Динамика экосистем северо-востока Волжского бассейна в голоцене

Автор: Дворников М.Г., Чащин П.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общие проблемы экологии
Статья в выпуске: 1-1 т.12, 2010 года.
Бесплатный доступ
В работе исследованы изменения, смены и преобразования экосистем в бореальном, атлантическом, суббореальном и субатлантическом климатических периодах голоцена. Установлено, что в заповедниках естественные условия и териокомплексы имеют сходство с прошлыми 4-3,5 тысячелетними зональными типами экосистем региона. Эти сведения могут использоваться в реконструкции экосистем голоцена.
Динамика, экосистемы, северо-восток волжского бассейна, голоцен
Короткий адрес: https://sciup.org/148198890
IDR: 148198890 | УДК: 56(471.34)
Dynamics of ecosystems of the north-east of the Volga river basin in Holocene
In this paper changes and transformations of ecosystems in boreal, Atlantic, subboreal and subarctic climatic periods of Holocene are studied. It was found out that in the preserves natural conditions and theriocomplexes are similar to the past 4-3.5-thousand-year zonal types of region ecosystems. Those data may be used for the reconstruction of Holocene ecosystems.
Текст научной статьи Динамика экосистем северо-востока Волжского бассейна в голоцене
Исследования эволюции экосистем Евразии при переходе от плейстоцена к голоцену (24-8 тыс. лет назад) на основе реконструкции обширных палеобиологических сведений уточняют и детализирует этапы и масштабы ранее известных представлений о послеледниковой эпохе [9]. Вместе с тем также известно, что в начале голоцена на одних и тех же хронологических отрезках в пределах разных территорий проходила динамика экосистем разного масштаба [15]. В частности, в течение голоцена происходили заметные изменения климата, площади лесотундры, среднета ё жных и южнота ё жных, смешанных и широколиственных лесов и лесостепи. Эти процессы усилились в связи со становлением и развитием к современному этапу человеческого общества в Среднем Поволжье, Предуралье и на Урале. В связи с отмеченным комплексное изучение региональной динамики экосистем в голоцене является одним из актуальных направлений исторической экологии.
На основе предложенного нами экологоисторического концептуального подхода проводились в регионе (51-63° с.ш. и 43-65° в.д.) изучения: структурно-функциональной организации биогеоценозов (БГЦ), ориентированные на оценку их состояния [4] и реконструкцию динамики экосистем на основе палеоботанических, палеозоологических и археологических сведений; инвентаризации современной биоты, структуры и продуктивности, фито- и зоомассы, параметров биологического круговорота, свойственных для зональных малонарушенных (заповедных) БГЦ и их более крупных иерархических единиц, находящихся в длительно динамичном развитии в широком диапазоне воздействия природных и антропогенных факторов [5, 6]. В данном случае мы расположили
Дворников Михаил Григорьевич , кандидат биологических наук, доцент кафедры экологи. Т. (88332) 574381; Чащин Павел Владимирович, младший научный сотрудник.
на карте известные археологические памятники в системе координат (с учетом их радиоуглеродных дат, здесь и далее), в соответствии с колебаниями влажности климата, динамикой границ экосистем, установленных на основе палинологических и раннеголоценовых фаунистических сведениях [2, 5, 7, 11, 13, 14, 16, 17 и др.], и провели анализ изменения и смены эдификаторов и фаунистических комплексов млекопитающих в БГЦ мезолита и современности.
Предложенный подход основан на том, что природная зональность — важнейший фактор эволюционных преобразований и географического распространения видов. Выявленные изменения эдификаторов и доминирующие в БГЦ млекопитающие, составляющие пищевые ресурсы человека, представлены в таблице.
