Динамика ферментативной активности и пигментного состава в тканях водного погруженного растения Ceratophyllum demersum в условиях воздействия ксенобиотиков и последующей реабилитации

Автор: Макурина Ольга Николаевна, Розина Светлана Алексеевна, Розенцвет Ольга Анатольевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 4-5 т.17, 2015 года.
Бесплатный доступ
В статье рассматривается влияние стрессовых концентраций ионов свинца, катионных синтетических поверхностно-активных веществ (СПАВ) и их сочетания на ферментативную активность, пигментный состав и содержание водорастворимых белков в тканях водного погруженного растения Ceratophyllum demersum.
Водные растения, тяжелые металлы (тм), поверхностно-активные вещества (пав), каталаза, пероксидаза, водорастворимые белки, хлорофилл а, хлорофилл b, каротиноиды, окислительный стресс, доктор биологических наук, профессор кафедры биохимии
Короткий адрес: https://sciup.org/148203963
IDR: 148203963 | УДК: 582.671:581.5.
Dinamics enzimatic activity and pigment contein it tissues of aquatic macrophyt Ceratophyllum demersum under the impact of xenobiotics and subsequent rehabilitation
In this paper the effects of ion of lead, cationic surfactants and it’s combination on enzymatic activity, pigment fan protein contain in water submerged plant Ceratophyllum demersum are considered.
Текст научной статьи Динамика ферментативной активности и пигментного состава в тканях водного погруженного растения Ceratophyllum demersum в условиях воздействия ксенобиотиков и последующей реабилитации
В последние два десятилетия значительно возрос интерес к экологическим аспектам загрязнения водных объектов синтетическими поверхностно-активными веществами (СПАВ), получаемыми из углеводородов нефти. Это обусловлено, с одной стороны, возрастающими масштабами производства и объемами использования этих соединений в различных отраслях промышленности и сельского хозяйства, в том числе в производстве синтетических моющих средств (СМС), а с другой – чрезвычайной широким диапазоном отрицательного влияния СПАВ как на водные экосистемы, так и на организм человека, а также их устойчивостью к биодеградации [31]. Катионные СПАВ наиболее устойчивы к биодеградации и представляют большую опасность для живых организмов по сравнению с анионными и амфотерными СПАВ [19,35].
Активные формы кислорода (АФК), образующиеся при поступлении поллютантов в организм, способны повреждать нативную структуру клеточных мембран и инициировать процессы перекисного окисления липидов (ПОЛ), что может привести к развитию окислительного стресса. Вместе с тем, в клетках существуют антиоксидантные системы, включающие ферменты (каталазу, пероксидазу, супероксиддисмутазу и др.) и низкомолекулярные соединения, которые обеспечивают защиту живых организмов от АФК [20, 27, 30].
Фотосинтетические пигменты (хлорофиллы и каротиноиды) являются основными компонентами фотосинтетического аппарата зеленых растений, и изменение их содержания служит чувствительным маркером нарушений метаболизма растительной клетки в целом [5]. ТМ нарушают водный статус и газообмен, снижают содержание пигментов и инактивируют ключевые ферменты метаболических путей. СПАВ вызывают солюбилизацию мембран, нарушая работу пигментного аппарата. Количественное содержание пигментов и их соотношение в тканях растений являются фактором, определяющим физиологическое состояние растения, отражают изменения роста и развития при различных стрессах [5, 22].
Содержание водорастворимых белков в растительных тканях изменяется под действием огромного количества стрессоров (в том числе поллютантов) и служит чувствительным маркером изменений клеточного метаболизма [33].
В отечественных работах изучены биологические эффекты различных видов СПАВ на ряд высших наземных и водных растений [12, 13], приведены механизмы действия и выдвинута идея самоочистки сточных вод с помощью водных макрофитов [9]. В работах Мурзина и соавт. [11] изучено действие катионных и анионных СПАВ по отдельности и в сочетании с ионами меди и кадмия на биохимические показатели высшего водного растения Egeria densa . Согласно исследованиям наших соотечественников [11, 13], наиболее эффективная концентрация ионов свинца – 100 мкМоль/л, катионных СПАВ – 1%. Период воздействия токсиканта составил 12 и 72 часа. Реабилитация проводилась в течение пяти суток. Объектом исследования был выбран пресноводный макрофит с широким ареалом обитания Ceratophyllum demersum, фактор воздействия –ионы свинца (100 мкМоль/л ионов свинца в растворе ацетата свинца), катионные СПАВ (1% раствор широкодоступного катионного СПАВ – ополаскиватель для белья «Дося» и их сочетание в указанных концентрациях.
