Динамика численности белобровика Turdus iliacus и степень возврата птиц на места предыдущего гнездования в Карелии (по данным индивидуального мечения)
")
Автор: Хохлова Татьяна Юрьевна, Яковлева Марина Владимировна
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 753 т.21, 2012 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140152740
IDR: 140152740
Changes in the number and return rate in the redwing Turdus iliacus in Karelia (according to individual tagging)
Текст статьи Динамика численности белобровика Turdus iliacus и степень возврата птиц на места предыдущего гнездования в Карелии (по данным индивидуального мечения)
Несмотря на наличие у многих видов птиц выраженной тенденции к возврату взрослых особей на места предыдущего гнездования (Мальчевский 1974; Соколов 1991; Бардин 1993; Паевский 2008; и др.), часть особей и пар занимают в следующем году новые участки, даже если старые остаются свободными. Многолетние исследования в Карелии в заповеднике «Кивач» в 1979-2003 годах и на стационаре Института биологии КарНЦ РАН «Маячино» в 1979-1997 годах позволили проверить, насколько качество биотопов влияет на постоянство гнездовых территорий у белобровика Turdus iliacus – северного вида с полицик-личным размножением.
Работы проводили на контрольных площадях с разным уровнем численности белобровиков, отражающим степень их привлекательности для данного вида, с использованием различных методов отлова и индивидуального мечения гнездящихся дроздов (Хохлова 2009). В «Киваче» контрольная площадка (24 га) была заложена вблизи жилой зоны на участке смешанного леса с высокой плотностью белобровика; поиск гнёзд с целью обнаружения окольцованных птиц, вёлся на расстоянии до 1.5 км от неё. На стационаре «Маячино» в восточном Приладожье работы проводили в субоптимальных для вида условиях, где численность белобровика была на порядок ниже, чем в «Киваче». Здесь под контролем находилась прибрежная полоса лесов (135 га), в пределах которой сухие вырубки, зарастающие хвойно-лиственными молодняками, перемежались с участками не вырубленного старого хвойного леса и небольшими влажными участками в поймах ручьёв и мелиоративных канав. На рубеже 1970-1980-х годов высота подроста на вырубках не превышала 1 м, в 2000-х годах эти насаждения превратились в «жердняки» 35-40летнего возраста, полностью потеряв свою привлекательность для белобровика.
Белобровик – обитатель светлых разреженных лесов, приопушеч-ной и прибрежной полос, кустарниковых зарослей, соседствующих с открытым ландшафтом. Повсеместно избегает густых спелых хвойных массивов и охотно поселяется в садах и городских парках. В Карелии этот дрозд обычен. На оптимальных по условиям площадках его плот- ность доходит до 3-4 пар/га. В субоптимальных местообитаниях отдельные пары могут поселяться в нескольких километрах друг от друга. В Карелии, как правило, небольшие участки, плотно заселённые белобровиком, окружены большими массивами, где он отсутствует или имеет низкую гнездовую плотность. В результате её средние показатели для больших территорий, включающих разные биотопы, относительно невысоки. Так, по учётам в «Киваче», где старовозрастные леса слабо изменились на протяжении последних десятилетий, они колебались по годам от 0.4 до 2.6 пар на 1 км маршрута по (рис. 1 А, Б), составляя при пересчёте на площадь в среднем 15 пар/км2. Средние показатели плотности для мозаичных лесов островного Заонежья составили 2830 пар/км2. В хвойно-лиственных молодняках Прионежья они колебались от 9 до 26 (Зимин, 1976). В парках Петрозаводска средняя плотность 5.4, г. Сортавалы – 25.1 км2 (Сазонов, 1990). В северной Карелии средняя гнездовая плотность – 9.5 пар/км2, хотя в оптимальных местообитаниях (заросли кустарников) даже на Кольском полуострове она может достигать 120 пар/км2 (Шутова и др. 1993).
