Динамика флорального органогенеза Syringa josikaea Jacq. при интродукции в условиях арктического климата г. Мурманска

Автор: Василевская Наталья Владимировна, Морозова Дарья Анатольевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 4 (165), 2017 года.
Бесплатный доступ
Представлены данные морфофизиологического анализа генеративных почек Syringa josikaea Jacq., интродуцированной в г. Мурманске. Показано, что в условиях арктического климата дифференциация осей соцветий и органогенез цветков могут происходить как за год до цветения, так и в весенний период за два - три месяца до его начала. Выявлена поливариантность флорального органогенеза сирени венгерской, произрастающей в разных районах города. Экспериментальные данные по асинхронности процессов флоральной дифференциации у сирени впервые получены в условиях Арктики.
Флоральный органогенез, арктика
Короткий адрес: https://sciup.org/14751192
IDR: 14751192 | УДК: 581.3
Dynamics of floral organogenesis of Syringa josikaea Jacq. at the time of introduction in conditions of the arctic Murmansk climate
Research data of the morpho-physiological analysis of the generative buds of Syringa josikaea Jacq., introduced in Murmansk, are presented. It is shown that in conditions of the Arctic climate the differentiation of axes of the inflorescence and organogenesis of flowers can occur both a year before flowering and in the spring period two or three months before its inception. The polyvariety of the floral organogenesis of the Hungarian lilac, growing in different districts of Murmansk has been revealed. Experimental data on the asynchronous processes of the lilac floral differentiation were first obtained in the Arctic.
Текст научной статьи Динамика флорального органогенеза Syringa josikaea Jacq. при интродукции в условиях арктического климата г. Мурманска
При интродукции растений в новые для них условия произрастания особое значение имеет изучение морфогенеза, так как биологический ритм развития во многом определяет степень их приспособленности к местному климату [11]. Известно, что сроки и темпы закладки генеративных почек варьируют в зависимости от климатических условий региона, ритмов развития растения и его зимостойкости. Причем для морозоустойчивых видов характерны ранние сроки прохождения основных фенофаз и закладки репродуктивных органов [1]. У растений, успешно интродуцируемых в северных условиях, отмечается более интенсивная дифференциация органов цветка. Этот факт подтверждается исследованиями и в отношении сирени венгерской [14].
Род Syringa L. широко распространен в различных регионах мира благодаря своей декоративности, при этом данные по числу видов противоречивы [29], [31]. Одни исследователи считают, что существует 40 видов сиреней, распространенных как в Европе, так и в Азии [31]. По другим данным, род Syringa L. включает 27 видов, из которых 25 происходят из Азии и только 2 из Европы [29], одним из европейских видов является Syringa josikaea Jack. Информация по флоральному органогенезу и цитоэмбриологии видов, сортов и культиваров сиреней в литературе достаточно ограниченна [9], [10], [24], [27], [28], [30], значительно лучше изучена фенология их цветения [4], [18], [22], [23], [25], [32]. В последние годы начались активные исследования репродуктивного развития наиболее перспективных куль
тиваров S. vulgaris [24], [28], [30], аналогичные исследования по S. josikaea немногочисленны [3], [14], [16]. Вместе с тем именно сирень венгерская успешно интродуцирована на урбанизированных территориях Евро-Арктического региона, в то время как S. vulgaris в условиях полярного дня не образует репродуктивных органов и не цветет.
Цель исследования – изучение динамики флорального органогенеза Syringa josikaea Jacq. (сирени венгерской) в условиях арктической урбанизированной территории (на примере г. Мурманска).