Целью нашей работы явилось исследование динамики каталазной, пероксидазной активности, пигментного состава и содержания водорастворимых белков в тканях водного погруженного растения C. demersum при воздействии ионов ТМ (на примере свинца), катионных СПАВ (на примере ополаскивателя для белья «Dosia») и их сочетания, а также в период реабилитации, после удаления поллютантов из воды.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом исследования был выбран пресноводный макрофит роголистник погружённый ( Ceratophyllum demersum L.) [7].
Эксперимент проводился в лабораторных условиях при одинаковой интенсивности и регулярности светового потока, а также при постоянной температуре (20°С). Для этого в опыте была использована комбинация люминесцентных ламп и установлен постоянный период освещения, равный 18 ч.
В ходе эксперимента растения были разделены на 4 группы, различающиеся средой выращивания. Контрольная группа растений находилась в среде отфильтрованной водопроводной воды, первая опытная группа инкубировалась в присутствии 100 мкМоль/л ионов свинца в растворе ацетата свинца, вторая – в присутствии 1% катионного СПАВ, третья – в присутствии обоих поллютантов в указанных концентрациях. Непосредственно перед началом исследований фрагменты растений длиной до 50 мм, считая от точки роста, помещали в стеклянные емкости объемом 1 дм3.
Продолжительность воздействия поллютанта составила 12 и 72 часа. По истечении указанного периода экспозиции часть растений из каждой группы отбирали на исследования, а часть переносили в чистую отфильтрованную воду для реабилитации. Длительность реабилитации составила 5 суток: для 12 часов инкубации – это 132 час, для 72 часов инкубации – 192 час эксперимента. После реабилитации также проводили измерения биохимических показателей.
Методы исследования . Исследование уровня каталазной активности проводили по методу по Королюк М.А. и Ивановой Л.И. [8]. Определение пероксидазной активности осуществляли общепринятым методом по Бояркину А.М. (метод основан на определении скорости реакции окисления бензидина до образования синего продукта окисления определенной концентрации) [2]. Определяли содержание водорастворимых белков по методу М. Брэдфорд [21] и выражали ферментативную активность в удельных единицах на г белка. В растительных тканях исследовали содержание фотосинтетических пигментов по методу Брагинского Л.П. (количественное определение пигментов основано на определении оптической плотности пигментов на длинах волн, являющихся максимумами их поглощения) [1]. Статистическую обработку данных (среднее значение, стандартное отклонение) проводили с использованием стандартных статистических методов и компьютерных программ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Инкубация в среде ионов свинца приводила к развитию хлороза листьев C. demersum , эффект усиливался по мере увеличения срока инкубации. Токсическое действие 1% раствора катионных СПАВ вызвало опад листьев после 12 часов воздействия и фрагментацию растения после 72 часов. Эффекты сочетания ксенобиотиков стали причиной хлороза и частичного опада листьев. Реабилитация от воздействия ионов свинца и сочетания поллютантов прошла более менее успешно, после инкубации в среде катионных СПАВ растение не восстановилось.
Динамика каталазной активности представлена на рис. 1а. Токсическое действие 100 мкМоль/л ионов свинца вызвало снижение уровня каталазной активности в два раза на 12 и 72 часы инкубации, влияние 1% раствора катионных СПАВ привело к повышению ферментной актив-
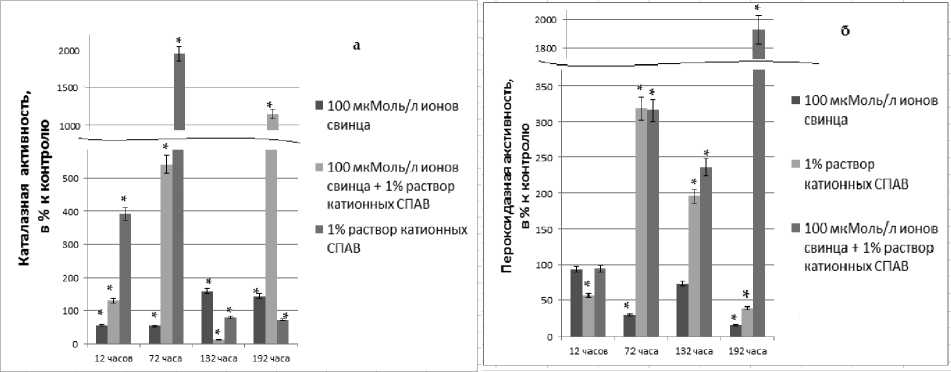
Рис. 1. Динамика каталазной (а) и пероксидазной (б) активности в тканях С. demersum ;
контроль принят за 100%; * - степень достоверности р< 0,01
ности в 4 раза к 12 часу инкубации и в 19 раз к 72 часу инкубации, а сочетание ксенобиотиков повысило каталазную активность в 1,3 и 5,4 раза соответственно.
