Динамика количества чувствительных и моторных нейронов, участвующих в регенерации седалищного нерва крысы после его перерезки

Автор: Тимофеева Л.Б., Благова Н.В., Величанская А.Г., Ермолин И.Л.
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 1 т.19, 2011 года.
Бесплатный доступ
В настоящем исследовании проведён коли- чественный анализ популяций и размерных групп чувствительных и моторных нейронов, принимаю- щих участие в регенерации седалищного нерва от 30 до 300 суток после его перерезки.
Чувствительные нейроны, моторные нейроны, регенерация, седалищный нерв
Короткий адрес: https://sciup.org/143176983
IDR: 143176983
Quantitative analysis of sensory and motor neurons related to the rat sciatic nerve regeneration after its cutting
In the present investigation we have made the quantitative analysis of cell body sizes of sensory and motor neurons populations, related to the sciatic nerve regeneration from 30 to 300 postoperative days.
Текст научной статьи Динамика количества чувствительных и моторных нейронов, участвующих в регенерации седалищного нерва крысы после его перерезки
Введение. Одним из актуальных вопросов в современной медицине является вопрос о возможностях регенерации периферических нервов. Повреждения нервного ствола вызывают расстройства движений, чувствительности и вегетативно — трофические нарушения. Эффективность различных методов лечения остаётся недостаточной. Во многом это связано с недостатком сведений о динамике регенерации повреждённого нерва. Успех восстановления периферического нерва в определённой степени определяется выживаемостью после травмы нейронов, задействованных в его формировании. При этом сведения о динамике количества регенерирующих нейронов при повреждёнии нерва имеют принципиальное значение для его восстановления. Существует большое количество исследований, оценивающих выживаемость нейронов после повреждения седалищного нерва [1—5], но в них гибель нейронов оценивается по отношению к общему количеству нейронов в спинномозговом узле (СМУ) или в сегменте спинного мозга (ССМ). Однако известно, что в формировании седалищного нерва участвуют не все клетки СМУ и ССМ, а лишь определённые популяции нейронов [6-9]. Устойчивость клеток к повреждению в этих популяциях оценивалась немногими авторами [10-11]. Основой успешной регенерации периферических нервов является прорастание нервных волокон из проксимальной культи в дистальную. Известно, что только часть из выживающих нейронов данной популяции способны преодолеть место травмы [12]. Следует так же отметить, что работы, связанные с оценкой динамики количества нейронов, прорастающих в дистальную культю, а также их размерных групп, в литературе отсутствуют.
Цель исследования – изучить общее количество и соотношение размерных групп популяций чувствительных и моторных нейронов, способных преодолеть соединительнотканный рубец через 30, 90, 150 и 300 суток после перерезки седалищного нерва.
Материалы и методы. Исследования проведены на 27 белых беспородных самцах крыс массой 200-250г. Животные были разделены на контрольную (n=7) и экспериментальную (n=20) группы.
Для выявления популяции нейронов, формирующих седалищный нерв, был применён метод ретроградного маркирования нейронов флуоресцентным красителем Mini-Ruby — dextran, tetramethylrhodamine and biotin 10,000 MW, lysine fixable (Molecular Probes). Для этого у контрольных животных выделяли седалищный нерв, пересекали его на уровне верхней трети бедра и апплицировали красителем. После стандартной обработки материала [13] на криостате были приготовлены продольно ориентированные серийные срезы толщиной 25 мкм.
У экспериментальных животных маркирование нейронов производилось через 30, 90, 150 и 300 суток после перерезки седалищного нерва на уровне верхней трети бедра. Флуоресцентный краситель вводился в дистальную культю нерва. Материалом для гистологического исследования послужили СМУ L4, L5 и L6 и ССМ L4, L5 и L6, поскольку ранее проведённые исследования пока-
Таблица 1.
Динамика количества чувствительных и моторных нейронов, формирующих седалищный нерв в норме и после его перерезки.
Результаты и обсуждение. Результаты подсчёта маркированных нейронов показали, что через 30-ть суток после перерезки седалищного нерва в реиннервации его дистальной культи участвуют 20,47 % нейронов СМУ от их общего количества в популяции в норме. Количество моторных нейронов к 30-м суткам составило 23,36 % от нормы (табл.1).