Исследования проводились в г. Мурманске – самом большом в мире незамерзающем порту за полярным кругом (68°58’ с. ш., 33°4’ в. д.). Мурманск растянут вдоль Кольского залива Баренцева моря на 25 км, причем отдельные районы разделены между собой сопками и участками естественной растительности. Город находится в Атлантико-Арктической зоне умеренного климата. Средняя температура января – февраля -10– 11 °C, средняя температура июля: +12–13 °C [7]. Большая часть осадков в Мурманске, примерно 500 мм/год, выпадает с июня по сентябрь. Снеговой покров держится в городе в среднем 210 дней и сходит к маю. Ветер имеет муссонный характер: зимой преобладают южные ветра, летом – северные. Полярная ночь начинается 29 ноября и заканчивается 13 января (44 дня), полярный день – с 22 мая по 22 июля (62 дня) [21]. Продолжительность вегетационного периода в Мурманске составляет примерно 120–130 дней. Основными источниками загрязнения атмосферы города являются Мурманские котельные, Мурманский морской порт, завод по термической обработке твердых бытовых отходов, автотранспорт.
МАТЕРИАЛЫ И МЕТОДЫ
Об ъект ом исследов а н и я явл яется Syringa josikaea Jacq. (сирень венгерская), представитель рода Syringa L. семейства Oleaceae Hoffmgg & Link.
Syringa josikaea – эндемичный кустарник Западных Карпат, реликтовый вид, имеющий узкий ареал и находящийся под угрозой исчезновения [29]. Несколько локальных ценопопуляций S. josikaea существуют здесь на небольших участках в долинах вдоль ручьев, часто представлены всего лишь несколькими экземплярами и разделены между собой большими расстояниями. В настоящее время в естественных условиях сирень венгерская встречается очень редко, произрастает только в Украинских Карпатах и горах Апусени на территории Румынии [26].
S. josikaеа – быстрорастущий, высоко засухоустойчивый и морозостойкий вид, к почвенным условиям сравнительно неприхотлив, высокая экологическая пластичность определила его успешную интродукцию в северных районах европейской части России [5], [7]. При интродукции является одним из самых зимостойких видов по сравнению с другими представителями рода Syringa L. [15]. Сирень венгерская хорошо переносит городские условия, устойчива к атмосферному загрязнению, задымлению и загазованности воздуха [13], [18], [20]. Высокая устойчивость к условиям городской среды, декоративность и успешность размножения ставят S. josikaеа на одно из первых мест в ассортименте кустарников для озеленения Мурманской области [8]. С целью интродукции саженцы сирени венгерской были впервые завезены в Полярно-альпийский ботанический сад в 1936 году из Ботанического сада г. Ленинграда [8], и уже в 1940 году S. josikaеа была высажена в городах Мурманск и Мончегорск. В настоящее время доля участия сирени венгерской в городских посадках Мурманска составляет 28 % [6].
Генеративные почки S. josikaеа смешанного типа [14], кроме соцветия в них закладываются 2–4 пары листьев. Для этого вида характерно формирование генеративных почек как в год, предшествующий цветению, так и во время него, что объясняет более длительное цветение по сравнению с другими видами сиреней [16].
Пробные площади в посадках S. josikaea на территории г. Мурманска заложены с севера на юг и отличаются высотой над уровнем моря, микроклиматическими условиями, уровнем антропогенной нагрузки: ПП1 – сквер у ТЦ «Мир» (Ленинский район); ПП2 – сквер на ул. Ленинградской (Октябрьский район); ПП3 – сквер у остановки Автопарк и ПП4 – сквер у остановки ул. Шевченко (Первомайский район). По результатам многолетних наблюдений установлено, что во всех районах города самый теплый месяц – июль, холодные – январь и февраль [21]. В северном (Ленинский) и южном (Первомайский) районах зима холоднее, чем в центральном (Октябрьский), который расположен ближе к Кольскому заливу Баренцева моря. Заморозки в черте города в августе не наблюдаются, а на южной окраине возможны [21]. В г. Мурманске только одна метеостанция, поэтому данных по климатическим различиям районов города нет.
