Динамика изменений большеберцового нерва при удлинении голени собак автодистрактором

Автор: Шевцов В.И., Варсегова Т.Н., Щудло М.М., Ерофеев С.А.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 1, 2004 года.
Бесплатный доступ
Изучали динамику изменения количественных показателей большеберцовых нервов на этапах удлинения голени 14 взрослых беспородных собак высокодробными автоматическими дистракторами. В конце дистракции на дистальном уровне средней 1/3 голени наблюдается уменьшение общей площади поперечного сечения нерва и суммарной площади пучков нервных волокон в сравнении с контролем (контралатеральный нерв). К концу фиксации нерв становится равномерно тоньше, через 30 дней после снятия аппарата площадь его поперечного сечения у двух животных восстанавливается, а у двух становится достоверно больше контрольных значений. Морфологические признаки тракционного повреждения отсутствуют. Происходящие изменения в соотношении внутрипучковых структурных компонентов нерва приводят к повышению объемной плотности миелиновых нервных волокон по сравнению с контралатеральным нервом, что является показателем восстановления морфологической структуры и функции нерва.
Автодистракция, большеберцовый нерв, морфометрия, стереология
Короткий адрес: https://sciup.org/142120688
IDR: 142120688
The dynamics of tibial nerve changes in the process of canine leg lengthening using an autodistractor
The dynamics of the changes of tibial nerve quantitative indices has been studied at the stages of leg lengthening in 14 adult mongrel dogs using automatic distractors of high division. Decrease of the total area of the nerve cross-section and the general area of nerve fiber bundles is noted at the end of distraction at the distal level of the leg middle 1/3 comparing with the control (contralateral) nerve. The nerve becomes uniformly thinner by the end of fixation, the area of its cross-section is restored in 30 days after the autodistractor removal in two animals and in the other two it becomes to be reliably more than the control values. There are no morphological signs of traction injury. The changes which occur in the correlation of the nerve intra-bundle structural components lead to the increase of the volume density of the nerve myelin fibers in comparison with the contralateral nerve, being an indicator of the recovery of the nerve morphologic structure and function.
Текст научной статьи Динамика изменений большеберцового нерва при удлинении голени собак автодистрактором
К настоящему времени в специальной литературе накопилось значительное количество фактов, свидетельствующих о высокой чувствительности периферической нервной системы к различным экспериментальным воздействиям [7, 10, 12]. Морфологическое описание берцовых нервов в условиях дистракционного остеосинтеза было предметом специальных исследований [3, 6, 8]. По мере разработки новых методических подходов возрастает интерес к изучению количественных показателей периферических нервов, отражающих полноту их функционального и морфологического
МАТЕРИАЛ
Исследовали большеберцовые нервы 14 взрослых беспородных собак, которым после закрытой флексионной остеоклазии удлиняли восстановления после различных видов повреждений [4, 9, 11] и объективно характеризующих направленность и временные изменения их морфофункционального состояния при удлинении конечности в высокодробных режимах. Учитывая отсутствие таких данных, мы поставили перед собой задачу изучить в эксперименте на животных динамику изменения гистоморфометрических показателей большеберцового нерва на этапах удлинения голени высокодробными автоматическими дистракторами.
И МЕТОДЫ голень на 15% в течение 28 дней аппаратом Илизарова с автоматическим приводом, обеспечивающим дистракцию костных фрагментов с суточной скоростью 1,0 мм и шагом 0,017 мм. Животных выводили из опыта передозировкой барбитуратов в конце дистракции (n=7), через месяц фиксации (n=3) и через месяц после снятия аппарата (n=4). Большеберцовые нервы контралатеральной (условный контроль) и опытной конечностей на уровне средней 1/3 голени закрепляли на твердой основе и иссекали для морфологического исследования. Проксимальный и дистальный концы иссеченных фрагментов нервов замораживали в жидком азоте и на микротоме-криостате МК-25 ТУ 64-1-856-78 при t = -18°С разлагали на тотальные поперечные срезы толщиной 7-10 мкм. В них кальцийкобальтовым методом [З. Лойда, 1982] выявляли активность миозиновой АТФ-азы. Остальной материал фиксировали в смеси 2% растворов глутарового и параформальдегидов на фосфатном буфере (рН 7,4), постфиксировали в 1% растворе тетраоксида осмия, дегидратировали в этаноле возрастающей концентрации и заливали в эпоксидные смолы. Поперечные полутонкие (0,5-1,0 мкм) срезы большой площади получали на ультратоме “Nova” фирмы LKB (Швеция), монтировали на заранее целлоидинированные предметные стекла и окрашивали раствором метиленового синего и основного фуксина.