Б
А
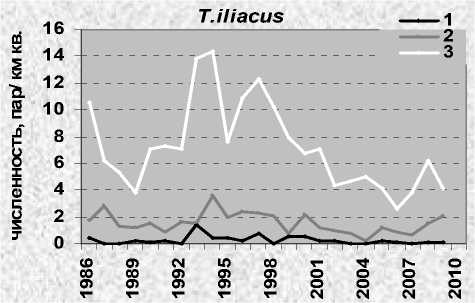
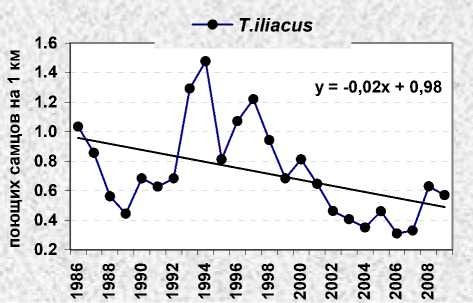
Рис. 1. Численность белобровика Turdus iliacus в лесах разного типа в заповеднике «Кивач». А: 1 – сосняки, 2 – ельники, 3 – лиственные и смешанные леса; Б: все биотопы
В период наших исследований гнездовая плотность белобровика в Карелии, в том числе в оптимальных местообитаниях заповедника «Кивач» (Яковлева 2011), была подвержена большим колебаниям с общей тенденцией к её понижению. Её динамика полностью соответствовала общему ходу изменений численности европейской популяции вида (European… 2010). Максимальные показатели зарегистрированы на рубеже 1970-х и 1980-х годов (рис. 2). На контролируемой площади в «Киваче» они доходили до 112.5 пар/км2 (1982), в «Маячино» – до 34.0 пар/км2 (1979). Потом началось снижение численности с отдельными небольшими и несинхронными подъёмами, которое с 1985 года перешло в затяжную депрессию, спровоцированную экстремальной обстановкой на зимовках в Европе (Väisanen et al. 1989). Несколько лет подряд неудачные зимовки сочетались с неблагоприятными условиями в гнездовое время (холодная дождливая погода, бедная кормовая база), что усугубило ситуацию. В 1988 году гнездовая плотность белобровика в Карелии достигла минимума, опустившись до 20.8 пар/км2 на площадке в «Киваче» и 2.3 пар/км2 – в «Маячино». Аналогичная картина с незначительными отклонениями наблюдалась в 1980-е годы и на соседних территориях Финляндии (Väisanen, Routasuo 1992), а также в Псковской области (Бардин 2008). После этого европейская популяция стала восстанавливаться, но пока так и не достигла первоначального уровня. Процесс восстановления шёл с подъёмами и спадами. В «Киваче» резкие повышения численности отмечены в 1990 и 1993 годах, небольшие падения – в 1991 и 1992 и более глубокое (до уровня предыдущей депрессии) – в 1995 и 1999 годах. В «Маячино» аналогичные колебания показателей происходили на фоне постепенного снижения уровня численности, вызванного лесной сукцессией и обезвоживанием площадей. В 1997 году в окрестностях стационара найдена всего одна гнездящаяся пара, со следующего года здесь регистрировали только проходящих птиц (рис. 2).
120 m
5 100
га 80 с
Л о 60 о Z ф 40 5
т 20
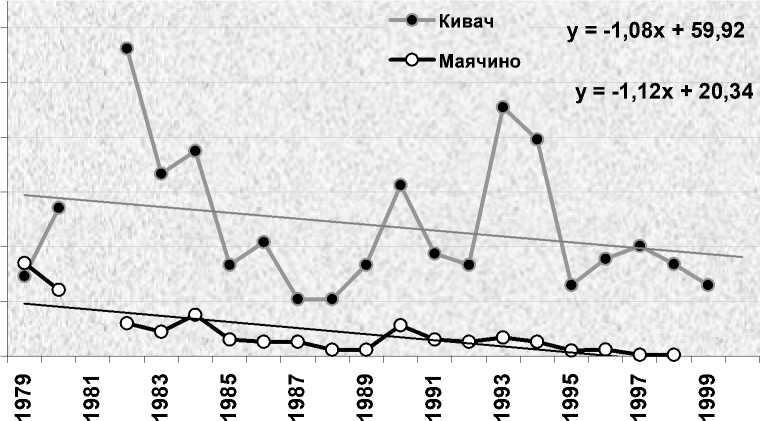
Рис. 2. Численность белобровика Turdus iliacus на контролируемых площадях в заповеднике «Кивач» (24 га) и субоптимальных биотопах в Маячино (135 га).