Ранее нами прово- проведено их подразделение на большие (75 мкм > d > 35 мкм) и мелкие (35 мкм > d) нейроны в соответствии с литературными данными [15-16]. Анализ размерных групп моторных нейронов показал, что в регенерации седалищного нерва участвуют 13,38 % больших нейронов и 173,38 % мелких нейронов от их количества в норме (рис.2).
Причиной увеличения количества мелких мотонейронов, по-видимому, являются значительные дегенеративные изменения перикарионов крупных мотонейронов после повреждения их аксонов, вследствие чего последние переходят в категорию мелких [17].
К 90-м суткам общее количество чувствительных нейронов по сравнению с предыдущим сроком существенно не изменилось и составило 19,25 % от количества нейронов в норме. Существуют данные о регенерации ветвей спинномозгового нерва на уровне Т13 [18], согласно которым в популяциях, участвующих в формировании данных ветвей на сроке 90 суток место травмы дилось подразделение популяции чувствительных нейронов данной популяции в зависимости от размера перикариона на большие (d > 35 мкм), средние (35 мкм > d > 20 мкм) и мелкие (20 мкм > d) [14]. Анализ размерных групп чувствительных нейронов показал, что на 30-е сутки в регенерации дистальной культи седалищного нерва принимают участие 5,58 % больших нейронов; 29,22 % средних нейронов; 11,68 % мелких нейронов от их количества в норме (рис.1).
На основании измерений профильных тел моторных нейронов было
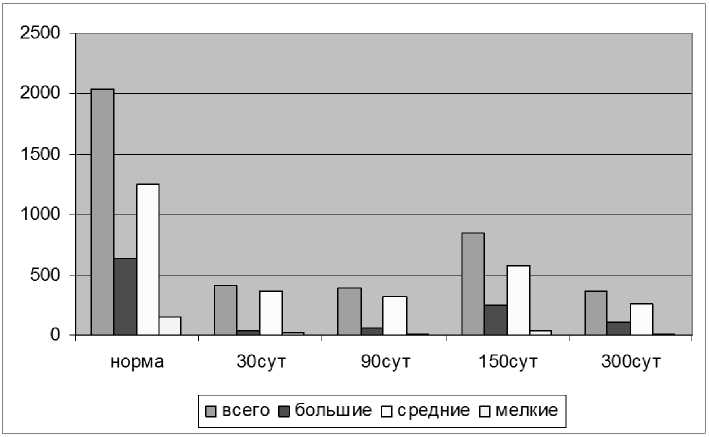
Рис. 1. Изменение количества и размерных групп чувствительных нейронов, дендриты которых проросли в дистальную культю седалищного нерва
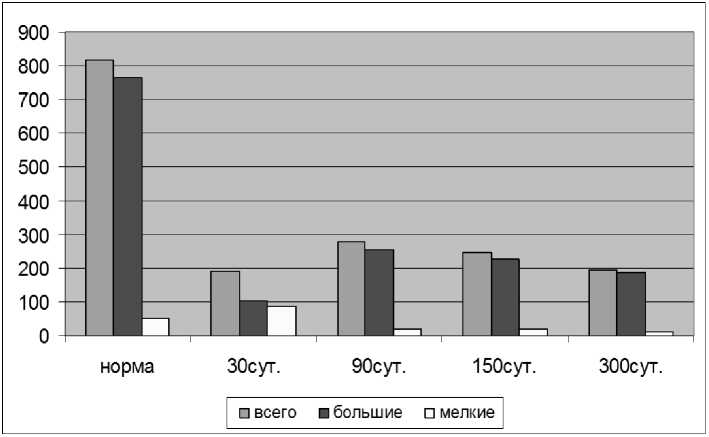
Рис. 2. Изменение количества и размерных групп моторных нейронов, аксоны которых проросли в дистальную культю седалищного нерва
составило 34,01 % от нормы. По сравнению с 30-ми сутками увеличение их количества статистически недостоверно. Количество больших моторных нейронов увеличилось в 2 раза по сравнению с 30ми сутками и составило 33,86 % от их количества в норме, а количество малых нейронов уменьшилось в 4 раза и составило 42,04 % от их количества в норме (рис.2).