Препараты изучали в большом исследовательском фотомикроскопе и стереомикроскопе фирмы “Opton”, оснащенных аппаратнопрограммным комплексом “DiaMorph”, что позволило оцифровывать изображения в ходе исследования. В цифровых изображениях гистохимических препаратов в программе “MEDIAS” (“DiaMorph”, Россия) измеряли общую площадь поперечного сечения нерва, суммарную площадь пучков нервных волокон вместе с окружающим их периневрием. Полученные значения сопоставляли по критерию Манна-Уитни (двусторонний вариант).
В цифровых изображениях полутонких срезов методом точко-счетной объемометрии рассчитывали относительный объем нервных волокон (V Vnf ), эндоневрия (V Vendo ) [1]. Определяли численные плотности мякотных нервных волокон (N Amnf ), их реактивно измененных форм (Na dmnf ) и долю последних в общем объеме выборки. Анализ цифрового материала проводили методами непараметрической статистики в программе Microsoft Excel 97 по W-критерию Вил-коксона для независимых выборок (статистические тесты запрограммированы И.П. Гайдыше-вым [2] и помещены в динамически подключаемую библиотеку).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Через 28 дней удлинения на дистальном уровне общая площадь поперечного сечения большеберцового нерва оперированной конечности и суммарная площадь пучков нервных волокон в 1,5 раза меньше, чем на контралатеральной стороне (рис. 1). На проксимальном уровне достоверные отличия по этому параметру отсутствуют (p>0,05).
Мякотные нервные волокна не обнаруживают патологических изменений, вместе с тем встречаются отдельные проводники крупного калибра с признаками реактивно-деструктивных изменений в виде вакуолизации аксона и дезинтеграции миелина, составляющие в среднем 1,78% в нервах опытной и 0,47% – контралатеральной конечностей.
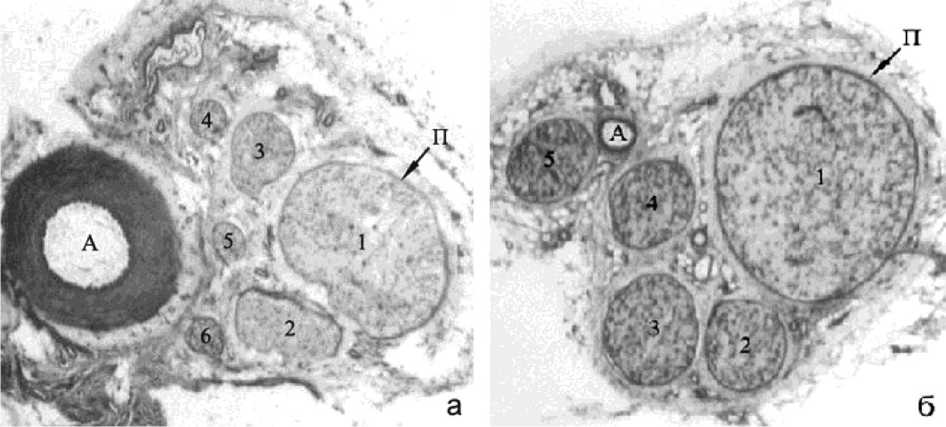
Рис. 1. Большеберцовый нерв, дистальный уровень, дистракция 28 дней: а) оперированная, б) контралатеральная конечности; 1-6 – пучки нервных волокон, П – периневрий, А – артерия. Криостатные срезы, реакция на миозиновую АТФ-азу. Об.- 1, ок.- 10х. Эксперимент № 2084
Объемная доля нейральных элементов в нерве оперированной конечности на 14,5% ниже, чем в контралатеральном нерве (p < 0,05) (табл. 1), а доля эндоневрия соответственно выше. Численная плотность мякотных волокон в удлинённом нерве незначительно выше, чем в контралатеральном (p>0,05). Как на оперированной, так и на контралатеральной стороне встречаются тонкие миелинизирующиеся ремаковские волокна малого калибра с разной степенью зрелости мякотной оболочки.