Для СПАВ характерно образование плёнки на границе раздела сред: воздух-вода, вода-ткани растения. В эксперименте с катионными СПАВ и их сочетанием с ионами свинца наблюдалось образование плёнки на поверхности сосуда и на организме растения, из-за чего оно погрузилось на дно сосуда. Вероятно, плёнка катионных СПАВ препятствовала нормальному дыханию тканей С. demersum, поэтому индуктивно повышалось содержание молекул каталазы в клетках и, соответственно, ферментативная активность повышалась в разы.
Реабилитация от токсических эффектов ионов свинца показала, что каталазная активность превысила контрольные значения в 1,4 - 1,5 раза, что может быть связано процессами ликвидации последствий окислительного стресса, вызванного ионами тяжёлого металла. Реабилитация от действия катионных СПАВ привела к снижению ферментативной активности на 21 - 27%, что является следствием истощения пула фермента и ресурсов клетки после усиленного синтеза каталазы в период инкубации. Интересно отметить, что реабилитация от 12 часов сочетанного воздействия ксенобиотиков привела к снижению каталазной активности в семь раз, а реабилитация от 72 часов инкубации вызвала повышение ферментативной активности в 11,5 раз. Возможно, поллютанты взаимодействуют между собой, характеристика взаимодействия меняется с течением времени, усиливается окислительный стресс и образование активных форм кислорода (АФК), поэтому реабилитация от 72 часов инкубации в среде сочетания поллютантов является результатом дальнейшей борьбы организмы С. demersum с токсическими эффектами.
Динамика пероксидазной активности представлена на рис. 1б.
Через 12 часов инкубации 100 мкМоль/л ионов свинца и сочетание ксенобиотиков не вызвало достоверного изменения пероксидазной активности. Токсическое действие 1% раствора катионных СПАВ после 12 часов воздействия привело к снижению пероксидазной активности в 2 раза. К 72 часу инкубации ферментативная активность резко изменилась: в среде с добавлением ионов свинца она снизилась в 3 раза, а в средах с добавлением катионных СПАВ и сочетанием ксенобиотиков - повысилась в 3,2 раза.
Из литературных данных известно, что двухвалентные ионы ТМ в высоких концентрациях способны частично или полностью вытеснять металлы из активного центра ферментов, в результате чего теряется их активность [14, 32]. На наш взгляд, по-видимому происходило вытеснение кальция из молекул фермента ионами свинца, что и приводило к значительному ингибированию пероксидазной активности в тканях С. demersum в условиях инкубации в среде с добавлением 100 мкМоль/л ионов свинца. Токсический эффект СПАВ на пероксидазную активность может проявляться по-разному. В исследованиях Давлетшина А.И. с сотр. [3,4] показано, что СПАВ способны как вызывать активацию фермента пероксидазы, так и приводить к ингибированию его активности, воздействуя как на молекулы фермента непосредственно, так и на его мембранное окружение, либо на состояние субстрата, что и наблюдалось в условиях инкубации: после 12 часов пероксидазная активность понизилась в два раза, а к 72 часу - повысилась в 3,2 раз. Эффекты сочетания ксенобиотиков повторяют эффекты катионных СПАВ.