Все экспериментальные площадки заложены в районах с повышенной техногенной нагрузкой: ПП1 – сквер у ТЦ «Мир» расположен недалеко от промышленной зоны; ПП2 – сквер на ул. Ленинградской находится рядом с железной дорогой и в непосредственной близости с Мурманским морским портом, где происходит перевалка и дробление угля открытым способом; ПП3 – сквер у остановки Автопарк и ПП4 – сквер у остановки ул. Шевченко расположены в зонах интенсивного движения автотранспорта. В качестве контрольной площадки выбраны посадки сирени около Музея военно-воздушных сил Северного флота в поселке Сафоново, в 19 км на север от г. Мурманска. Сквер расположен на берегу Кольского залива, вдали от автомобильных дорог и промышленных производств.
На каждой экспериментальной площадке маркировали по 10 кустов S. josikaea. Для изучения динамики флорального органогенеза сирени венгерской в декабре 2014 года и мае 2015 года (до начала распускания почек) на каждой пробной площади и в контроле формировали общую пробу. Сбор верхушечных почек побегов проводился с 10 кустов сирени (по 5 почек с каждого растения), общий объем пробы N = 50. Материал фиксировали в 50 % этиловом спирте. В ходе морфофизиологического анализа изучали следующие показатели: длина зачаточного соцветия, число зачатков цветков на различных этапах органогенеза, общее число зачатков цветков в соцветии. Выделялись следующие этапы органогенеза: почка в вегетативном состоянии; формирование конуса нарастания соцветия; появление зачатков осей соцветия I и II порядка; формирование цветковых бугорков; появление чашелистиков; образование лепестков; появление тычинок; смыкание лепестков и образование зачатков плодолистиков; смыкание чашелистиков и сформированный цветок [16]. Длина зачаточных соцветий измерялась с помощью окулярмикро-метра.
Исследования флорального органогенеза S. josikaеа проводили с помощью светового стереомикроскопа SM XX Carl Zeiss Jena при увеличениях (2,5 × 25; 4 × 25). В каждой пробе исследовано по 30 генеративных почек (N = 30). Данные обрабатывались методом однофакторного дис- персионного анализа, результаты считались достоверными при p ≤ 0,05. Микрофотографии выполнены с помощью малоформатной CCD-камеры, которую устанавливали на окулярную трубку.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Исследования показали, что органогенез цветка S. josikaea начинается с образования фло-ральной меристемой цветкового бугорка, затем формируются зачатки четырех чашелистиков, занимающих ортогональное положение (рис. 1). В процессе дифференцировки венчик появляется как округлая удлиненная структура, по внешним краям которой образуются зачатки 4 лепестков.
Рис. 1. Появление чашелистиков Syringa josikaea Jacq. (слева – декабрь 2014 года; справа – май 2015 года, увеличение 10 × 4)
Образование андроцея начинается с заложения двух примордиев тычинок (рис. 2), они еще не дифференцированы на тычиночные нити и пыльники, в продольном сечении представляют собой цилиндрические структуры и состоят из меристематических клеток. Двукарпельный гинецей формируется последним в ходе органогенеза цветка, расположен в медиальном положении.
Рис. 2. Появление тычинок Syringa josikaea Jacq. (слева – декабрь 2014 года; справа – май 2015 года, увеличение 10 × 4)
В результате морфофизиологических исследований декабрьских образцов генеративных почек S. josikaеа, произрастающей в разных районах Мурманска, выявлено наличие эмбриональных соцветий, различающихся по уровню флораль-ного органогенеза (рис. 3, табл. 1). По-видимому, это соцветия, заложившиеся летом и прошедшие дифференциацию осенью до вхождения в состо- яние зимнего покоя [14], что характерно для сиреней [28].
Исследования показали, что в образцах зимних почек S. josikaea из скверов на ул. Ленинградской (ПП 2 ) и ул. Шевченко (ПП 4 ), так же как и в контроле, уже осенью сформировано большое число зачатков цветков (см. рис. 3, табл. 1), находящихся на разных этапах органогенеза.
