 |
Биомасса и численность F. vesiculosus снижаются по направлению от кутовой части залива к среднему колену, что сложно объяснить влиянием загрязнения. Для этого вида, как показывают более ранние исследования [15], переменная соленость стимулирует ростовые процессы, следовательно, кутовая часть залива является более оптимальным местом обитания, а отсутствие других многолетних водорослей в этой части залива снижает конкуренцию за субстрат.
Таким образом, для F. vesiculosus и F. distichus Кольского залива антропогенное воздействие не является основным лимитирующим фактором. По-видимому, определяющими являются те же факторы, что и в экологически чистых бухтах и заливах (соленосный режим, субстрат, прибойность) [16], либо концентрации загрязнителей не оказывают ингибирующего действия.
Заключение
Впервые показано, что поселения литоральных водорослей F. vesiculosus и F. distichus Кольского залива имеют не только сезонные ритмы, но и многолетние. В условиях хронического загрязнения обратную связь между биомассой и численностью и их динамику можно рассматривать как адаптацию на популяционном уровне для сохранения стабильности поселения. Среднемноголетние данные показывают, что на популяционном уровне для водорослей, обитающих в Кольском заливе, загрязнение не является лимитирующим фактором.
Поскольку популяционные характеристики фукоидов подвержены многолетним изменениям, то их использование для оценки степени загрязнения на основании одномоментных исследований является некорректным.
Список литературы Динамика биомассы и численности фукусовых водорослей Кольского залива Баренцева моря
- Макаров В. Н., Шошина Е. В. Динамика и стратегия сезонного роста//Промысловые и перспективные для использования водоросли и беспозвоночные Баренцева и Белого морей. Апатиты: Изд-во КНЦ РАН, 1998. С. 97-115.
- Макаров В. Н., Шошина Е. В. Динамика сезонного роста Laminaria saccharina в Баренцевом море//Биология моря. 1996. № 4. С. 238-248.
- Блинова Е. И. Вертикальное распределение и количественный учет макрофитов Айновских островов (Баренцево море). М.-Л.: Наука, 1965. С. 41-56.
- Малавенда С. С., Малавенда С. В. Черты деградации в фитоценозах южного и среднего колен Кольского залива Баренцева моря//Вестник МГТУ. 2012. Т. 15, № 4. С. 794-802.
- Гурьянова Е. Ф., Закс И. Г., Ушаков П. В. Литораль Кольского залива//Тр. Ленингр. об-ва естествоиспытателей. Л., 1930. Т. 60, № 2. С. 17-107.
- Малавенда С. С. Морфофизиологические особенности бурой водоросли Fucus distichus L. В экосистемах Баренцева моря: автореф. дис.. канд. биол. наук. М., 2009. 23 с.
- Хайлов К. М., Парчевский В. П. Иерархическая регуляция структуры и функции морских растений. Киев: Наук. думка, 1983. 256 с.
- Морские макрофиты в градиенте бытового эвтрофирования/С. А. Ковардаков, А. В. Празукин, С. Е. Завалко, М. А. Изместьева, Д. Д. Рыидина//Водные ресурсы. 1984. № 5. С. 88-103.
- Миничева Г. Г. Показатели поверхности водорослей в структурно-функциональной оценке макрофитобентоса (на примере северо-западной части Черного моря): автореф. дис. … канд. биол. наук. Севастополь, 1989. 20 с.
- Завалко С. Е., Шошина Е. В. Многоуровневая морфофизиологическая оценка состояния фукусовых водорослей в условиях антропогенного загрязнения (Кольский залив, Баренцево море)//Вестник МГТУ. 2008. Т. 11, № 3. С. 423-431.
- Малавенда С. В., Шавыкин А. А., Ващенко П. С. Биомасса макрофитобентоса и районы его наибольшей уязвимости от разливов нефти в Кольском заливе//Защита окружающей среды в нефтегазовом комплексе. 2015. № 12. С. 5-12.
- Шошина E. В., Канищева О. В. Возрастная структура поселений Fucus vesiculosus на литорали Кольского залива//Вестник Удмуртского университета. 2015. Т. 25. Вып. 1. С. 40-47.
- Малавенда С. С., Комракова Д. Г., Малавенда С. В. Изменения структуры литоральных фитоценозов Мурмана при антропогенном воздействии//Вестник МГТУ. 2013. Т. 16, № 3. С. 486-492.
- Кузнецов В. В. Белое море и биологические особенности его флоры и фауны. М.-Л.: Изд-во АН СССР, 1960. 320 с.
- Малавенда С. В. Влияние солености на фукусовые водоросли Баренцева моря: автореф. … канд. биол. наук. Мурманск, 2007. 27 с.
- Малавенда С. В., Bоскобойников Г. М., Матишов Г. Г. Роль интенсивности движения воды и солености в формировании структуры популяции Fucus vesiculosus l. (Phaeophyta) Баренцева моря//Доклады Академии наук. 2007. № 3. С. 424-426.