В пребореальный и бореальный периоды (10,2-8,0 тыс. л.н.) в регионе и смеженных с ним территориях были расположены следующие экосистемы: 1) Разреженные сосновоберезовые леса с участками растительных сообществ тундр . В состав палеофитоценозов (между 61° и 67° с.ш.) входили представители темнохвойных и широколиственных пород наряду с небольшими участками тундростепных растительных ассоциаций. Сообщества млекопитающих были разделены на четыре экологические группы. К первой группе относились представители открытых ландшафтов, населяющие эти территории с плейстоцена — узкочерепная полевка, песец, северный олень и др. Во вторую группу входили виды, характерные для полуоткрытых и лесных ландшафтов — заяц-беляк, рыжая полевка, медведь и др. Третья группа состояла из широко распространенных азональных видов — бобр, водяная полевка, лисица и др. Четвертая группа была представлена одним видом — Mammuthus primigenius , последняя популяция которого в пребореале была приурочена к восточным областям данной экосистемы. 2) Сосново-березовые и сосново-ело-
Таблица 1. Изменения и преобразования экосистем в Вятско-Камском междуречье
|
у СУ X х X у у ф X с 2 3 S с У 8 |
Наименование торфяников и радиоуглеродные даты проб |
Координаты территории |
Характеристика условий, преобразований (эдификаторы и доминанты) в экосистемах |
|
к 00 ОО о ' и |
Чистое 8052+186 |
Повышенная влажность. Характерны разряженные сосновоберезовые леса со значительным участием карликовой березы, единична на севере ель. Содержание пыльцы Betula nana достигало 60%. В отдельных формациях на юге ель не отмечена. Млекопитающие: лось, северный олень, бобр, медведь, росомаха, куница. Из птиц доминируют белая куропатка и глухари. Плотность населения человека - менее 0,03 чел./км2. |
|
|
Каринский 8697+165 |
|||
|
Прокопьевский 9508+263 |
|||
|
у ч 00 И 5 5 К in < 00 |
Прокопьевский 4561+95 |
Чередование повышенной и пониженной влажности. В междуречье преобладают сосновые формации (до 60% пыльцы) с участием березы и широколиственных пород (лещина, вяз, липа, дуб, орешник, граб) от 5% на севере до 30% в центре и на юге. Встречается ель. Уменьшилось количество пыльцы Betula nana до 10%. Появились светлолюбивые папоротники Pieridium aqullinum . Млекопитающие: лось, северный олень, косуля, кабан, медведь, барсук, куница, бобр, белка, заяц-беляк. Птицы: глухарь, тетерев, рябчик. Плотность населения человека - 0,03-0,08 чел./км2. |
|
|
Чистое 5044+45 |
|||
|
Каринский 5612+170 |
|||
|
Прокопьевский 5739+117 |
|||
|
Лычное 6120+75 |
|||
|
Дымное 6471+197 |
59 ° 10'с.ш. 52 ° 30' в.д. |
||
|
Каринский 6589+110 |
|||
|
Муньковский 7778+99 |
|||
|
« к 5 в оо н О Q |
Каринский 2648+104 |
Влажность пониженная, в конце периода - повышенная. Преобладают хвойные формации. Еловые леса с большой долей сосны и березы в составе древостоев. Заметное участие в растительном покрове имеют широколиственные породы - 10-12%. Лесистость 97%. В конце этого периода прослеживаются антропогенные изменения почвенного и растительного покрова, и произошло его разделение на зоны тайги и подтайги. Лесистость последней 70%, южной части - 30%. Млекопитающие: на юге заметно присутствие домашних животных, обычны лось, северный олень, заяц-беляк, бобр, медведь, куница, лисица, волк. В средней и южной частях встречаются косуля и кабан. Плотность населения человека 0,09-4 чел./км2. |