Снижение пероксидазной активности в период реабилитации от действия ионов свинца (на 27% от 12 часов инкубации и 85% от 72 часов инкубации) соотносится с повышением каталазной активности. Ионы свинца привели к повреждениям в клетке и развитию окислительного стресса, активно ликвидируемым ферментом каталазой, в результате чего развивается субстратное ингибирование пероксидазы. Реабилитация от 12 часов воздействия 1% раствора катионных СПАВ привела к повышению пероксидазной активности в 2 раза, реабилитация от 72 часов – к снижению ферментативной активности в 2,5 раз. Столь значительное снижение пероксидазной активности может быть следствием истощения пула фермента после инкубации. Реабилитация от сочетанного воздействия ксенобиотиков привела к повышению пероксидазной активности в 2,4 раза (от 12 часов) и 19 раз (от 72 часов), что, как и в случае с каталазой, свидетельствует о дальнейшем влиянии токсикантов на организм C. demersum.
Изменение содержания водорастворимых белков на протяжении всего эксперимента представлено на рис. 2. После 12 часов инкубации в среде поллютантов только действие ионов свинца привело к достоверному снижению в 1,7 раза содержания водорастворимых белков. Инкубация в течение 72 часов в среде 100 мкМоль/л ионов свинца вызвала снижение содержания водорастворимых белков в 1,4 раза, в среде с добавлением 1% раствора катионных СПАВ – в 6,7 раз, в среде с сочетанием ксенобиотиков – в 4,3 раза. Подобная тенденция могла быть обусловлена инактивацией или даже денатурацией белков этими ксенобиотиками [34]. Известно, что СПАВ способны также повреждать молекулы нуклеиновых кислот [6], и мы предполагаем, что поступление катионных СПАВ в организм индуцировало структурные изменения в молекулах ДНК, которые влекли за собой нарушение процессов белкового синтеза.
Из литературных данных известно, что в присутствии СПАВ возможно увеличение поступления ТМ в ткани растений [25]. По-видимому, подобный механизм имел место в проведенной нами работе. В этих условиях предположительно более интенсивно протекали процессы повреждения белковых молекул и ингибирования их биосинтеза. В условиях повышенного поступления ТМ в клетку, и такой эффект известен из литературы [28], могли также усиливаться процессы протеолиза, приводящие к деградации белков.
Реабилитация от 12 часового воздействия ксенобиотиков показала снижение содержания водорастворимых белков на 21 – 40 %, тогда как в результате реабилитации от 72 часов инкубации от эффектов ионов свинца и катионных СПАВ содержание водорастворимых белков достоверно не отличалось от контрольных величин. Реабилитация от сочетания ксенобиотиков показала снижение содержания водорастворимых белков в 2,6 раза, что свидетельствовало о сложной и длительной адаптации растений к токсическому действию смеси поллютантов.
Динамика содержания хлорофилла а представлена на рис. 3а. Наибольшие отклонения в содержании данного пигмента вызвало воздействие 1% раствора катионных СПАВ. К 12 часам содержание хлорофилла а превысило контрольные значения в 2,2 раза и стало снижаться в последующие часы инкубации. Через 72 часа воздействия катионных СПАВ содержание хлорофилла а превышало контрольные значения всего на 16%, а в период реабилитации снизилось и составило 34% от контроля. Ионы свин-
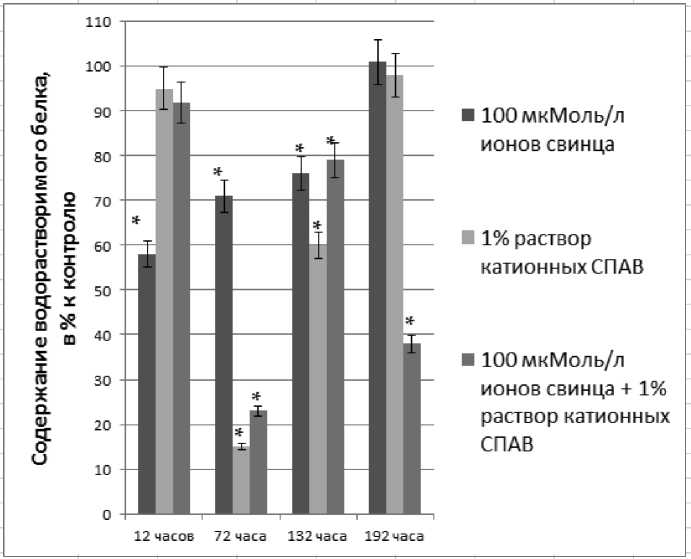
Рис. 2. Динамика содержания водорастворимых белков в тканях C. demersum; контроль принят за 100%; * - степень достоверности р< 0,01
ца и сочетание ксенобиотиков через 12 часов воздействия привели к снижению содержания хлорофилла а в 1,7 и 2 раза соответственно. Однако на 72 часу инкубации в присутствии 100 мкМоль/л ионов свинца содержание хлорофилла а возросло и превысило контрольные показатели в 1,7 раз. В период реабилитации содержание хлорофилла а сначала сравнялось со контрольными значениями, а потом, к 192 часу эксперимента, понизилось в 3 раза. Реабилитация от сочетанного действия поллютантов показала, что содержание хлорофилла а превысило контрольные значения на 15 – 19%.
