Динамика растительного сообщества и микробиома хроносерий посттехногенных почв в известняковом карьере в условиях рекультивации

Автор: Дмитракова Я.А., Абакумов Е.В., Першина Елизавета Владимировна, Иванова Екатерина Андреевна, Андронов Евгений Евгеньевич
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Растение и почва
Статья в выпуске: 3 т.53, 2018 года.
Бесплатный доступ
Посттехногенные экосистемы - удобная модель для изучения процессов начального почвообразования и восстановления экосистем. Их можно рассматривать как естественные модели восстановления микробных сообществ во времени (временные ряды и хроносерии). Исследование разновозрастных стадий почвообразования на отвалах карьеров разного типа позволяет получить фактическую информацию о скорости и направленности почвообразования в разнообразных субстратно-фитоценотических комбинациях. Среди карьеров нерудных полезных ископаемых особенное место в структуре горной добыче на Северо-Западе занимают участки по добыче известковых пород, приуроченные к Ижорской возвышенности. Целью настоящей работы было изучение динамики фитоценозов и почвенного микробиома в процессе сукцессии на отвалах различного возраста при естественном самозарастании и рекультивации известнякового карьера. Исследование проводили на карьере Печурки (Сланцевский р-н, Ленинградская обл.) весной и летом 2016 года. Проективное покрытие растительности и видовой состав оценивали на каждом экотопе карьера...
Первичная сукцессия растительности и микроорганизмов, известняковые карьеры, педогенез, метагеном, рекультивация
Короткий адрес: https://sciup.org/142216558
IDR: 142216558 | УДК: 631.48:631.461:577.2 | DOI: 10.15389/agrobiology.2018.3.557rus
Dynamics of the plant community and microbiom of chrono-series of post-technological soil in limestone quarry in the conditions of recultivation
Post-technogenic ecosystems represent informative natural models of initial pedogenesis and restoration under the abandoned and reclamation practices. These soils can be considered as natural experiments on restoration of microbial communities within the age row (time series or chronoseries). Investigation of different aged stages of soil formation on the spoil banks of the quarries give a possibility to obtain initial data on the rate and trends of the pedogenesis in various combinations of substrates and phytocenosis. Among the quarries of mineral substrates in North-West region, particular place is devoted to quarries for lime stone exploitation located on Izhora upland. The purpose of this study conducted in one of the largest limestone quarries of Leningrad region was to examine succession of plant and microbial communities. Species composition and vegetation cover were estimated for different plant communities within each ecotype of quarry. Also at the each plot the following characteristics of soil were measured: pH; organic carbon; soil basal respiration and substrate induced respiration; texture; CO2 of carbonates and moisture content...
Текст научной статьи Динамика растительного сообщества и микробиома хроносерий посттехногенных почв в известняковом карьере в условиях рекультивации
Увеличение территорий нарушенных земель, происходящее в результате разработки и добычи полезных ископаемых, представляет собой осо-
Работа выполнена при поддержке гранта РНФ ¹ 17-16-01030 «Динамика почвенной биоты в хроносериях посттехногенных ландшафтов: анализ почвенно-экологической эффективности процессов восстанов- ления экосистем».
бую экологическую проблему. Как правило, антропогенные ландшафты выводятся из оборота лесных и сельскохозяйственных земель, а процессы восстановления почвенно-растительного покрова идут крайне медленно (1). Изучение почвообразования в условиях техногенеза служит теоретической базой для рекультивации нарушенных земель и восстановления ландшафтов (2, 3). Дерново-карбонатные почвы, формирующиеся на Ижорском плато, характеризуются высоким плодородием и дренированностью, благодаря чему эти территории долгое время активно использовались в сельскохозяйственном производстве (4). Высокая карбонатность отвальных пород обусловливает потенциальное плодородие известковых отвалов, однако специфика сложения карьерно-отвальных комплексов, а также относительно низкое содержание элементов питания и неблагоприятные физические свойства субстрата создают трудности при организации работ по рекультивации этих территорий (5). При выборе сельскохозяйственного направления рекультивации довольно трудоемкой и дорогостоящей задачей становится выравнивание поверхности (3). Однако сложный рельеф, формирующийся в процессе разработки карьеров, обеспечивает высокое разнообразие экологических ниш, благоприятных для существования большого количества видов растений и животных, включая те, которые имеют высокую природоохранную ценность. Так, в различных условиях и географических зонах выявлено, что заброшенные карьеры могут увеличивать биоразнообразие, выступая в качестве рефугиумов для редких видов растений и животных (6, 7).
В мировой практике рекультивации особое внимание уделяется использованию регенерационных возможностей самих природных экосистем, где главную роль играют биотические факторы почвообразовательного процесса — растительный покров и сопряженный с ним почвенный микробиом (8, 9). Именно с деятельностью микроорганизмов напрямую связано плодородие и обеспечение экосистемных функций почвы, включающих сбалансированность цикла биогенных элементов, а также экологическую адаптацию растений к стрессовым условиям (10). Кроме того, высокий адаптивный потенциал почвенных микроорганизмов, способных в короткие сроки приспосабливаться к любым изменениям среды и осваивать все доступные экологические ниши, позволяет использовать структуру микробиома в качестве одного из наиболее чувствительных экологических индикаторов, маркирующих разные стадии почвовосстановления (11).