|
|
Лычное 2656+166 |
|||
|
Прокопьевский 2666+89 |
58 ° 51'с.ш. 50 ° 25' в.д. |
||
|
Прокопьевский 2908+100 |
|||
|
Муньковский 4083+143 |
|||
|
Л и о S " и и X 00 g 8 К 1 cd । \о 4 ю CN |
Муньковский 1299+41 |
56 ° 54'с.ш. 50 ° 39' в.д. |
С начала и до средневековья пониженная, далее повышенная влажность. Еловые и елово-пихтовые с участием сосны средне- и южнотаежные леса (в том числе подтаежные). Уменьшение доли широколиственных пород на севере. Средняя тайга - лесистость более 80%, выше 59 ° с.ш; Южная тайга (от 59 ° с.ш. до 57 ° с.ш.) - лесистость 30-52%. Подтайга 57 ° -55 ° 50' с.ш. - лесистость 15-30%. На северо-востоке пихта до 40% в составе древостоев/ Млекопитающие: лось, северный олень, медведь, росомаха, колонок, куница, бобр, заяц-беляк, бурундук, белка. Северный олень, росомаха, бурундук на юге редки. Волк и лисица на севере и юге обычны. Птицы: глухарь, рябчик, тетерев, куропатка белая (в средней тайге обычна). В XX в. доля хвойных пород 50%, в лесах появились акк-лиматизанты: ондатра, американская норка, енотовидная собака, вновь появились кабан и косуля. Северный олень, европейская норка и белая куропатка внесены в Красную книгу. Плотность населения - 40 чел./км2. Доля пашни 22-37%. |
|
Лычное 1330+82 |
59 ° с.ш. 52 ° 16' в.д. |
||
|
Чистое 1979+114 |
58 ° 07'с.ш. 47 ° 30' в.д. |
||
|
Муньковский 2200+80 |
|||
|
Каринский 2440+107 |
58 ° 35'с.ш. 50 ° 20' в.д. |
вые леса с участием широколиственных пород в сочетании с лесостепными сообществами. Эти экосистемы расположены между 50° и 62° с.ш. Они являлись переходными между перигляциальными лесными сообществами позднеледни-ковья и межледниковыми лесными биомами голоцена. Это были ландшафты, представлявшие собой сочетание сосново-березовых и еловых лесов с участием широколиственных пород с участками лесостепных и тундростепных сообществ. Комплекс млекопитающих состоял из трех групп жизненных форм. К первой группе относятся виды, унаследованные от позднего плейстоцена и характерные для открытых ландшафтов — пищуха степная, узкочерепная полевка, печец, северный олень. Вторая группа включает представителей лесных и лесостепных биомов — крот европейский, заяц-беляк, куница, лось и др. 3) Лесостепные экосистемы занимали большую часть территории, за исключением горных областей. В них входили представители как лесных, так и степных палеофитоценозов. Сообщества млекопитающих включали в себя виды, объединенные в две группы: широко распространенные и характерные для лесостепных ландшафтов — заяц-русак, лесная мышь, кабан, барсук и др.
Особенностью фауны млекопитающих на Урале являлось сохранение на этой территории некоторых представителей мамонтового комплекса, позднее вымерших. В сообщества млекопитающих Урала входили виды различных экологических групп, адаптированных к ландшафтным условиям тундр, степей и лесов — пищуха степная, песец, северный олень, лось, медведь, а также азональные — бобр и др. [9]. Сохраняя преемственность с вышеотмеченным, нами были прослежены дальнейшие динамики экосистем.