Через 150 суток общее количество чувствительных нейронов, преодолевших область травмы, значительно увеличилось и составило 41,72 % от количества нейронов в норме. Увеличение количества преодолевают дендриты только 4 % нейронов. По-видимому, данное расхождение результатов связано с тем, что в работе И. Л. Ермолина повреждение нерва производилось значительно ближе к телам нейронов. Так же нами было выявлено, что на 90-е сутки в размерных группах произошло незначительное увеличение количества больших нейронов по сравнению с предыдущим сроком. Их количество составило 9,18 % от количества нейронов в норме. Количество средних и мелких нейронов незначительно уменьшилось и составило 25,69 % и 9,13 % соответственно (рис.1).
Общее количество моторных нейронов, проросших в дистальную культю, на данном сроке произошло во всех размерных группах нейронов. Количество больших нейронов по сравнению с предыдущим сроком увеличилось более чем в 4 раза и составило 37,92 % от нормы. Количество средних и мелких нейронов так же существенно увеличилось и составило соответственно 46,46 % и 19,75 % от их количества в норме (рис.1).
Общее количество регенерировавших моторных нейронов через 150 суток после перерезки нерва составило 30,26 % от нормы. Количество больших и мелких моторных нейронов по сравнению с предыдущим сроком практически не изменилось и составило 33,46 % и 42,86 % от их количества в норме соответственно (рис.2).
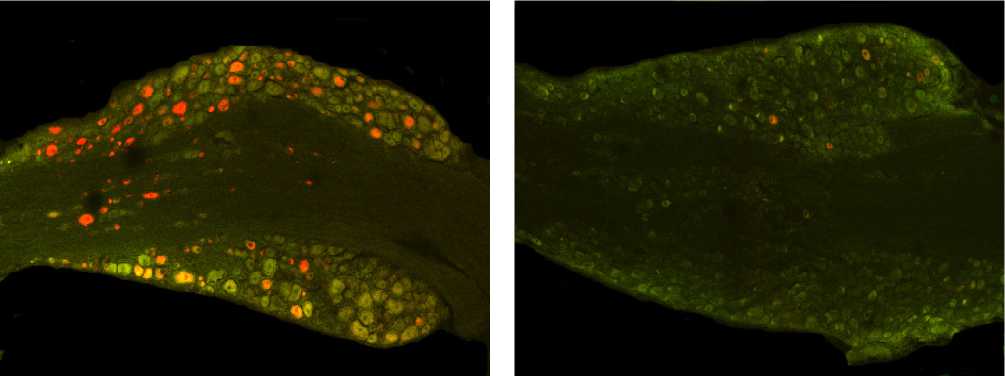
Рис. 3. Маркированные нейроны СМУ L4 в норме (а) и через 300 суток (б) после перерезки седалищного нерва. Окраска Mini-Ruby. Ув.100.
Таблица 2.
Процентное соотношение чувствительных нейронов в размерных группах в норме и после перерезки седалищного нерва.
|
Срок, сутки |
Количество нейронов в размерной группе, % |
|||
|
Большие |
Средние |
Мелкие |
Общее количество |
|
|
Норма |
31,30 |
61,00 |
7,7 |
100 |
|
30 |
8,53 |
87,08 |
4,39 |
100 |
|
90 |
20,40 |
75,40 |
4,2 |
100 |
|
150 |
31,09 |
65,51 |
3,65 |
100 |
|
300 |
33,78 |
63,41 |
2,8 |
100 |
К 300-м суткам общее количество чувствительных нейронов уменьшилось до 17,94 % от нормы (рис.1,3). Количество больших и средних нейронов по сравнению с предыдущим сроком уменьшилось более чем наполовину и составило соответственно 16,12 % и 20,66 % от их количества в норме, количество же мелких нейронов уменьшилось более чем в пять раз и составило 3,82 % от нормы.
Общее количество регенерирующих моторных нейронов на данном сроке составило 24,03 % от нормы (рис.1,4). Количество больших нейронов по сравнению с предыдущим сроком уменьшилось на треть и составило 24,30 % от их количества в норме, а количество малых нейронов уменьшилось в два раза и составило 20,11 % от нормы.