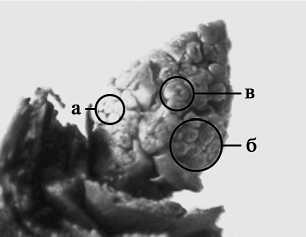
Рис. 3. Соцветие Syringa josikaea Jacq. с зачатками цветков на разных этапах органогенеза (декабрь 2014 года): а – появление чашелистиков; б – появление тычинок; в – смыкание чашелистиков, сформированный цветок
В этих пробах высока доля цветков, у которых сформированы зачатки чашелистиков (см. рис. 1), бугорки тычинок и плодолистиков (см. рис. 2), встречаются уже сформированные цветки (см. табл. 1). Этап появления чашелистиков отмечен у 30 % цветков в образцах почек с ул. Ленинградской, 23 % с ул. Шевченко, что выше, чем в контроле (18 %). На ул. Шевченко (ПП 4 ) зачатки цветков, имеющие бугорки тычинок, составляют 39 %, на ул. Ленинградской (ПП 2 ) – 16,4 %. Наиболее развиты соцветия сирени в центре города (ПП2), в них 26 % составляют сформированные цветки, в контроле – 27 %.
В генеративных почках S. josikaеа , собранных в декабре в скверах у ТЦ «Мир» (ПП1) и остановки Автопарк (ПП3), эмбриональные соцветия значительно отстают по уровню дифференциации от контрольных. Оси соцветий у сиреней на этих экспериментальных площадках развиты слабо, цветки находятся на ранних этапах органогенеза, представлены в основном цветковыми бугорками, зачатками чашелистиков и лепестков (см. табл. 1). В образцах площадки ТЦ «Мир» у 33 % цветков сформированы зачатки чашелистиков, у Автопарка – 40 %. В пробах не выявлено сформированных цветков. Среднее количество зачатков цветков в соцветиях сирени в образцах из сквера у ТЦ «Мир» в пять раз, в сквере у Автопарка – в три раза меньше, чем в контроле (см. табл. 1).
Морфофизиологические исследования весенних генеративных почек S. josikaеа выявили неожиданный результат. В майских пробах с площадок ПП1, ПП3, где в зимний период были отмечены слабая дифференциация соцветий по сравнению с контролем и минимальное общее число зачатков цветков (на соцветие), находя-
Таблица 1
Средние показатели числа зачатков цветков в соцветии Syringa josikaea г. Мурманска на разных этапах органогенеза в декабре 2014 года (шт./соцветие)
Рис. 4. Смыкание чашелистиков, сформированный цветок Syringa josikaea Jacq. (слева – зима 2014 года, увеличение 10 × 4; справа – весна 2015 года, увеличение 10 × 2,5)
Этот факт подтверждает данные о том, что у S. josikaea , в отличие от S. vulgaris , генеративные почки формируются как в год, предшествующий цветению, так и в течение года, когда происходит цветение [16]. При этом, как показали настоящие исследования, в условиях арктического климата г. Мурманска весной, после выхода растений из состояния зимнего покоя, заложение и дифференцировка цветков S. josikaea может происходить очень интенсивно.
В ходе исследования проведены измерения длины зачаточных соцветий. Получено, что в декабрьских образцах почек длина эмбриональных соцветий варьирует в пределах 500–
2200 мкм (табл. 3) по площадкам г. Мурманска и статистически значимо отличается (t ф = 3,3 – 7,4 при t Кр = 2,76, p ≤ 0,05) от длины соцветий (1300–4000 мкм) в весенний период в образцах контроля, сквера возле ТЦ «Мир» (ПП1) и сквера возле остановки Автопарк (ПП3). Полученные данные свидетельствуют о том, что в условиях атлантико-арктического климата у S. josikaеа , интродуцированной на высокоширотной урбанизированной территории, в весенний период начинается интенсивный рост эмбриональных соцветий и они значительно увеличиваются в размерах. Кроме того, выявлены различия в размерах соцветий по районам города в зимнее время: значимо (t ф = 4,1–4,4 при t Кр = 2,76, p ≤ 0,05) отличаются показатели размеров в контроле, в сквере возле ТЦ «Мир» (ПП1) и в сквере по ул. Шевченко (ПП4). В весенний период данные различия нивелируются. Самые крупные соцветия S. josikaea зимой сформированы на площадке ул. Шевченко (ПП4), к маю максимальных размеров достигают зачаточные соцветия в пробах экспериментальной площадки Автопарк (ПП3) (табл. 3).