В эпоху мезолита и неолита в условиях атлантического климата с распространением смешанных и широколиственных лесов 8052+186 — 4561+95 л.н. В бассейне р. Печоры и р. Вычегды (61-63° с.ш. и 48-57° в.д.) на стоянках первобытных охотников отмечены кости лося и бобра, встречались и кости северного оленя, медведя, волка, лисицы, т.е. 6 таежных видов. В средней части региона в бассейнах р. Вятка и Ветлуга стоянок было больше (56-58° с.ш. и 40-55° в.д.), в частности одними из характерных — Баринка II — 8265+130 л.н., Баринка I — 7435+70 л.н., Чернушка I — 6870+70 л.н. и т.д., далее эти территории занимали во-лосовские общности, возраст их поселений от 5150+40 до 4500+60 л.н. К примеру, Криушин-ская стоянка (57°40' с.ш. и 48°30' в.д.) находится вблизи заповедника «Нургуш», видовой состав добываемых зверей несколько иной, чем на севере: лось, бобр, северный олень, кабан, сибирская косуля, медведь, заяц-беляк, барсук, куница. На юге региона количество стоянок по рекам Кама, Белая, Д ё ма, Ик и их бассейнов более значительно, к примеру Деуковская II (55°50' с.ш. и 53°45' в.д.). На стоянках отмечены кости лося, кабана, зайца-беляка, бобра, медведя, лисицы, куницы и барсука. С наступлением суббореального периода с похолоданием и регрессией гидрологической сети в регионе происходит и сокращение ареала широколиственных лесов, увеличивается роль хвойных и мелколиственных элементов леса.
В период перехода от неолита к ранней бронзе с началом суббореального периода 4083+143 — 2648+104 л.н. более суровыми стали условия проживания людей и количество стоянок и поселений были ниже 58° с.ш., на отдельных стоянках охотников и рыбаков (Усть-Лудяна - 57°45' с.ш. и 49° в.д. и Май- данская, Волосовская культура - 56°10' с.ш. и 45° в.д.), до периода становления современных экосистем, уже используются в пищу домашние животные — крупный рогатый скот (КРС). Костные останки диких зверей составляют 94,1% [6]. Возраст данных энеолитических поселений был 4420+50 — 3975+80 л.н. В этот период стали преобладать еловые леса с участием сосны и березы. Вместе с тем, в это время при потеплении был и возврат широколиственных пород к северу, то есть до 63° с.ш.
В переходный период поздней бронзы к эпохе железа в рассматриваемом регионе, в условиях субатлантического периода 2656+166 л.н. — современность, хозяйственная деятельность срубной, приказанской и балановской культур, в наступающее повышенное увлажнение климата, сменяется культурой финноугорских плем ё н, хорошо владеющих навыками охоты. К этому периоду уже закончилось становление современных лесорастительных районов в пределах средней и южной тайги, в то же время сюда проникли пихта и значительно на юге были преобразованы человеком подтаежные и широколиственные леса. В регионе уже стало добываться 13 видов зверей, при этом во всех типах экосистем осваивались ресурсы копытных и значительно зайца, белки, куницы, лисицы, выдры и бобра. В южнота ё жных экосистемах охота и рыболовство ещ ё имели важное значение в жизнеобеспеченности населения (до 39% костных останков) и даже активизировались по сравнению с прошлой эпохой.
Последующий этап развития человеческого общества, преобразование им экосистемы, их состояние прослеживается уже по историческим сведениям. Расселение человека стало основано на непрерывном преобразовании экосистем. Поселения с высокой плотностью населения, развитой культурой, животноводством, земледелием, зачатками производства имели больше шансов осваивать территориальные пространства, эффективно добывать и использовать природные ресурсы (см. табл.). В частности, заселение в VI в. до н.э. бассейнов рр. Ветлуга, Вятка, Кама и Северного Приуралья шло на взаимосмешении местного населения с пришлым. Интенсивно осваивались территории от Самарской Луки на юге до правобережья р. Камы на севере, в том числе Башкирское Пре-дуралье, долины не только крупных, но и малых рек и водоразделы, которые в предшествующее время не были заселены человеком. Пришельцы владели мастерством пашенного земледелия и металлообработки, разводили мясомолочный и тягловый скот, имели плуги, косы-горбуши, серпы, возделывали рожь, пшеницу, полбу, овёс, горох [3]. С VIII по XIV в. основано было одно из первых на Востоке Евро- пы феодальное государство. С XII по XIV в. север региона осваивают русские, а южную его часть с XIII в. — татары. Именно в историческом периоде интенсивно уменьшилась площадь лесов и изменился их состав, в частности в спектрах пыльцы стало больше лиственных пород. Вместе с тем, в малонарушенных территориях, ныне в условиях заповедников, сохранились природные условия, структура лесонасаждений и параметры биогеоценотических процессов, свойственные зональным экосистемам [5].