Содержание хлорофилла b на протяжении всего эксперимента представлено на рис. 3б. Токсическое действие поллютантов привело к снижению содержания хлорофилла b в 1,5 – 3 раза на 12 час инкубации, однако на 72 час содержание пигмента повысилось в 2,4 раз (100 мкМоль/л ионов свинца) и в 1,6 раза (1% катионных СПАВ). Эффект сочетанного действия ксенобиотиков снизил содержание хлорофилла b в 2 раза на 72 час инкубации. Реабилитация показала резкое изменение содержание пигмента в тканях C. demersum . Хотя на 132 час эксперимента содержание хлорофилла b после воздействия ионов свинца и сочетания поллютантов достоверно не отличалось от контрольных величин, на 192 час эксперимента содержание хлорофилла b после воздействия ионов свинца составило всего 2,5 % от контроля, а после сочетанного воздействия – превысило контроль в 20 раз. В период реабилитации от токсических эффектов катионных СПАВ содержание хлорофилла b снизилось в 3,3 раза на 132 час эксперимента и в 7,7 раз на 192 час.
Токсическое действие поллютантов привело к снижению содержания хлорофилла b в 1,5 – 3 раза на 12 час инкубации, однако на 72 час содержание пигмента повысилось в 2,4 раз (100 мкМоль/л ионов свинца) и в 1,6 раза (1% катионных СПАВ).
Эффект сочетанного действия ксенобиотиков снизил содержание хлорофилла b в 2 раза на 72 час инкубации. Реабилитация показала резкое изменение содержание пигмента в тканях C. demersum . Хотя на 132 час эксперимента содержание хлорофилла b после воздействия ионов свинца и сочетания поллютантов достоверно не отличалось от контрольных величин, на 192 час эксперимента содержание хлорофилла b после воздействия ионов свинца составило всего 2,5 % от контроля, а после сочетанного воздействия – превысило контроль в 20 раз. В период реабилитации от токсических эффектов катионных СПАВ содержание хлорофилла b снизилось в 3,3 раза на 132 час эксперимента и в 7,7 раз на 192 час.
Динамика содержания каротиноидов представлена на рис. 4. Токсические эффекты ионов свинца привели к повышению содержания каротиноидов в 3,8 раз на 12 час инкубации и к снижению на 40% на 72 час эксперимента. Реабилитация показала увеличение содержания пигментов на 132 час в 2,2 раза и на 192 час – в 1,3 раза.
Раствор катионных СПАВ на двенадцатый час инкубации также вызвали повышение содержания каротионидов в 2,4 раз, а в последующие часы реабилитации их содержание превышало контроль на 11 – 24%. Хотя сочетанное действие ксенобиотиков и вызвало снижение содержания каротиноидов на 40% на 12 час и увеличение в 4,7 раза на 72 инкубации, после периода реабилитации опытные показатели достоверно не отличались от контрольных.