Цветение сирени венгерской на широте г. Мурманска начинается в первой декаде июля. В других северных городах России, но на более низких широтах, цветение данного вида отмечается во второй половине мая – середине июня [4], [18]. Цветение S. josikaea летом 2015 года началось на всех экспериментальных площадках и в контроле практически синхронно – 10–11 июля. Однако окончание цветения различалось по площадкам: раньше всего (18 июля) сирень отцвела в сквере у ТЦ «Мир», 20 июля – в сквере на ул. Ленинградской, 21 июля – в скверах у остановок ул. Шевченко (ПП4) и Автопарк (ПП3), в конт-
|
Таблица 3 Длина зачаточных соцветий Syringa josikaea в г. Мурманске (в мкм) |
|||||
|
Период |
КП |
ПП 1 |
ПП 2 |
ПП 3 |
ПП 4 |
|
Декабрь 2014 года |
1100 ± 100 |
500 ± 100 |
2100 ± 500 |
900 ± 100 |
2200 ± 200 |
|
Май 2015 года |
3000 ± 600 |
2300 ± 200 |
1300 ± 100 |
4000 ± 400 |
3100 ± 500 |
Примечание. КП – контрольная площадка в п. Сафоново; ПП1 – сквер у ТЦ «Мир»; ПП2 – сквер в районе ул. Ленинградской; ПП 3 – сквер у ост. Автопарк; ПП 4 – сквер у ост. Шевченко.
роле – 18 июля. Продолжительность цветения у ТЦ «Мир» составила 7 суток, на ПП2– 10 суток, на ПП 3 , ПП 4 – 11 суток, в контроле – 7 суток. Таким образом, выявленные различия в ходе фло-рального органогенеза сирени в разных районах г. Мурманска не сказались на начале цветения, и только отчасти – на его продолжительности.
Данные по срокам заложения соцветий и органогенезу цветков у сиреней различаются в зависимости от вида, климатической зоны и сроков цветения [14], [16], [28].
Исследования флорального органогенеза соцветий и тканевой дифференцировки органов цветка у близкого S. josikaea европейского вида – S. vulgaris на территории Восточной Европы показало, что примордии чашелистиков и лепестков появляются в середине июля, до конца октября идут процессы их тканевой дифференцировки, затем наступает зимний покой [28]. После выхода из глубокого зимнего покоя дифференциация чашелистиков и лепестков продолжается и значительно ускоряется в марте – апреле [28]. Примордии тычинок появляются у зачатков цветков S. vulgaris в начале августа. В середине августа происходит дифференциация на тычиночные нити и пыльники. После выхода из состояния покоя в начале марта начинаются активная дифференцировка спорогенной ткани и формирование материнских клеток микроспор [28], к концу апреля пыльцевые зерна полностью сформированы. Заложение гинецея у S. vulgaris происходит в середине августа. В сентябре плодолистики вытягиваются в длину и формируются меристематические клетки [28].
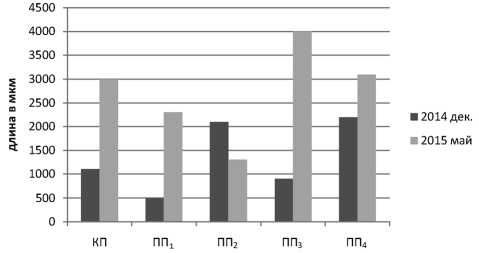
Рис. 5. Длина зачаточных соцветий Syringa josikaea в г. Мурманске (в мкм). Обозначения: КП – контрольная площадка в п. Сафоново; ПП1 – район ТЦ «Мир»; ПП2 – парк в районе ул. Ленинградской; ПП 3 – ост. Автопарк;
ПП4 – ост. Шевченко
По данным З. С. Луневой [14], у S. josikaea начало закладки соцветий происходит в конце июня, дифференциация длится в течение июля – сентября. В Иркутске этот процесс начинается в первых числах июля и заканчивается к середине сентября [16]. В г. Бишкеке [16] формирование репродуктивных почек отмечено в первой декаде июня, после окончания цветения. К концу июня в зачаточных цветках сформирован околоцветник, в первой декаде августа – все органы в виде меристематических бугорков. В зиму S. josikaea уходит с хорошо развитыми околоцветником, тычинками, зачатками плодолистиков.