Наряду с активными этническими процессами в регионе при снижении влажности климата изменялись и масштабы использования природных ресурсов, менялся состав домашних животных [6]. В регионе осваивалось охотниками 22 вида диких млекопитающих. Возрастающий спрос на пушнину привел к потере промысловых ресурсов бобра, соболя и др. Развитие динамики раннеголоценовых фаунистических комплексов млекопитающих можно проследить на сравнительном материале. К примеру, рассмотрим списки видов локальных фаун бореальных и степных биомов, азональных и интразональных территорий, составленных на основе опубликованных сведений по заповедникам (с севера к югу от Печоро-Илычского до Оренбургского и с запада к востоку от Керженского до Висимского заповедников) Предуралья и Урала и прилегающих территорий [5]. Всего нами выделено 64 индикаторных вида для 15 локальных фаун, которые распространены в регионе.
Бореальные биомы и интразональные территории. А). Предуралье, Северный Урал: 1. Хвойные леса равнин и возвышенностей; 2. Смешанные леса равнин и возвышенностей; 3. Хвойные леса предгорий и гор; 4. Смешанные леса предгорий и гор; 5. Горные луга, тундры. Б). Предуралье, Средний Урал. 6. Хвойные леса предгорий и гор; 7. Смешанные леса предгорий и гор; 8. Горные луга, тундры. В). Предуралье, Южный Урал: 9. Хвойные леса возвышенностей и гор; 10. Смешанные леса возвышенностей и гор; 11. Широколиственные леса возвышенностей и гор; 12. Горные луга, тундры. Биомы степей: Предуралье, Южный Урал. 13. Колки, поймы лесостепи; 14. Луговая степь, овраги лесостепи; 15. Степь.
Для сравнительного анализа современных и раннеголоценовых фаун более крупных таксонов (Северный, Средний и Южный Урал) были выделены 6 фаун: А. Рецентная фауна Северного Урала; Б. Рецентная фауна Среднего Урала; В. Рецентная фауна Южного Урала; Д. Раннеголоценовая фауна Северного Урала; Е. Раннеголоценовая фауна Среднего Урала; Ж. Раннеголоценовая фауна Южного Урала. При этом мы использовали опубликованные фауни стические списки млекопитающих Урала, свойственные для раннего голоцена [8]. Фаунистические списки современной фауны млекопитающих заповедников Северного, Среднего, Южного Урала и прилегающих территорий получены на основе объединения списков фаун лесов, горных лугов и тундр соответствующих таксонов более низкого ранга и адаптации их к фаунистическим спискам млекопитающих раннего голоцена. Сравнение фаун проводили с помощью индекса сходства (общности) Чека-новского-Соренсена (Czekanovski-Dice-
Sorensen) для качественных данных:
2a
ICS =___ (a + b) + (a + c)
2 a где a — число видов, присутствующие в обоих списках; b — число видов, имеющихся только в j-м списке; c — число видов, принадлежащих только к-му списку.
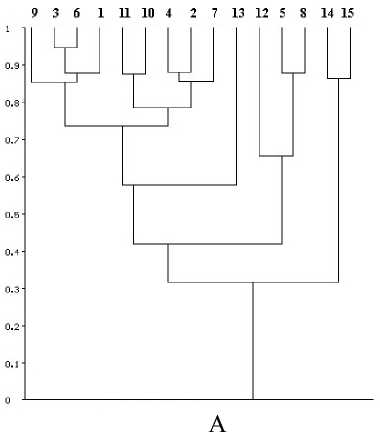
Индекс I CS линейно зависит от различий по числу видов, а также от относительного числа общих видов в сравниваемых списках. Это достоинство определяет его широкое применение в фаунистических исследованиях [12]. Индекс I CS игнорирует число отрицательных совпадений (число видов, отсутствующих в обоих списках). Уровень видового богатства (УВБ) оценивали как суммарное число видов, зарегистрированных на каком-либо конкретном территориальном таксоне (группе таксонов). Концентрацию видового богатства (КВБ) оценивали как долю видового богатства отдельного территориального таксона (группы таксонов) от общего состава фауны (%). Вычисления индексов и построение дендрограмм по методу не взвешенной средней (UPGMA-метод) выполняли с помощью программы BIODIV [1]. Результаты оценки сходства локальных современных видов фаун млекопитающих бореальных и степных биомов Урала ( I CS) приведены на дендрограмме (рис. А).