Исходя из анализа содержания хлорофиллов a и b в растении в период воздействия катионных СПАВ, можно предположить, что многократное увеличение содержания хлорофиллов служило ответом клетки на возросшие потребности в дыхании (т.к. катионные СПАВ образовывали пленку на поверхности стеблей и листьев, препятствуя проникновению кислорода внутрь), однако
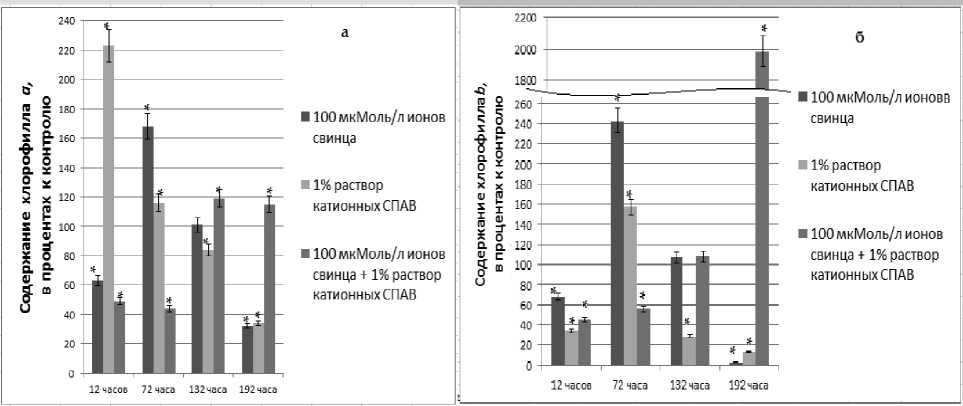
Рис. 3. Динамика содержания хлорофилла а (а) и хлорофилла b (б) в тканях C. demersum в процентах к контролю, контроль принят за 100%; * – степень достоверности р< 0,01
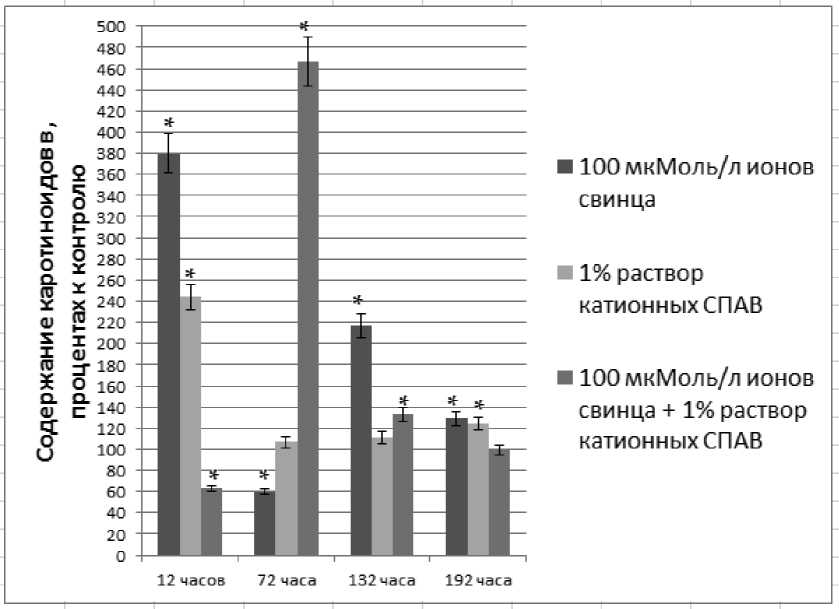
Рис. 4. Динамика содержания каротиноидов в тканях C. demersum в процентах к контролю, контроль принят за 100%; * - степень достоверности р< 0,01
в метаболизме уже возникали необратимые нарушения, приводящие к последующей деградации пигментов в период последействия. Снижение содержания хлорофиллов a и b при воздействии ионов свинца, вероятно, связано, с повреждением ионами ТМ тиллакоидных мембран и нарушением работы пигментного аппарата.
Из литературных данных известно, что в фотосинтетическом аппарате каротиноиды выполняют функцию антиоксидантов, обладающих способностью замедлять фотохимические реакции, приводящие к образованию свободных радикалов, повреждению мембран хлоропластов и деградации хлорофиллов [23]. По-видимому, увеличение их содержания в растении при воздействии поллютанта и в период реабилитации было вызвано необходимостью защиты хлорофиллов в условиях стресса.
Во время реабилитации фитотоксическое действие поллютантов сохранялось и предположительно приводило к истощению энергетических ресурсов клетки, что затрудняло поддержание нормальной жизнедеятельности растения. На наш взгляд, и это также подтверждается данными литературных источников [24, 26, 29, 36], повреждение пигментного комплекса в растительных тканях было обусловлено солюбилизацией тил-лакоидных мембран, изменением их жирно-кислотного состава, нарушением ультраструктуры хлоропластов, а также солюбилизацией хлоро-филл-белковых комплексов под действием СПАВ. После процессов реабилитации от токсических эффектов ионов свинца и катионных СПАВ в тканях C. demersum сохранялось низкое содержание хлорофиллов a и b, а содержание каротиноидов лишь на 11 – 24% превысило нормальные значения, что свидетельствует о процессах ликвидации окислительного стресса.
