Динамика кадмиевой устойчивости проростков кукурузы и содержание в них растворимых углеводов в условиях наличия или отсутствия теплового закаливания

Автор: Чудинова Лариса Алексеевна, Чернышева Юлия Дмитриевна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2019 года.
Бесплатный доступ
Изучалось изменение устойчивости проростков кукурузы к действию 1мМ раствора ацетата кадмия в условиях наличия или отсутствия предобработки закаливающей температурой (+40°С, 3 ч), а также ауксином (ИУК). Установлено формирование двух фондов глюкозы и сахарозы - свободного (СФ) и компартментированного (КФ) в побегах в нормальных условиях и их перераспределение при кадмиевом стрессе. Показано, что тепловая предобработка, а также экзогенная ИУК способствуют значительному росту уровня как СФ, так и КФ глюкозы в условиях последующего действия кадмия. Сделано предположение, что стресс-индуцируемое изменение компартментации растворимых сахаров является условием реализации их защитных функций, которые зависят не столько от количества растворимых сахаров в клетках, сколько от их способности к метаболизации. Предполагается, что при кратковременном воздействии кадмия (стадия стресс-реакции) оба пула сахаров обеспечивают сочетание повышенного уровня жизнедеятельности и устойчивости, а при более длительном (стадия адаптации) - участвуют в формировании металлоустойчивости растений.
Проростки кукурузы, ацетат кадмия, тепловое закаливание, глюкоза, сахароза, свободная и компартментированная фракции
Короткий адрес: https://sciup.org/147227067
IDR: 147227067 | УДК: 581.1:632.122.1 | DOI: 10.17072/1994-9952-2019-78-83
Dynamics of cadmium resistance of maize seedlings and the content of soluble carbohydrates in terms of the presence or absence of thermal hardening
A study was made of the change in the stability of maize seedlings to the action of a 1 mM solution of cadmium acetate under the conditions of the presence or absence of pretreatment by hardening temperature (+ 40°C, 3 hours), as well as auxin (IAA). The formation of two funds of glucose and sucrose - free (SF) and compartmented (CF) in the shoots of maize under normal conditions and their redistribution under cadmium stress has been established. It is shown that thermal pretreatment, as well as exogenous IAA, contributes to a significant increase in the level of both SF and CF glucose under conditions of subsequent action of cadmium. It has been suggested that the stress induced change in the compartmentation of soluble sugars is a condition for the realization of their protective functions, which depend not so much on the amount of soluble sugars in the cells, but on their ability to metabolize. It is assumed that with short-term exposure to cadmium (the stress-reaction stage) both sugars pools provide a combination of an increased level of vital activity and sustainability, and with a longer-term (adaptation stage) they participate in the formation of metal resistance of plants.
Текст научной статьи Динамика кадмиевой устойчивости проростков кукурузы и содержание в них растворимых углеводов в условиях наличия или отсутствия теплового закаливания
Явление, когда действие одного неблагоприятного фактора среды приводит к повышению устойчивости растений к другим факторам названо перекрестной адаптацией или «кросс-адаптацией». Оно известно довольно давно, но до сих пор его молекулярнобиохимические механизмы остаются до конца не выясненными [Кузнецов, Дмитриева, 2005]. В отношении комбинированного действия на растения тяжелых металлов и неблагоприятных температур в литературе в настоящее время имеются лишь единичные
исследования, результаты которых противоречивы [Титов, Казнина, Таланова, 2014].
Имеются основания считать, что в процессе перекрестной адаптации участвуют разнообразные низкомолекулярные соединения (пролин, органические кислоты, растворимые сахара, полиамины и др.), большинство из которых проявляют полифункциональность и выполняют протекторные функции [Чудинова, Суворов, 2011].