Отмечено, что в первом случае выделились 4 удаленных друг от друга кластера. В первый кластер входят фауны лесных биомов Северного, Среднего и Южного Урала. В его составе два субкластера — фауны хвойных лесов и фауны смешанных и широколиственных лесов.
Последний субкластер в свою очередь состоит из двух субкластеров более низкого порядка — фауны Южноуральских горных смешанных и широколиственных лесов и фауны смешанных лесов Северного и Среднего Урала. Все перечисленные фауны характеризуются высоким уровнем сходства (ICS > 0,7). Следующий кластер представлен фауной колков и пойм лесостепей Южного Урала и Предуралья. Он занимает промежуточное положение между фаунами открытых пространств и лесных фаун (0,5 < ICS < 0,6). В составе следующего класте- pa — фауны горных тундр. Уровень сходства с предыдущими кластерами низкий (0,4 < ICS < 0,5). Наиболее удален от других кластер представленный биомами степей. Уровень сходства его с остальными кластерами самый низкий (0,3 < ICS < 0,4). Во втором случае степень и характер сходства фаун отображены на дендро грамме (рис. Б). Современные и раннеголоце-
новые фауны образуют два отдельных кластера (0,6 < I CS< 0,7). Небольшие отличия связаны с присутствием в составе раннеголоценовых фаун плейстоценовых реликтов, как вымерших (бизон, шерстистый носорог, гигантский олень и др.), так и существующих в настоящее время (пищуха степная, хомячок Эверсманна и др.).
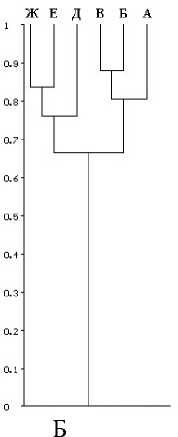
Рис. Дендрограмма сходства: современных видов фаун млекопитающих бореальных и степных биомов Урала — А; современных и раннеголоценовых фаун — Б
Уровень и концентрация видового богатства рассмотренных современных локальных фаун (1—15) в целом соответствуют известным закономерностям [9, 10]. Они повышаются в направлении с севера на юг (Северный Урал — Средний Урал — Южный Урал) и при снижении высоты над уровнем моря (тундры — хвойные леса — смешанные леса Предуралья и Урала). В степных биомах уровень и концентрация видового богатства снижаются. Самые низкие показатели уровня видового богатства и, соответственно, концентрации видового богатства в биомах тундр, самые высокие — в биомах смешанных лесов. Для современной фауны таксонов более высокого ранга также были рассчитаны показатели УВБ и КВБ. Указанные выше закономерности относительно уровня и концентрации видового богатства соблюдаются и для таксонов высшего ранга. Это объясняется наличием на Южном Урале более широкого набора экологических ниш. Из данного примера следует, что в рассматриваемом регионе формирование фаунистических комплексов развивалось динамично, в то же время векторизовано, интенсивность процессов проявлялась, согласно развития зональных природных условий. Современный состав фаунистических комплексов малонарушенных экосистем заповедников имеет большое сходство с таковыми, характерными для суббореальных и субатлантических природных условий голоце на. Именно рубеж 3,5-3 тыс. л.н. является исходным для исчисления возраста современных БГЦ в Вятско-Камском междуречье..