Таким образом, общее количество регенерирующих чувствительных нейронов на сроках 30, 90 и 300 суток после операции достоверных отличий не имеет и составляет от 20,47 % до 17,94 %. При этом по мере увеличения послеоперационного срока происходит увеличение количества больших и уменьшение количества средних и мелких нейронов в популяции. К 300 суткам данные изменения в размерных группах больших и средних нейронов достигают статистически достоверных значений по сравнению с 30-ми сутками. Достоверность уменьшения количества мелких нейронов установить не удалось ввиду их малочисленности. Следует отметить значительное увеличение общего количества нейронов через 150 суток до 41,72 % от нормы по отношению к предыдущим срокам (рис.1), которое затронуло все размерные группы, но в большей степени большие нейроны. При оценке соотношения размерных групп нейронов в популяции СМУ выявлено его значительное изменение к 30-м суткам (табл. 2). К 300 суткам процентное соотношение размерных групп в популяции приближается к норме.
Общее количество моторных нейронов, преодолевающих область травмы, и участвующих в посттравматической регенерации седалищного нерва, достоверно не изменяется на всех сроках наблюдения и составляет от 23,36 % до 34,01 % от популяции в норме. Количественные показатели на 30-е и 300-е сутки отличаются менее чем на 1 %.
При оценке соотношения размерных групп нейронов в популяции ССМ выявлено, что на 30-е сутки эксперимента количество мелких мотонейронов достигает 46,33 % от выявленной популяции. В норме этот показатель составляет 6,35 %. Количество больших нейронов на 30-е сутки
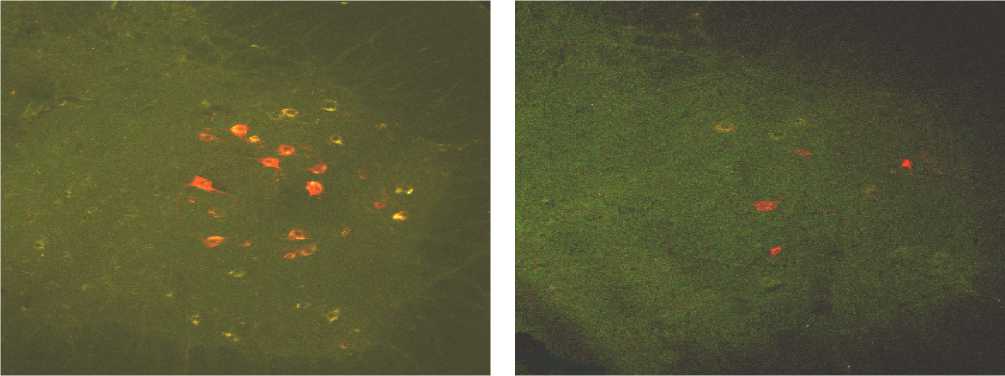
Рис. 4. Маркированные нейроны спинного мозга на уровне L4 в норме (а) и через 300 суток (б) после перерезки седалищного нерва. Окраска Mini-Ruby. Ув.100.
Таблица 3.
Процентное соотношение моторных нейронов в размерных группах в норме и после перерезки седалищного нерва.
Таким образом, после перерезки седалищного нерва к 30-м суткам область травмы преодолевает лишь пятая часть как чувствительных, так и моторных нейронов от общего количества нейронов популяции. Изменение общего количества нейронов популяций СМУ и ССМ L4 – L6 на остальных экспериментальных сроках по сравнению с 30-ми сутками не имеет существенных колебаний. Исключением является значительное увеличение количества нейронов в популяции СМУ на 150-е сутки. К 300-м суткам показатели по чувствительным и моторным нейронам имеют тенденцию к уменьшению.
Список литературы Динамика количества чувствительных и моторных нейронов, участвующих в регенерации седалищного нерва крысы после его перерезки
- Челышев Ю. А. Выживание и фенотипическая характеристика аксотомированных нейронов спинальных ганглиев / Ю. А. Челышев, И. С. Рагинов, Д. С. Гусева, Р. Ф. Масгутов // Морфология. - 2004. - т. 125, № 3. - С.4549.