Размещение гнездящихся пар даже при высокой численности вида очень неравномерно из-за склонности белобровиков к парцеллярному распределению (Tiainen 1977; Данилов и др. 1984; Рябицев 1993; и др.). На контролируемой площади в «Киваче» гнёзда некоторых пар в парцеллах располагались всего в 10-20 м друг от друга. Самцы регулярно устраивали соревновательное пение, однако большой агрессивности к соседям не проявляли, поддерживали их тревогу при опасности и в случае необходимости вместе атаковали приближающегося врага.
На контролируемой территории в Приладожье с низкой плотностью населения белобровика, составившей в годы его регулярного гнездования (1979-1994) в среднем 9.6 пар/км2, картина была несколько иной (рис. 3). Внутри площади выделялся участок сырого черноольшаника (25 га) с густым травяным покровом, который заселялся с более высокой средней плотностью – 14.4 пар/км2, что втрое ниже, чем в те же годы на контрольной площадке в «Киваче» (52.8 пар/км2). На остальной территории, где белобровики занимали относительно удобные места, расположенные далеко друг от друга, преимущественно в годы подъёмов численности этот показатель составил всего 5.6 пар/км2 (Хохлова, Яковлева 1995).
го 12
о 10
т 6
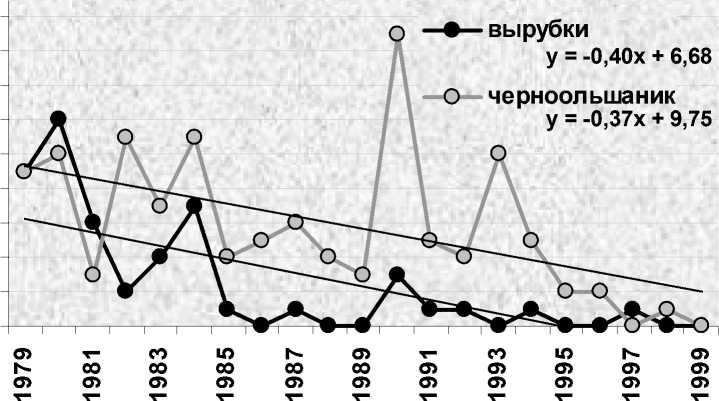
Рис. 3. Число гнездящихся пар белобровика Turdus iliacus на контролируемых площадях в «Маячино» в ежегодно (черноольшаник, 25 га) и не ежегодно (вырубки, 110 га) заселяемых биотопах
Качество участка, занимаемого парой, от которого зависело, в том числе, наличие или отсутствие близких соседей, накладывало отпечаток на многие стороны жизни и поведение белобровиков. Прежде всего, оно сказывалось на широте их перемещений как при добывании корма и вождении выводка, так и при поисках нового партнера в случае потери старого, что особенно значимо для видов с полицикличным размножением.
Размеры территории, которую белобровики посещали во время выкармливания птенцов, зависели от расположения ближайших кормовых угодий. Если гнёзда располагались внутри или вблизи них, родители, избегавшие собирать корм у самого гнезда, летали относительно недалеко – за 100-150 м. При скудной кормовой базе или пересыхании почв в середине лета (особенно на вырубках) они могли улетать в поисках червей за 300-500 м и далее. Сюда же родители уводили слёт- ков, которые иногда задерживались здесь ещё на 2-3 недели. О первых выводках заботились в основном самцы. Самки же сразу приступали ко второму гнездованию (табл. 1), нередко начиная постройку нового гнезда, длящуюся 4-7 дней, а иногда и откладку яиц ещё до вылета птенцов первых выводков (Захарова, Хохлова 1987).
Таблица 1. Продолжительность одного репродуктивного цикла и интервала между началом первых и вторых кладок у белобровика Turdus iliacus
|
Интервал между: |
Число пар |
Продолжительность, сут |
||
|
Min |
Max |
M ± m |
||
Началом кладки и вылетом птенцов 153 22 31 25.7+0.13
Началом кладок первого и второго цикла 63 24 55 33.3+0.82
Расстояние между первым и вторым гнёздами зависело от качества территории. Там, где на протяжении всего сезона сохранялись хорошие кормовые условия (прежде всего обилие дождевых червей), вторые гнёзда обычно располагались недалеко от первых. В случаях, когда кормовые угодья оказываются слишком далеко, птицы, переместившись туда вместе с первым выводком, оставались там для второго размножения. Поэтому расстояние между гнёздами двух репродуктивных циклов одной пары в оптимальных местообитаниях «Кивача» меньше, чем в субоптимальных в «Маячино» (табл. 2). Различия статистически значимы ( χ 2 = 12.16, df = 2, P < 0.01).