Кабан и сибирская косуля вновь были отмечены в добыче охотников уже в современных и измененных, частично преобразованных, человеком экосистемах VI-XIV и XX вв. В последние 100 лет в связи с интенсивным преобразованием человеком зональных границ, изменением структуры лесов и интродукцией ряда промысловых зверей, количество таксонов увеличилось в таежных, смешанных лесных формациях (см. табл.). Площади последних увеличились во всех зонах, эти условия послужили для расселения косули, кабана и др. видов к северу. В итоге, данные о биоте заповедников, вполне аргументировано, можно использовать как эталоны и в реконструкции экосистем.
Рассмотренные материалы позволили нам выделить и динамику разного масштаба в развитии экосистем региона.
При интенсивной охоте в мезолите-неолите без изменения экосистем человеком происходили естественные колебания структуры фаунистических комплексов в соответствии с развитием БГЦ.
При изменении экосистем в эпохи бронзы и железа в результате пастьбы, выжигания лесов, распашки земель происходили и смены структуры фаунистических комплексов.
При глобальных преобразованиях в историческое время лесостепных и лесных экосистем и биомов происходило смешение фаунистических комплексов, только в малонарушенных территориях сохранилась их структура и плотность населения характерная для зональных типов.
ÑÏÈÑÎÊ ËÈÒÅÐÀÒÓÐÛ
-
1. Баев П.В., Пенев Л.Д. BIODIV — программа для вычисления параметров биологического разнообразия и кластерного анализа. М., 1990.
-
2. Благовещенская Н.В. Динамика лесных экосистем верхнего плато Приволжской возвышенности в голоцене // Экология. 2006. № 2.
-
3. Голдина Р.Д. Вклад Е.А. Халиковой в изучение этнической принадлежности средневековых памятников Южной Удмуртии // Вопросы древней истории Волго-Камья. Казань: Мастер Лайн, 2002.
-
4. Дворников М.Г. Комплексные экологические исследования в пойменных биогеоценозах р. Вятка // Самарская Лука: Бюл. 2004. № 15.
-
5. Дворников М.Г. Млекопитающие в экосистемах бассейна реки Вятка (на примере особо охраняемых и освоенных территорий). Киров, 2007.
-
6. Дворников М.Г. Ретроспективный анализ животноводства в экосистемах Северо-Востока Волжского бассейна // Изв. Самар. НЦ РАН. 2009. Т. 11. № 1.
-
7. Косинцев П.А., Симакова А.Н., Пузаченко А.Ю., Ванн Кольфсхотен Т Экосистемы раннего голоцена (пребореал-бореал РВ-ВО <10,2->=8,0 тыс.л.н.) // Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24-8 тыс. л.н.). М.: Товарищество научных изданий КМК, 2008.
-
8. Косинцев П.А. Комплексы млекопитающих раннего голоцена (пребореал-бореал <10,2->=8,0
-
9. Маркова А.К., Т. ван Кольфсхотен, С. Бохнкке, Косинцев П.А., И. Мол, Пузаченко А.Ю., Симакова А.Н., Смирнов Н.Г., А. Верпоорте, Головачев И.Б. Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24-8 тыс. л.н.). М.: Товарищество научных изданий КМК, 2008.
-
10. Мордкович В.Г., Баркалов А.В., Василенко С.В . и др . Видовое богатство членистоногих ЗападноСибирской равнина // Евразиатский энтомол. жур-н. 2002. Т. 1. Вып. 1.
-
11. Петренко А.Г. Древнее и средневековое животноводство среднего Поволжья и Предуралья. М.: Наука, 1984.
-
12. Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982.
-
13. Прокашев А.М., Жуйкова И.А., Пахомов М.М. История почвенно-растительного покрова Вятско-Камского края в послеледниковье. Киров, 2003.
-
14. Рябова Т.П. Развитие растительности Башкирского Предуралья в голоцене // Научные доклады Высшей школы. Биологические науки. 1965.