Направленность превращений углеводноферментного комплекса в этих условиях до настоящего времени остается предметом дискуссии. Одни авторы считают, что это результат деструктивных процессов, связанных с повреждением клеток в условиях стресса, другие – одной из составляющих комплекса защитных реакций на стресс [Колупаев, Трунова, 1992; Титов, Казнина, Таланова, 2014]. При этом участие углеводного обмена в формировании устойчивости растений к токсическому действию тяжелых металлов практически не изучено. Кроме того, в большинстве работ исследовалась динамика концентраций углеводов в растительных клетках в состоянии стресса, но не учитывалась их компартментация (или разделение на отдельные фонды). Необходимость компартменти-рования низкомолекулярных сахаров определяется их полифункциональностью на уровне клетки. Проявляется компарментация в том, что в определенных физиологических или биохимических процессах участвует не вся масса внутриклеточного вещества, а только некоторая его часть; иными словами – в разной доступности разных фондов для определенных функций, что обеспечивается наличием внутриклеточных мембран [Ходос, 1975; Холодова, 1978]. Не исключена, хотя и занимает меньшее место, компартментация, основанная на различии свойств отдельных зон цитозоля в пределах клетки.
В связи с этим цель данной работы – изучение динамики кадмиевой устойчивости проростков кукурузы и содержания в них свободной (СФ) и ком-партментированной (КФ) фракций глюкозы и сахарозы в условиях наличия или отсутствия предобработки повышенной температурой, а также ауксином (ИУК).
Объект и методы исследований
Исследования проводили на проростках кукурузы сахарной (Zea mays L.) (сорт Кубанская). Семена стерилизовали в растворе перманганата калия, выращивали на вермикулите в климатической камере ШН-М при температуре 20–22°C, влажности 75% и освещенности 16 ч. в сут. (1-й вариант – контроль). Затем 12-суточные проростки подвергали стрессовым воздействиям по следующей схеме: 2-й вариант – экспозиция в 1мМ растворе ацетата кадмия; 3-й вариант – экспозиция в 1мМ растворе ацетата кадмия + ИУК (10мг/л); 4-й вариант – проростки выдерживали 3 ч. при 40°С, затем выращивали в 1мМ растворе ацетата кадмия; 5-й вариант – проростки выдерживали 3 ч при 40°С, затем выращивали в 1мМ растворе ацетата кадмия с добавлением ИУК (10мг/л).
Анализировали интенсивность роста по длине побега и его сырой массе на 3-, 7- и 10-е сут. кадмиевой экспозиции. Содержание СФ сахаров (экстракция из гомогената 10 мин. при 80°С) и КФ сахаров (экстракция из гомогената 50 мин. при 80°С) определяли по методу Вознесенского в 3-кратной повторности [Большой практикум …, 2012]. При статистической обработке данных использовали программу Stadia.
Результаты и их обсуждение
Поскольку интенсивность роста выступает как интегральный процесс, отражающий итог всех функциональных и метаболических изменений в растении, то для оценки динамики устойчивости проростков к кадмиевому стрессу мы использовали такие биометрические ростовые показатели, как длина и сырая масса побегов. Полученные нами результаты показали ингибирующее действие 1 мМ раствора ацетата кадмия на интенсивность роста проростков во все сроки определения, особенно при увеличении продолжительности воздействия до 10 сут. (2 вар.). В последнем случае интенсивность роста уменьшилась на 20% относительно контроля (рис. 1).
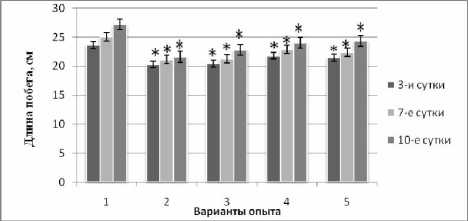
Рис. 1 . Изменение длины побегов кукурузы при кадмиевом стрессе в условиях наличия или отсутствия предобработки повышенной температурой и ИУК:
1 – контроль; 2 – экспозиция при 1мМ ацетате кадмия; 3 – экспозиция при 1мМ ацетате кадмия + ИУК (10мг/л); 4 – 3 ч. при 40°C, затем экспозиция при 1мМ ацетате кадмия; 5 – 3 ч. при 40°C, затем экспозиция при 1мМ ацетате кадмия + ИУК (10мг/л). * – различие с контролем достоверно
При комбинировании тепловой предобработки и последующего монодействия кадмия (4 вар.) было отмечено достоверное снижение торможения интенсивности роста проростков по сравнению со 2-м вариантом, в котором тепловая предобработка отсутствовала. Положительный эффект теплового воздействия на устойчивость проростков к кадмиевому стрессу сохранялся при дополнительном введении в корневую среду ИУК (5 вар.). В отношении сырой биомассы побегов мы наблюдали сходную динамику, но с меньшей амплитудой.