Таблица 2. Расстояние между гнёздами первого и второго циклов размножения у белобровиков Turdus iliacus в оптимальных («Кивач») и субоптимальных («Маячино») местообитаниях
|
Расстояние между гнёздами, м |
Число пар |
|||
|
«Кивач» |
«Маячино» |
|||
|
Абс. |
% |
Абс. |
% |
|
|
0–50 |
27 |
71.1 |
7 |
26.9 |
|
51–100 |
4 |
10.5 |
8 |
30.8 |
|
101–200 |
6 |
15.8 |
5 |
19.2 |
|
201–300 |
1 |
2.6 |
2 |
7.7 |
|
301–500 |
– |
– |
2 |
7.7 |
|
501–1000 |
– |
– |
2 |
7.7 |
|
Всего: |
38 |
100.0 |
26 |
100.0 |
Дальность перемещений птиц между циклами, по-видимому, может быть значительно больше приведённой в таблице 2, на что указывают регистрируемые вплоть до начала июля факты появления на площадях новых активно поющих самцов и пар, сразу приступающих к размножению. Высказывают предположение, что миграции в репродуктивный период предпринимают в первую очередь птицы, потерявшие свои гнёзда или выводки (Большаков, Резвый 1981; Рябицев 1993; Резвый, Большаков 1987; Зимин 1988). Однако так ведут себя не все особи: в Карелии многие пары белобровиков, у которых первый цикл закончился неудачей, повторно гнездились на том же участке или недалеко от него (Хохлова, Яковлева 2009). В Псковской области на стационаре «Осыно» в оптимальных для вида стациях белобровики даже в случае гибели кладки или выводка строили новое гнездо обычно в пределах 10-30 м от разорённого. Максимальная дистанция составила 130 м при том, что метод проведения исследования позволял обнаруживать меченых птиц на расстоянии до 3.5 км (Головань 1996). Прослеживание судеб отдельных меченых особей также подтвердило сильную привязанность белобровиков, особенно самцов, к выбранному гнездовому участку в оптимальных биотопах (Головань 1997). Не исключено, что уходят, прежде всего, дрозды, потерявшие партнёра до завершения гнездового сезона и не нашедшие нового поблизости. Во всяком случае, в 1993 году годовалый самец, самка которого погибла незадолго до вылета птенцов из майского гнезда, докормив выводок, навсегда покинул окрестности стационара «Маячино». Это подтверждает также и очень малое число регистраций (2) гнездования на площадях одной самки с разными партнёрами в течение одного сезона. Однако более вероятно, что у белобровика в этом плане нет жёсткого поведенческого стереотипа, что, как и высокая пластичность гнездо-строения (Хохлова, Яковлева 2009), способствует поддержанию их популяций в высоких широтах с коротким северным летом.
В последующие сезоны не все птицы появляются на местах прошлогоднего гнездования. Детальный анализ ситуации с белобровиком в «Маячино» показал, что на сухие вырубки они возвращаются вдвое реже, чем на участок влажного черноольшаника и в пойму ручья, несмотря на то, что контролируемая площадь вырубок в 3 раза больше. Различия долей вернувшихся птиц статистически значимы (критерий Фишера с φ -преобразованием исходных данных F = 5.85, df = 1, 127, P < 0.05). Это касалось как впервые гнездящихся особей, так и птиц, пришедших сюда в старшем возрасте (табл. 3). При этом на свои участки в черноольшанике некоторые птицы прилетали до 4 лет подряд, тогда как на вырубку дважды вернулся лишь один самец.