-
15. Смирнов Н.Г. Динамика видов и их комплексов как предмет исследований исторической экологии // Экология. 2006. № 6.
-
16. Шаландина В.Г. Растительный покров северо-запада Татарии в позднем голоцене и его изменения под влиянием антропогенных факторов // Антропогенные факторы в истории развития современных экосистем. М.: Наука, 1981.
-
17. Шаландина В.Г. История растительного покрова Северо-востока Приволжской возвышенности в голоцене // Самарская Лука: Бюл.. 1993. № 4.
тыс.л.н.) // Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24-8 тыс. л.н.). М.: Товарищество научных изданий КМК, 2008.
DYNAMICS OF ECOSYSTEMS OF THE NORTH-EAST OF THE VOLGA RIVER BASIN IN HOLOCENE
Список литературы Динамика экосистем северо-востока Волжского бассейна в голоцене
- Баев П.В., Пенев Л.Д. BIODIV -программа для вычисления параметров биологического разнообразия и кластерного анализа. М., 1990.
- Благовещенская Н.В. Динамика лесных экосистем верхнего плато Приволжской возвышенности в голоцене//Экология. 2006. № 2.
- Голдина Р.Д. Вклад Е.А. Халиковой в изучение этнической принадлежности средневековых памятников Южной Удмуртии//Вопросы древней истории Волго-Камья. Казань: Мастер Лайн, 2002.
- Дворников М.Г. Комплексные экологические исследования в пойменных биогеоценозах р. Вятка//Самарская Лука: Бюл. 2004. № 15.
- Дворников М.Г. Млекопитающие в экосистемах бассейна реки Вятка (на примере особо охраняемых и освоенных территорий). Киров, 2007.
- Дворников М.Г. Ретроспективный анализ животноводства в экосистемах Северо-Востока Волжского бассейна//Изв. Самар. НЦ РАН. 2009. Т. 11. № 1.
- Косинцев П.А., Симакова А.Н., Пузаченко А.Ю., Ванн Кольфсхотен Т. Экосистемы раннего голоцена (пребореал-бореал РВ-ВО =8,0 тыс.л.н.)//Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24-8 тыс. л.н.). М.: Товарищество научных изданий КМК, 2008.
- Косинцев П.А. Комплексы млекопитающих раннего голоцена (пребореал-бореал =8,0 тыс.л.н.)//Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24-8 тыс. л.н.). М.: Товарищество научных изданий КМК, 2008.
- Маркова А.К., Т. ван Кольфсхотен, С. Бохнкке, Косинцев П.А., И. Мол, Пузаченко А.Ю., Симакова А.Н., Смирнов Н.Г., А. Верпоорте, Головачев И.Б. Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24-8 тыс. л.н.). М.: Товарищество научных изданий КМК, 2008.
- Мордкович В.Г., Баркалов А.В., Василенко С.В. и др. Видовое богатство членистоногих Западно-Сибирской равнина//Евразиатский энтомол. журн. 2002. Т. 1. Вып. 1.
- Петренко А.Г. Древнее и средневековое животноводство среднего Поволжья и Предуралья. М.: Наука, 1984.
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982.
- Прокашев А.М., Жуйкова И.А., Пахомов М.М. История почвенно-растительного покрова Вятско-Камского края в послеледниковье. Киров, 2003.
- Рябова Т.П. Развитие растительности Башкирского Предуралья в голоцене//Научные доклады Высшей школы. Биологические науки. 1965.
- Смирнов Н.Г. Динамика видов и их комплексов как предмет исследований исторической экологии//Экология. 2006. № 6.
- Шаландина В.Г. Растительный покров северо-запада Татарии в позднем голоцене и его изменения под влиянием антропогенных факторов//Антропогенные факторы в истории развития современных экосистем. М.: Наука, 1981.
- Шаландина В.Г. История растительного покрова Северо-востока Приволжской возвышенности в голоцене//Самарская Лука: Бюл.. 1993. № 4.