- Tandrup T. Delayed loss of small dorsal root ganglion cells after transection of the rat sciatic nerve /T. Tandrup, C. Woolf, R. Coggeshall // J Comp Neurol. - 2000. - Vol. 422, № 2. - P. 172 - 180. 3. McKay Hart А. Primary sensory neurons and satellite cells after peripheral axotomy in the adult rat: timecourse of cell death and elimination / A. McKay
- Hart, T. Brannstrom, M. Wiberg et al. // Exp Brain Res. - 2002. - Vol.142, № 3. - P.308 - 318.
- Groves M. J. Profile of adult rat sensory neuron loss, apoptpsis and replacement after sciatic nerve crush / M. J. Groves, A. Schanzer, A. J. Simpson et al. // J. Neurocytol. - 2003. - Vol. 32, № 2. - P. 113 - 122.
- Terao S. Upper Motor Neuron Lesions in Stroke Patients Do Not Induce Anterograde Transneuronal Degeneration in Spinal Anterior Horn Cells / S. Terao, M. Li, Y. Hashizume et al. // American Heart Association. - 1997. - Vol. 28. - Р. 25532556.
- Swett J. E. Motoneurons of the rat sciatic nerve / J. E. Swett, R. P. Wikholm, R. H. Blanks et al. // Exp Neurol. - 1986. - Vol.93, № 1. - P. 227 - 252.
- Swett J. E. Sensory neurons of the rat sciatic nerve / J. E. Swett, Y. Torigoe, V. R. Elie // Exp Neurol. - 1991. - Vol.114. - P. 182 - 103.
- Благова Н. В. Особенности распределения популяции мотонейронов спинного мозга, уча ствующих в образовании седалищного нерва взрослых крыс / Н. В. Благова, И. Л. Е рмолин // Журнал теоретической и практической медицины. - 2010. - т.8. - С. 7677.
- Тимофеева Л. Б. А фференты седалищного не рва взрослой крысы / Л. Б. Т имофеева, И. Л. Е р молин // Журнал теоретической и практической медицины. - 2010. - т.8. - С. 219220.
- Himes B. Death of some dorsal root ganglion neurons and plasticity of others following sciatic nerve section in adult and neonatal rats / B. Himes, A. Tessler // The Journal of Comparative Neurology. - 1989. - Vol. 284, № 2. - P. 215230.
- ValeroCabre А. Peripheral and spinal motor reorganization after nerve injury and repair / A. ValeroCabre, К. Tsironis, Е. Skouras et al. // J. Neurotrauma. - 2004. - Vol.21, № 1. - Р. 95108.
- Тимофеева Л. Б. Морфология популяций чув ствительных и моторных нейронов, формирующих седалищный нерв в норме и при его перерезке у взрослых крыс / Л. Б. Т имофеева, Н. В. Благова, А. Г. Величанская и др. // Современные техноло гии в медицине. - 2010. - № 3. - C.1823.
- Ермолин И. Л. Деафферентация сенсорных нейронов при перерезке периферического нерва у взрослой крысы / И. Л. Е рмолин // Морфологи ческие ведомости. - 2005. - № 34. - С. 2729.
- Тимофеева Л. Б. Соотношение размерных групп в популяции афферентных нейронов после перерезки седалищного нерва / Л. Б. Т имофеева // Морфология. - 2010. - Т. 137, № 4. - С. 188189.
- Brushart T. M. Alteration in connections between muscle and anterior horn motoneurons after peripheral nerve repair / T. M. Brushart, M. M. Mesulam // Science. - 1980. - Vol. 9, № 208. - P. 603605.
- Быков В. Л. Частная гистология человека (краткий обзорный курс)/ В. Л. Быков. - СПб.:СОТИС, 1999. - 300с.
- Novikov L. N. Brain-derived neurotrophic factor promotes survival and blocks nitric oxide synthase expression in adult rat spinal motoneurons after ventral root avulsion / L. N. Novikov, L. N. Novikovа, J. - O. Kellerth // Neuroscience Letters. - 1995. - Vol. - P.45-48.
- Ермолин И. Л. Количественная оценка маркированных нейронов спинномозгового узла Т13 при регенерации периферического нерва у взрослых крыс (сообщение III) // Нижегородский медицинский журнал. - 2006. - № 2. - С. 24-29.