ЗАКЛЮЧЕНИЕ
Предыдущими исследованиями [16] показано, что у S. josikaea , в отличие от других видов сиреней, проявляется тенденция к увеличению периода формирования репродуктивных органов в течение двух вегетационных сезонов. У части соцветий и цветков осенью уже пройдены основные этапы органогенеза, в то же время отдельные генеративные почки уходят в зиму в стадии формирования осей соцветия. Именно такой особенностью развития авторы объясняют длительное цветение данного вида. В исследованиях И. Г. Пенкиной [16] показано, что род Syringa L. можно отнести к группе древесных растений, закладывающих генеративные органы в год, предшествующий цветению. При этом автор указывает, что S. josikaea в данном случае является исключением.
Исследования, проведенные в условиях арктического климата г. Мурманска, показали, что тенденция пролонгирования периода флорально-го органогенеза еще более усиливается при интродукции S. josikaea в условиях высоких широт. Заложение зачатков соцветий и цветков у сирени венгерской на широте г. Мурманска может происходить за год до цветения, тогда в состояние зимнего покоя соцветия S. josikaea уходят с зачатками цветков разного уровня дифференциации: на ранних этапах развития – цветковые бугорки, зачатки чашелистиков, зачатки лепестков, на более поздних этапах органогенеза – с бугорками тычинок и плодолистиков, со сформированными цветками. Кроме того, дифференцировка осей соцветий и флоральный органогенез у S. josikaea могут начинаться весной, сразу после выхода из состояния зимнего покоя. При этом данный процесс происходит настолько активно, что за два – три месяца большая часть цветков в соцветии достигает поздних этапов органогенеза. Это подтверждает уже известный факт, что у видов, успешно интродуцируемых в северных широтах, отмечается более интенсивная дифференциация органов цветка [16]. По видимому, выявленная асинхронность флорального органогенеза у растений сирени разных ценопопуляций является проявлением поливариантности развития, когда в экстремальных для вида климатических условиях реализуется его генетический потенциал и проявляется высокий уровень адаптивных реакций. Известно, что S. josikaеа – реликтовый вид, в естественных условиях произрастающий в горных районах. В . Л. Комаров [12] предполагал, что Syringa L. является родом третичного периода, который в свое время был широко распространен в Старом Свете, в период повышенной геологической активности ареал рода был оттеснен на юг. Согласно М. Г. Попову [19], виды рода Syringa L. росли в пребореальных мезотермальных лесах на северной окраине арктотретичной флоры, отступающих к югу под влиянием плейстоценового ледника. S. josikaеа как вид сформировался в условиях континентального климата [17], ему свойственны высокая экологическая пластичность и устойчивость. По видимому, S. josikaеа является видом, для которого характерна широкая норма реакции, что позволяет успешно не только вегетировать, но и цвести в условиях полярного дня и низких температур на урбанизированных территориях арктического региона. Такая многовариантность развития уже выявлена у аборигенных видов растений Субарктики, когда под воздействием факторов среды, в частности температуры, в условиях высоких широт наблюдается поливариантное прохождение онтогенеза, что выражается в изменении продолжительности его этапов, разной скорости заложения органов, интенсивности ветвления и образования репродуктивных органов [2]. Для интродуцированных видов древесных растений в условиях арктической урбанизированной территории данные по поливариантности флорального органогенеза получены впервые.