Реабилитация от сочетанного действие ксенобиотиков показала, что все показатели, кроме содержания хлорофилла b , вернулись к контрольным значениям. Вероятно, этот пигмент взял на себя основную работу по выполнению защитных и дыхательных функций.
Токсические эффекты ионов свинца и катионных СПАВ оказывают разнонаправленное действие на ферментативную активность и содержание пигментов и водорастворимых белков в тканях C. demersum. Ионы ТМ замещают ионы металлов в активных центрах ферментов, нарушают работу пигментного аппарата, вызывая хлороз. Основной токсический эффект катионных СПАВ – солюбилизация мембран, выход ферментов в клетку, нарушение внутриклеточных взаимосвязей, реабилитация от 1% катионных СПАВ крайне затруднительна для тканей C. demersum.
Эффекты сочетанного воздействия ксенобиотиков развивают и в реабилитационных период. Преобладают токсические эффекты катионных СПАВ, однако поллютанты взаимодействуют между собой, смягчая повреждения мембранных структур. Для более полной реабилитации от сочетанного воздействия требуется, вероятно, более 5 суток.
Список литературы Динамика ферментативной активности и пигментного состава в тканях водного погруженного растения Ceratophyllum demersum в условиях воздействия ксенобиотиков и последующей реабилитации
- Бриттон Г. Биохимия природных пигментов. Пер. с англ. М.: Мир, 1986. 422 с.
- Бояркин А.Н. Быстрый метод определения активности пероксидазы//Биохимия. 1951. Т. 16. Вып. 4. С. 352-355.
- Давлетшин А.И., Калабина Н.А., Зайцев С.Ю., Егоров В.В. Влияние поверхностно-активных веществ на активность пероксидазы. Влияние катионных ПАВ//Биоорган. химия. 1998. Т. 24. № 6. С. 430 -432.
- Давлетшин А.И., Сильвестрова И.Г., Зубов В.П, Егоров В.В. Влияние ПАВ различной природы на активность пероксидазы и трипсина//Вестн. Моск. ун-та. Сер. 2. Химия. 1998. Т. 39. № 4. С. 272 -275.
- Головко Т.К., Далькэ, И.В., Бачаров, Д.С. Мезоструктура и активность фотосинтетического аппарата трех видов растений сем. Crassulaceae в холодном климате//Физиология растений. 2008. Т. 55, № 5. С. 671-680.
- Гостюхина О.Л., Солдатов А.А. и др. Влияние тетрадецилтриметиламмоний бромида на состояние ферментной системы антиоксидантной защиты тканей черноморского моллюска Mytilus galloprovincialis Lam.//Доп. НАН України. 2007. № 11. С. 147-151.
- Жизнь растений. В 6-ти т. Т. 5. Ч. 1. Цветковые растения . М.: Просвещение, 1980. С. 188-190.
- Королюк М.А., Иванова Л.И., Майорова И.Г., Токарев В.Е. Метод определения активности каталазы//Лаб. дело. 1988. № 1. С. 16-19.
- Лазарева Е.В., Остроумов С.А. Ускорение снижения концентрации поверхностно-активного вещества в воде микросома в присутствии растений: инновации для фитотехнологии. ДАН, 2009. Т. 425. №6. С. 843 -845.
- Маслова Т.Г., Мамушина, Н.С., Шерстнева О.А., Буболо Л.С., Зубкова Е.К. Структурно-функциональные изменения фотосинтетического аппарата у зимневегетирующих хвойных растений в различные сезоны года//Физиология растений. 2009. Т. 56, № 5. C. 672-681.
- Мурзин И.Р., Макурина О.Н., Косицына А.А., Розенцвет О.А. Особенности действия загрязнителей различной химической природы на содержание водорастворимых белков в тканях водного погруженного растения Egeria Densa//Вестник СамГУ Естественнонаучная серия, 2010. Т. №4 (78). С. 191-199.
- Остроумов С.А. Биологические эффекты при воздействии поверхностно-активных веществ на организмы. М.: МАКС-пресс, 2001. 334 с.
- Остроумов С.А. О биотическом самоочищении водных экосистем. Элементы теории//ДАН, 2004. Т. 396. №1. С. 136 -141.