Установленное нами достоверное повышение кадмиевой устойчивости проростков кукурузы после предварительной тепловой обработки по-видимому является результатом теплового закаливания, что согласуется с нашими исследованиям на проростках овса [Чудинова, Чернышева, 2017]. Известно, что тепловое закаливание растений, как правило, сопровождается изменениями термостабильности белков и определенных функций клеток [Титов, Казнина, Таланова, 2014], в результате чего могут активироваться защитно-приспособительные процессы и формироваться, в частности, повышенная металлоустойчивость. Можно предположить, что положительное действие ИУК на устойчивость растений к кадмию также связано с запуском комплекса защитных реакций, в том числе и с активацией экспрессии генов антиоксидантных ферментов.
Полученные нами результаты (рис. 2, 2 вар.) показали разную динамику содержания СФ и КФ глюкозы в побегах кукурузы в условиях ингибирования роста при монодействии кадмия в зависимости от стадии адаптации к данному стрессору. Так, 3-суточная экспозиция в ацетате кадмия вызывала значительное снижение, а 7 и 10-суточная – увеличение содержания обеих фракций глюкозы.
Можно предположить, что в ранние сроки адаптации снижение уровня СФ и КФ глюкозы происходит за счет ее интенсивного катаболизма, направленного на поддержание структурного и энергетического обмена. Как отмечали некоторые авторы [Колупаев, Трунова, 1992; Дерябин и др., 2007], уменьшение содержания растворимых сахаров в первые сутки действия стрессора может быть связано с расходованием глюкозы на перехват активных форм кислорода и включением альтернативных путей дыхания.
Установленное нами повышенное содержание СФ и КФ глюкозы на 7- и 10-е сут., очевидно, свидетельствует о более высоком метаболитическом и энергетическом статусе растений, который достигается в результате развития процесса адаптации в эти сроки экспозиции на растворе кадмия.
Считается, что повышенное содержание глюкозы и других растворимых углеводов помогает растениям перенести стресс, значительно повышая их устойчивость, поскольку они являются источниками энергии и пластических веществ, осмолитами, основной транспортной формой органических веществ, участвуют в «тушении» окислительного стресса, проявляют антиденатурирующее действие на белково-липидный комплекс мембран, стабили- зируют белки, подвергнутые нагреву [Кафи, Стюарт, Борланд, 2003; Дерябин и др., 2007].
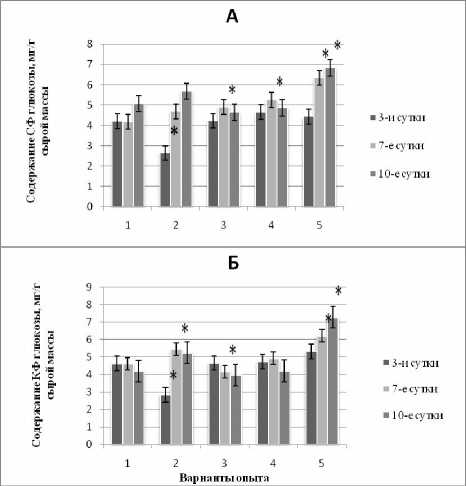
Рис. 2. Динамика СФ (А) и КФ (Б) глюкозы в побегах кукурузы при кадмиевом стрессе в условиях наличия или отсутствия предобработки повышенной температурой и ИУК
Накопление растворимых сахаров у одних растений определяется главным образом высокой интенсивностью фотосинтеза, а у других – гидролизом запасных полисахаридов, прежде всего – крахмала. Активация гидролитических ферментов (инвертазы и амилазы) при разрушении комплекса «фермент – белок-ингибитор», т.е. без участия бе-локсинтезирующего аппарата, приводит к быстрому увеличению количества глюкозы. Другой причиной роста уровня моносахаридов может быть торможение при стрессе их оттока из листьев в нефотосинтезирующие органы.