В то же время результаты исследований вызывают ряд вопросов. В частности, с чем связана асинхронность в ходе репродуктивного развития S. josikaеа в разных районах Мурманска? Возможно, причиной могут быть различный возраст исследуемых насаждений, различие микроклиматических условий и т. д. Для ответа на эти вопросы необходимы дальнейшие более детальные исследования флорального органогенеза древесных интродуцентов в условиях урбанизированных территорий Евро-Арктического региона.
Research data of the morpho-physiological analysis of the generative buds of Syringa josikaea Jacq., introduced in Murmansk, are presented. It is shown that in conditions of the Arctic climate the differentiation of axes of the inflorescence and organogenesis of flowers can occur both a year before flowering and in the spring period two or three months before its inception. The polyvariety of the floral organogenesis of the Hungarian lilac, growing in different districts of Murmansk has been revealed. Experimental data on the asynchronous processes of the lilac floral differentiation were first obtained in the Arctic.
Список литературы Динамика флорального органогенеза Syringa josikaea Jacq. при интродукции в условиях арктического климата г. Мурманска
- Булыгин Н. Е. Дендрология. Л.: Агропромиздат, 1991. 352 с.
- Василевская Н. В. Поливариантность онтогенетических процессов растений в условиях высоких широт. Мурманск: МГПУ, 2007. 230 с.
- Василевская Н. В., Морозова Д. А. Тератоморфизм пыльцы Syringa josikae Jacq. при интродукции на урбанизированных территориях Российской Арктики//Ученые записки Петрозаводского государственного университета. 2016. № 8 (161). С. 7-13.
- Видякина А. А., Семенова М. В. Фенологические наблюдения за развитием вегетативных и генеративных органов Syringa Josikae Jacq. в различных районах г. Тюмени//Вестник экологии, лесоведения и ландшафтоведения. 2009. № 9. С. 142-145.
- Герасимова А. А. Фенологический мониторинг древесно-кустарниковой растительности г. Тюмени: Дис.. канд. биол. наук. Тюмень, 2015. 273 с.
- Гонтарь О. Б., Святковская Е. А., Тростенюк Н. Н., Коробейникова Н. М., Шлапак Е. П., Носатенко О. Ю. Мониторинг состояния древесных насаждений на некоторых объектах озеленения в центральной части города Мурманска//Известия Самарского научного центра Российской академии наук. 2013. № 3 (2). Т. 15. С. 621-625.
- Гонтарь О. Б., Жиров В. К., Казаков Л. А., Святковская Е. А., Тростенюк Н. Н. Зеленое строительство в городах Мурманской области. Апатиты: Изд-во КНЦ РАН, 2010. 292 с.
- Гонтарь О. Б., Святковская Е. А., Тростенюк Н. Н., Жиров В. К. Этапы создания и анализ особенностей ассортимента декоративных древесных растений для оптимального озеленения урбанизированных территорий Крайнего Севера//Вестник МГТУ 2011. Т. 14. № 3. С. 577-582.
- Жакова С. Н. Репродуктивная биология некоторых видов и культиваров рода Сирень (Syringa L.): Дис.. канд. биол. наук. Пермь, 2015. 196 с.
- Жакова С. Н., Новоселова Л. В. Эмбриологические особенности строения и развития репродуктивных структур некоторых культиваров Syringa vulgaris L.//Вестник Оренбургского государственного университета. 2015. № 9 (184). С. 101-107.
- Кирина И. Б. Особенности прохождения органогенеза у разных сортов жимолости//Достижения науки и техники АПК. 2010. № 8. С. 27-28.
- Комаров В. Л. Учение о виде у растений. М.; Л.: Изд-во АН СССР, 1940. 212 с.
- Кочарян К. С. Эколого-экспериментальные основы зеленого строительства в крупных городах. М.: Наука, 2000. 184 с.
- Лунева З. С., Михайлов Н. Л., Судакова Е. А. Сирень. М.: Агропромиздат, 1989. 256 с.