- Прохорова Н.В., Матвеев Н.М., Павловский В.А. Аккумуляция тяжелых металлов дикорастущими и культурными растениями в лесостепном и степном Поволжье. Самара: Изд-во Самар. ун-та, 1998. 131 с.
- Стайлс В. Микроэлементы в жизни растений и животных. М.: Наука, 1949. 290 с.
- Титов А.Ф., Таланова В.В., Казнина Н.М., Лайдинен Г.Ф. Устойчивость растений к тяжёлым металлам. Петрозаводск: Карельский научный центр РАН, 2007. 172 с.
- Трахтенберг И.М. Тяжелые металлы во внешней среде: Современные гигиенические и токсикологические аспекты. Минск: Наука и техника, 1994. 286 с.
- Филенко О.Ф., Михеева И.В. Основы водной токсикологии. М.: Колос, 2007. 140 с.
- Холмберг К., Йенссон Б., Кронберг Б., Линдман Б. Поверхностно-активные вещества и полимеры в водных растворах. М.: БИНОМ. Лаборатория знаний, 2007. 528 с.
- Bhattacharjee S. Reactive oxygen species and oxidative burst: Roles in stress, senescence and signal transduction in plants//Curr. Sci. 2005. Vol. 89. No. 7. P. 1113 -1121.
- Bradford M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding//Anal. Biochem. 1976. Vol. 72. P. 248-254.
- Dubey R.S. Metal toxicity, oxidative stress and antioxidative defense system in plants/Ed. by S. Dutta Gupta. Reactive oxygen species and antioxidants in higher plants. Enfield: Science Publishers, 2010. P. 177 -203.
- Knox J.P., Dodge, A.D. Singlet oxygen and plants//Phytochemistry. 1985. Vol. 24. P. 889-896.
- Kobayashi I., Kunoh H. et al. Effects of several nonionic and anionic surfactants on cucumber protoplasts//J. Pesticide Sci. 1990. Vol. 15. P. 71-80.
- Maresova J., Hornik M. et al. Influence of anionic surfactants on Zn2+ and Sr2+ uptake by ivy (Hedera helix L.) leaves//Nova Biotechnol. 2009. Vol. 9. No. 3. P. 333-339.
- Markwell J.P., Thornber J.P. Treatment of the thylakoid membrane with surfactants//Plant Physiol. 1982. Vol. 70. P. 633-636.
- Mohd M., Taqi A.K., Firo M. Role of nitric oxide in regulation of H2O2 mediating tolerance of plants to abiotic stress: A synergistic signaling approach//J. Stress Physiol. Biochem. 2011. Vol. 7. No. 2. P. 34 -74.
- Palma J.M., Sandalio L.M. et al. Plant proteases, protein degradation and oxidative stress: Role of peroxisomes//Plant Physiol. Biochem. 2002. Vol. 40. P. 521-530.
- Rinallo C., Bennici A., Cenni E. Effects of two surfactants on Triticum durum Desf. plantlets//Env. Exp. Bot. 1988. Vol. 28. No. 4. P. 367-374.
- Rio L.A., Puppo A. Reactive oxygen species in plant signaling/Berlin: Springer, 2009. 245 p.
- Rosen M.J., Li F., Morall S.W., Versteeg D.J. The relationship between the interfacial properties of surfactants and their toxicity to aquatic organisms//Environ. Sci. Technol. 2001. Vol. 35. No. 5. P. 954-959.
- Schützendübel A., Polle A. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization//J. Exp. Bot. 2002. Vol. 53. No. 372. P. 1351 -1365.
- Singh P.K., Tewari R.K. Cadmium toxicity induced changes in plant water relations and oxidative metabolism of Brassica juncea L. plants//J. Environ. Biol. Vol. 24. No. 1. 2003. P. 107-112.
- Vincenzini M., Favilli F. et al. Detergents as selective inhibitors and inactivators of enzymes//Physiol. Chem. Phys. Med. 1985. Vol. 17. No. 3. P. 279-295.
- Ying G.G. Fate, behavior and effects of surfactants and their degradation products in the environment//Environ. Int. 2006. Vol. 32. No. 3. P. 417-431.
- Yang C. M., Hsu J.-C. et al. Pigment solubilization of the chloroplast thylakoid membranes by a surfactant//Bot. Bull. Acad. Sci. 1996. Vol. 37. P. 121-126.