На самых «неудобных» участках, заселяемых не ежегодно, обычно оказывались пришлые особи, не знакомые с данной территорией, среди которых преобладали впервые гнездящиеся птицы. Вследствие этого процент взрослых опытных особей был выше в биотопах с лучшими условиями. Различия в возрастной структуре населения местообитаний разного качества демонстрируют и другие виды птиц в разных регионах, включая Карелию (Зимин 1978, 1988; Grüll 1981; Шутов 1989; Хохлова, Яковлева 1995; Артемьев 2008; и др.). Это послужило основанием для «буферной» гипотезы, согласно которой весной при распределении гнездовых участков молодые птицы вытесняются из лучших угодий конкурирующими с ними старыми особями (Kluyver, Tinbergen 1953; Verbeek 1984). Внутривидовой конкуренцией объясняют в том числе и особенности территориального распределения другого вида семейства Turdidae – каменки-плясуньи Oenanthe isabellina, у которой оптимальные местообитания занимают двухлетние, а субоптимальные – годовалые особи (Иваницкий 1981).
Таблица 3. Возврат белобровиков Turdus iliacus в «Маячино» на участки разной привлекательности для вида
|
Возраст |
Число особей (возвратов) |
|||
|
Черноольшаник и пойма ручья (35 га) |
Вырубки (100 га) |
|||
|
Отловлено |
Из них возвращалось |
Отловлено |
Из них возвращалось |
|
|
Самцы Ad1 20 4 (8) 12 1 (2) Ad2 новые 20 8 (8) 5 0 Ad2 кольц. пришлые 2 0 2 2 (2) Всего 42 12 (16 ) 19 3 (4) Самки Ad1 29 5 (8) 11 1 (1) Ad2 новые 16 3 (5) 8 1 (1) Ad2 кольц. пришлые 1 0 3 1 (1) Всего 46 8 (13 ) 22 3 (3) |
||||
Таблица 4. Расстояние между гнёздами окольцованных белобровиков Turdus iliacus в смежные годы
|
Расстояние между гнездами в смежные годы, м |
Число птиц данного возраста при первом отлове |
|||||
|
Первогодки |
Старше 1 года |
|||||
|
Самцы |
Самки |
Всего |
Самцы |
Самки |
Всего |
|
|
0–50 |
7 |
1 |
8 |
7 |
1 |
8 |
|
51–100 |
6 |
2 |
8 |
11 |
8 |
19 |
|
101–200 |
3 |
5 |
8 |
12 |
3 |
15 |
|
201–300 |
2 |
4 |
6 |
6 |
3 |
9 |
|
301–500 |
6 |
5 |
11 |
2 |
1 |
3 |
|
501–1000 |
2 |
3 |
5 |
2 |
1 |
3 |
|
Более 1000 |
1 |
1 |
2 |
1 |
– |
1 |
|
Всего |
27 |
21 |
48 |
41 |
17 |
58 |
Однако при крайне низкой плотности гнездового населения белобровика на стационаре «Маячино» и склонности этого вида к парцеллярному распределению, наблюдаемые различия вряд ли можно связать с конкурентными отношениями особей. Более вероятно, что первогодки заселяют участки с недостаточными кормовыми и защитными условиями из-за отсутствия опыта. Ориентируясь в основном по весенней ситуации, они оседают на рано вытаявших прогалинах на вырубках, болотах, лесных полянах, и только после вылупления птенцов обнаруживают, что летом такие стации бедны их основным кормом – дождевыми червями. На такие участки птицы на следующий год уже не возвращаются, из-за чего оказываются на большем расстоянии от места гнездования в предыдущем году (первого в своей жизни), чем птицы более старших возрастов (табл. 4); различия статистически значимы ( χ2 = 11.21, df = 4, P < 0.05).
Перемещение молодых белобровиков на лучшие участки после первого гнездового сезона приводит к снижению доли вернувшихся птиц и особей старших возрастов в гнездовом населении субоптимальных местообитаний. В результате, величина популяционных показателей, полученных по результатам изучения локального населения соседних участков, может заметно отличаться и друг от друга, и от средних значений, характеризующих данную популяцию. Исходя из этого, при расчётах основных популяционных параметров, зависящих от этих характеристик, целесообразно, наряду с уровнем контроля и величиной контролируемой площади (Соколов 1991; Паевский 2008), по возможности, принимать во внимание и степень её привлекательности для данного вида.
Авторы выражают свою искреннюю признательность В.Б.Зимину, А.В.Артемьеву, Н.В.Лапшину, Л.С.Захаровой, А.В.Сухову, всем коллегам, аспирантам и студентам, принимавшим участие в сборе материалов в течение столь длительного периода.