- Мартынов Л. Г. Интродукция видов рода Сирень (Syringa) в условиях ботанического сада подзоны средней тайги//Известия Коми научного центра УрО РАН. 2013. № 4. С. 25-31.
- Пенкина И. Г. Сирени в Чуйской долине. Фрунзе: ИЛИМ, 1978. 56 с.
- Полякова Н. В. Биологические особенности представителей рода Syringa L. при интродукции в Башкирском Предуралье: Дис.. канд. биол. наук. Уфа, 2010. 188 с.
- Полякова Н. В., Путенихин В. П. Цветение сиреней в Башкирском Предуралье//Известия Самарского НЦ РАН. 2010. № 1 (3). С. 801-803.
- Попов М. Г. Очерк растительности и флоры Карпат. М.: МОИП, 1949. 303 с.
- Селенина Е. А., Седенин Н. А., Захарова М. А. Адаптация устьичного аппарата у тополя бальзамического (Populus balsamifera L.), сирени венгерской (Syringa josikaea Jacq. ex Rchb.) и яблони сибирской (Malus baccata (L.) Borkh.) к неблагоприятным условиям городской среды г. Красноярска//Вестник Иркутской государственной сельскохозяйственной академии. 2011. Вып. 44. С. 98-105.
- Яковлев Б. А. Климат Мурманска. Л.: Гидрометеоиздат, 1972. 107 с.
- Balaj N., Haxhinosto L., Hasani F., Lushi I., Balaj F. The study of flowering shrubs (Forsythia, Syringa, Cotoneaster) cultivars in the environment architecture and urbane landscape//Research Journal of Agricultural Science. 2012. Vol. 44 (2). P. 145-151.
- Brunsdon C., Comber L. Assessing the changing flowering date of the common lilac in North America: a random coefficient model approach//Geoinformatica. 2012. Vol. 16 (4). P. 675-690.
- Dadpour M., Naghiloo S., Peighambardoust S., Panahirad S., Aliakbari M., Movafeghi A. Comparison of floral ontogeny in wild -type and double flowered phenotypes of Syringa vulgaris (Oleaceae)//Scientia Horticulturae. 2011. Vol. 127 (4). P. 535-541.
- Densow B., Strzalkowska-Abramek M. Characteristics of blooming and pollen in flowers of two Syringa species (f. Oleaceae)//Acta Agrobotanica. 2013. Vol. 66 (4). P. 67-71.
- Dihoru G., Negrean G. The Red Book of Vascular Plants of Romania//Romanian Journal of Biology. 2009. № 54 (1). Р. 101-114.
- Jedrzejuk A., Lukaszewska A. High temperatures applied at fall forcing disturb ovule development in Syringa vulgaris L. "Mme Florent Stepman"//Acta Physiologiae Plantarum. 2008. Vol. 30. Issue 5. P. 673-678.
- Jedrzejuk A., Szlachetka W. Development of flower organs in common lilac (Syringa vulgaris L.) cv. Mme Florent Stepman//Acta Biologica Cracoviensia: Seris Botanica. 2005. Vol. 47. Issue 2. P. 41-52.
- Lendvay B., Pedryc A., Holm M. Characterization of Nuclear Microsattelite Markers for the Narrow Endemic Syringa josikae Jack. Fil ex Rchb.//Notulae Botanicae Horti Arrobotanici cluy -napoka. 2013. Vol. 41 (1). P. 301-305.
- Naghiloo S., Dadpour M., Gholamreza E., Peter K. Comparative study of inflorescence development in Oleaceae//American Journal of Botany. 2013. Vol. 100 (4). P. 647-663.
- Su G., Cao Y., Li C., Yu X., Tu P., Chai X. Photochemical and pharmacological progress on the genus Syringa//Chemistry Central Journal. 2015. Vol. 9 (2). P. 1-12.
- Wang H., Ge Q., Da J., Tao Z. Geographical pattern in first bloom variability and its relation to temperature sensitivity in the USA and China//I nternational Journal of Biometeorology. 2015. Vol. 59. Issue 8. P. 961-969.