К концу дистракции в большинстве наблюдений животные пользовались удлиненной конечностью, приступая на нее в стойке или при ходьбе. Положение стопы, как правило, было близко к физиологическому.
В конце фиксации площадь поперечного сечения нерва оперированной стороны меньше (в среднем в 1,2 раза) на обоих уровнях. Соответственно уменьшена и суммарная площадь пучков.
Объёмная плотность нервных волокон незначительно (на 2,2%) ниже, а численная плотность мякотных волокон в удлинённом нерве на 18% выше (p < 0,05), чем на контралатеральной стороне. Как и на предыдущем сроке эксперимента, в нерве оперированной конечности содержатся незрелые миелинизирующиеся волокна.
На стороне удлинения продолжают встречаться единичные (0,45%) реактивно-измененные мякотные волокна, а в нервах контралатеральной конечности (рис. 2) количество их возрастает до 1,09% (табл. 1). При этом численная плотность деструктивно изменённых мякотных нервных волокон и их процентная доля в общем объёме вы- борки по сравнению с контралатеральной стороной уменьшается примерно в 2 раза.
Объемная плотность эндоневрия большеберцового нерва опытной конечности, как и на предыдущем сроке, превышает контрольные значения на 8% при p < 0,05 (табл. 1).
Нужно отметить, что в период фиксации все собаки активно нагружали оперированную конечность, амплитуда движений в коленном суставе приближалась к норме и составляла 90-100º (норма 100-110º), в голеностопном суставе сохранялось ограничение амплитуды движений – 50-60º.
Через месяц после снятия аппарата у двух животных общая площадь нерва и суммарная площадь пучков нервных волокон на опытной конечности в 1,3 раза больше (p < 0,05), чем на контралатеральной. У остальных двух общая площадь нерва достоверно не отличается от контроля, а суммарная площадь пучков меньше в 1,2 раза (p < 0,05) .
Объёмная плотность нервных волокон на этом сроке наблюдения оказывается выше на оперированной стороне (в среднем на 5,5%). Численная плотность мякотных волокон ниже, однако это различие статистически незначимо. Процентное содержание мякотных нервных волокон с признаками реактивно-деструктивных изменений на опытной и контралатеральной сторонах составляет 0,54 и 0,92% соответственно.
Объемная плотность внутрипучковой соединительной ткани понижена на 13% по сравне- нию с контролем, при p<0,05 (табл. 1).
Таблица 1
Морфометрические характеристики большеберцовых нервов собак на этапах эксперимента
|
Сроки |
Число опытов |
Конечность |
Vv nf |
Vv endo |
Na mnf |
Na dmnf |
Доля (%) dmnf |
|
Д - 28 |
7 |
оперированная |
50,6* |
49,4* |
22,5 |
0,4* |
1,78 |
|
контралатеральная |
65,1 |
34,9 |
21,4 |
0,1 |
0,47 |
||
|
Ф - 30 |
3 |
оперированная |
72,2* |
27,8* |
22,2 * |
0,1* |
0,45 |
|
контралатеральная |
74,4 |
25,6 |
18,3 |
0,2 |
1,09 |
||
|
БА - 30 |
4 |
оперированная |
62,6* |
37,4* |
18,6 |
0,1* |
0,54 |
|
контралатеральная |
57,1 |
42,9 |
21,7 |
0,2 |
0,92 |
* - различия между опытной и контралатеральной конечностью достоверны по W – критерию Вилкоксона для независимых выборок (p < 0,05).