Многие авторы положительное влияние теплового закаливания на последующую устойчивость растений к разным стрессорам связывают с индукцией синтеза белков теплового шока (БТШ) [Кар-пец, Колупаев, 2009]. Имеются сведения об активном участии БТШ в защите клетки от токсичного действия тяжелых металлов. Так, показано, что соли кадмия способны индуцировать у растений синтез БТШ с молекулярными массами 70, 42, 40, 26, 23, 15 и 11 кДа [Титов, Казнина, Таланова, 2014].
Экзогенная ИУК в нашем эксперименте способствовала значительному росту уровня как СФ, так и КФ глюкозы во все сроки определения в условиях последующего действия кадмиевого стресса. Это согласуется с данными литературы, в которых указывается на участие ИУК, наряду с другими сигнальными молекулами, в активации процессов накопления низкомолекулярных протекторов при различных стрессах [Карпец, Колупаев, 2009].
По мнению ряда авторов [Колупаев, Трунова, 1992; Синькевич и др., 2008], одним из механизмов адаптации растений к абиотическому стрессу является накопление сахарозы, которая имеет двойную функцию – транспортную и сигнальную, регулирующую экспрессию генов и развитие растений.
Известно, что синтез сахарозы происходит в цитозоле клеток мезофилла листа на свету. Часть сахарозы, распадаясь на глюкозу и фруктозу, используется в процессе метаболизма, оставшаяся сахароза поступает во временное депо ассимилятов – вакуоль [Кузнецов, Дмитриева, 2005]. Метаболизм сахарозы характеризуется большой гибкостью, что определяется активностью трех ферментов: инвертазы, основная реакция которой – необратимое расщепление сахарозы на глюкозу и фруктозу, а также сахарозофосфатсинтетазы и са-харозосинтетазы. Гены этих ферментов различаются пространственно временной экспрессией в процессе онтогенеза и уровнями регуляции. В растениях зафиксировано увеличение активности инвертазы и сахарозосинтезирующих ферментов, а также изменение их изоферментного спектра в ответ на стрессы самой различной природы [Дерябин и др., 2007; Синькевич и др., 2008].
В нашем эксперименте была выявлена КФ сахарозы (вакуолярная фракция), тогда как СФ сахарозы (цитоплазматическая фракция) практически не определялась (рис. 3). В условиях монодействия кадмия (2 вар.) содержание КФ сахарозы на 10-е сут. экспозиции было на уровне контроля. В вариантах с тепловым закаливанием, а также в присутствии ИУК (4 и 5 вар.) количество сахарозы уменьшилось в 2 раза по сравнению с контролем. Похожая динамика сахарозы была отмечена нами ранее в эксперименте с проростками овса [Чудинова, Чернышева, 2017]. Низкое содержание или отсутствие в пробах сахарозы при тепловом закаливании, возможно, свидетельствует об интенсивном расщеплении этого дисахарида, что согласуется с достаточно высоким уровнем глюкозы в этих условиях. Возможно, что перераспределение моно- и дисахаров является следствием изменения активности разных форм инвертазы.
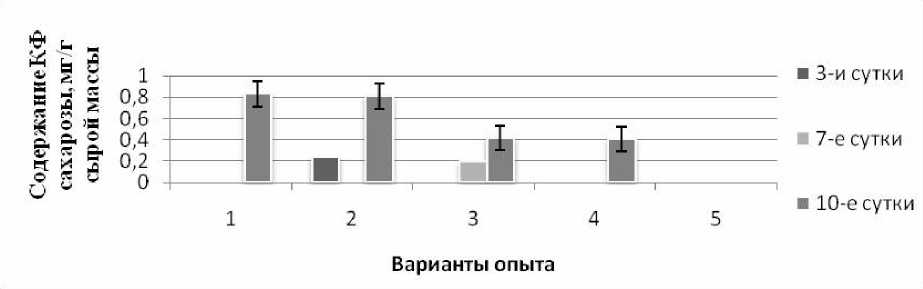
Рис. 3. Динамика содержания КФ сахарозы в проростках кукурузы при кадмиевом стрессе в условиях предобработки повышенной температурой и ИУК, а также без нее
Резюмируя полученные нами данные, следует отметить, что у проростков кукурузы при кадмиевом стрессе в условиях наличия или отсутствия теплового закаливания формируются два фонда растворимых сахаров: свободный (СФ) и компар-тментированный (КФ). Можно предположить, что при кратковременном воздействии кадмия (стадия стресс-реакции) оба фонда растворимых сахаров обеспечивают сочетание повышенного уровня жизнедеятельности и устойчивости, а при более длительном (стадия адаптации) – участвуют в формировании металлоустойчивости. Очевидно, КФ сахарозы – это тот фонд, который может быть востребован клеткой в случае необходимости, например, при адаптации к токсическому действию кадмия.аким образом, изменение компартмента-ции глюкозы и сахарозы является условием реализации их защитных функций растений и формирования термо- и металлоустойчивости, которые за- висят не столько от количества растворимых сахаров в клетках, сколько от их способности к мета-болизации.