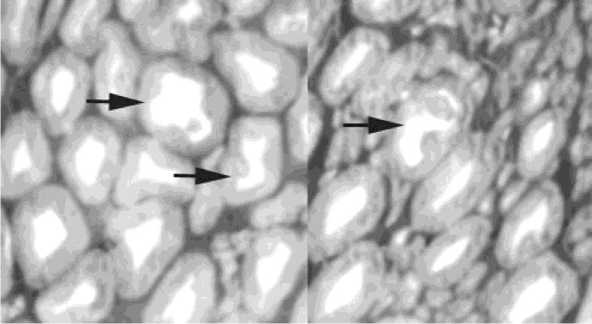
Рис. 2. Левый большеберцовый нерв. Полутонкий срез, окраска метиленовым синим-основным фуксином. Об. 40, ок. 12,5х. Эксперимент № 2210, дистракция 28, фиксация 30 дней. Стрелки – реактивноизмененные крупные мякотные нервные волокна
После снятия аппарата каких-либо ограничений функции оперированной конечности клинически выявлено не было.
Полученные результаты количественного исследования большеберцового нерва на разных этапах удлинения голени свидетельствуют о том, что в конце дистракции на оперированной стороне уменьшение общей площади поперечного сечения нерва и суммарной площади пучков нервных волокон наблюдается только на дистальном уровне средней 1/3 голени. Данный факт можно объяснить тем, что большеберцовый нерв связан с окружающими тканями в области голеностопного (скакательного) сустава и при удлинении голени его дистальный участок испытывает наибольшее растяжение. К концу периода фиксации нерв становится равномерно, но незначительно истонченным в сравнении с контралатеральной стороной, а через 30 дней после снятия аппарата площадь его поперечного сечения у двух животных восстанавливается, у двух же достоверно превышает контрольные значения.
Исследование показывает отсутствие каких-либо морфологических признаков тракционного повреждения нерва, что согласуется с данными K. Ikeda, K. Tomita, S. Tanaka [11], которые при удлинении бедра кролика с суточной скоростью 0,8-2,0-4,0 мм за 1 прием ни в одной из экспериментальных групп не обнаружили явлений валлеровской дегенерации волокон седалищного нерва. Нами отмечено развитие незначительных реактивных изменений толстых мякотных нервных волокон, которые, по мнению многих авторов [5, 6, 8], являются наиболее чувствительными к механическим воздействиям. В конце дистракции численная плотность реактивно- изменённых мякотных нервных волокон в четыре раза выше в нерве оперированной конечности, чем в контралатеральной, но на этапе фиксации и после снятия аппарата этот параметр в два раза выше в контралатеральном нерве. Их доля в общем объёме выборки меняется аналогично. Это свидетельствует не только об уменьшении действия факторов, влияющих на нервные волокна, но и о некотором приросте численности волокон с нормальной структурой.
Изменения соотношений площадей поперечного сечения нерва, суммарной площади его пучков, объёмной плотности нервных волокон и эн-доневрия, а также численной плотности мякотных волокон отражают различные процессы, протекающие на этапах эксперимента. Мы предполагаем, что это прежде всего результат перераспределения эндоневральной жидкости, изменения аксоплазматического тока, перекалибровки мякотных нервных волокон, новообразования и дифференцировки нервных проводников, а также морфогенетических процессов в оболочках нерва. Увеличение объёмной доли нейральных элементов и уменьшение доли изменённых нервных волокон в нерве оперированной конечности по сравнению с контралатеральным к концу эксперимента свидетельствует о восстановлении аксонального транспорта в зрелых волокнах и образовании новых нервных проводников.
Полученные в результате проведённого исследования данные об отсутствии морфологических признаков тракционного повреждения нерва указывают на малую травматичность оперативного вмешательства и удлинения конечности, а также о структурно-функциональной адаптации нейронов к увеличению длины конечности.