На перераспределение моно- и диформ сахаров, а также изменение их компартментации у растений при стрессе указывалось и в работах других авторов [Филатова, Кусакина, Орлова, 2009].
Список литературы Динамика кадмиевой устойчивости проростков кукурузы и содержание в них растворимых углеводов в условиях наличия или отсутствия теплового закаливания
- Большой практикум «Биохимия»: лабораторные работы / сост. М.Г. Кусакина, В.И. Суворов, Л.А. Чудинова. Пермь, 2012. С. 26-35.
- Дерябин А.Н. и др. Влияние сахаров на развитие окислительного стресса, вызванного гипотермией (на примере растений картофеля, экспрессирующих ген инвертазы дрожжей) // Физиология растений. 2007. Т. 54, № 1. С. 39-46.
- Карпец Ю.В., Колупаев Ю.Е. Ответ растений на гипертермию: молекулярно-клеточные аспекты // Вестник Харьковского национального аграрного университета. Серия биология. 2009. Вып. 1 (16). С. 19-38.
- Кафи М., Стюарт В.С., Борланд А.М. Содержание углеводов и пролина в листьях, корнях и апексах пшеницы, устойчивых и чувствительных к засолению // Физиология растений. 2003. Т. 50, № 2. С. 174-182.
- Колупаев Ю.Е., Трунова Т.И. Особенности метаболизма и защитные функции углеводов растений в условиях стрессов // Физиология и биохимия культурных растений. 1992. Т. 24, № 6. С. 523-533.
- Кузнецов Вл.В., Дмитриева Г.А. Физиология растений. М.: Высш. шк., 2005. 736 с.
- Синькевич М.С. и др. Динамика активности инвертаз и содержания сахаров при адаптации растений картофеля к гипотермии // Физиология растений. 2008. Т.55, № 4. С. 501-506.
- Титов А.Ф., Казнина Н.М., Таланова В.В. Тяжелые металлы и растения. Петрозаводск, 2014. 194 с.
- Титов А.Ф., Таланова В.В. Устойчивость растений и фитогормоны. Петрозаводск, 2009. 206 с.
- Филатова Л.А., Кусакина М.Г., Орлова Н.В. Влияние засоления на углеводный обмен Phaseolus vulgaris // Вестник Пермского университета. 2009. Вып. 10. Биология. С. 157-159.
- Ходос В.Н. Роль компартментов метаболитов в процессах регуляции и адаптации метаболизма в растительных клетках. Киев: Наук. думка, 1975. 158 с.
- Холодова В.П. Компартментация сахаров в тканях растений // Рост растений. Первичные механизмы. М.: Наука, 1978. C. 253-277.
- Чудинова Л.А., Суворов В.И. Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений // Вестник Пермского ун-та. Сер. Биология. 2011. Вып. 1. С. 17-20.
- Чудинова Л.А., Чернышева Ю.Д. Динамика содержания растворимых сахаров в проростках овса в условиях раздельного и комбинированного действия повышенных температур и ацетата кадмия // История и методология физиолого-биохимических и почвенных исследований: сб. науч. тр. по материалам науч. конф. Пермь, 2017. С. 48-51.