Спонтанная сукцессия растительных сообществ на карьерах с различными субстратами изучена достаточно подробно. Значительная часть отечественных исследований посвящена разнообразным аспектам сукцессии растительного покрова (12-14), довольно полно проанализированы процессы педогенеза (15, 16). Однако комплексные наблюдения, посвященные восстановлению почвенно-растительного покрова и связи этих компонентов, остаются малочисленными. Большинство зарубежных работ, напротив, сфокусированы на сопряженном развитии почвенно-растительного покрова, но не вследствие самозарастания, а после посадок определенных видов растений (17-20). Традиционно в работах по рекультивации земель основное внимание уделялось исследованию растительного покрова и перераспределению почвенного органического вещества (9, 15, 16). Значение микробиома в процессах восстановления экосистем долгое время оставалось недооцененным. В последнее десятилетие изучению микробиома рекультивируемых почв посвящены многие публикации (21-23), содержащие ценные сведения о темпах восстановления биомассы и активности микробных сообществ в хроносериях почв техногенных ландшафтов (24-26). Показано, что структура и состав микробиомов определяются совокупностью физических и агрохимических параметров (запасы почвенного органического вещества, общего азота, рН, катионообменная способность почв), значения которых существенно выше в рекультивируемых вариантах. Большое внимание уделяется использованию микробных сообществ в качестве индикаторов различных стадий почвовосстановления техногенных ландшафтов (27-29). При этом разнообразие и структура микробиомов в нарушенных и рекультивированных почвах все еще недостаточно изучены. Только с появлением метагеномных технологий стало возможным эффективно анализировать и интерпретировать разнообразие почвенной микробиоты. Вовлечение в исследование всего многообразия почвенного микробиома создает перспективы для разработки качественно новых систем для ускорения и оптимизации рекультивационных мероприятий на нарушенных территориях. В последнее время уделяется внимание вопросам создания микробных препаратов для улучшения качества почв и грунтов (30, 31).
Впервые на территории одного из самых крупных известняковых карьеров в Ленинградской области мы провели комплексное исследование трех главных компонентов экосистемы (почва, растительное сообщество и микробиом) в хроносериях различных экотопов в условиях рекультивации и самозарастания. Установлена таксономическая структура микробиома инициальных почв как для карьеров Северо-Запада, так и для инициальных почв России в целом.
Целью работы было изучение динамики фитоценозов и почвенного микробиома в процессе сукцессии на отвалах разного возраста при естественном самозарастании и рекультивации известнякового карьера.
Методика. Исследование проводили на известняковом карьере Печурки (Сланцевский р-н, Ленинградская обл.) весной и летом 2016 года. Добыча известняка здесь была прекращена в 2014 году. В первую очередь определили основные экотопы карьера, соответствующие элементарному почвенному ареалу. В пределах каждого экотопа были заложены 12 пробных площадок размером 25½25 м. На площадках оценивали общее проективное покрытие растительности и видовой состав высших сосудистых растений, напочвенных мхов и лишайников. Также в пределах каждого участка были заложены почвенные разрезы. На каждой из прикопок выполняли полевые описания почв, из почвенных горизонтов отбирали образцы (по 700 г) для выполнения лабораторных анализов.
Определяли следующие показатели: субстрат-индуцированное дыхание (СИД) почвенных образцов (32); базальное дыхание по методике для СИД (32), но в не обогащенной субстратом почве; содержание органического углерода по бихроматной окисляемости (метод И.В. Тюрина); рН водной суспензии и солевой рН (1:2,5 почва:раствор); обменную и гидролитическую кислотность; содержание СО2 карбонатов ацидиметрически (33); гигроскопическую и максимальную гигроскопическую влажность, а также полную влагоемкость (водовместимость) и наименьшую влагоем-кость гравиметрически (34); плотность почвы и твердой фазы почв, структурность и каменистость почвы — методом сухого просеивания; гранулометрический состав — пипет-методом Качинского с прирофосфатной пептизацией микроагрегатов (34); фракционно-групповой состав гумуса — по схеме И.В. Тюрина, модифицированной В.В. Пономаревой с соавт. (35).
Количество углерода микробной биомассы рассчитывали по формуле J.P.E. Anderson с соавт. (36): Смик (мкг С/г почвы) = СИД (мкл CО2/г почвы•ч-1) ½ 40,04 + 0,37, где СИД — субстрат-индуцированное дыхание. Микробный метаболический коэффициент (удельное дыхание микробной биомассы) находили как отношение величины базального дыхания (БД) к показателю Смик qСО2 (мкг СО2-С/мг Смик • ч-1) = БД/Смик.
По составленным для каждого участка спискам видов высших растений рассчитывали коэффициент видового сходства (коэффициент Съе-ренсона-Чекановского): K = 2c/(a + b), где a — число видов на первой площадке, b — число видов на второй площадке, с — число общих видов для первой и второй площадок. Также для каждого участка определяли наиболее типичные коэффициенты биоразнообразия. Индекс Симпсона (С) вычисляли по формуле: С = 1 - Z (n i /N)2, где n — оценка значимости каждого вида (проективное покрытие), N — сумма оценок значимости. Индекс Шенона вычисляли согласно формуле: H = - £ n i /N х log(n i /N).
Метод прямого отбора (forward selection) с двумя останавливающими критериями (37) для сокращения ошибки I типа применяли для оценки влияния факторов окружающей среды на структуру фитоценозов. Используя только значимые переменные (P < 0,05), выполняли канонический анализ соответствий (canonical correspondence analysis, CCA), предложенный C.J.F. Ter Braak (38), чтобы узнать, как факторы окружающей среды действуют на распределение растительных сообществ.
Образцы почвы отбирали с глубины 8-10 см на площадках ¹¹ 19. Из образов (0,2 г) выделяли ДНК с помощью набора PowerSoil® DNA Isolation Kit («Mo Bio Laboratories, Inc.», США) согласно предписаниям производителя. Очищенные препараты ДНК (10-15 нг) служили матрицами в реакции ПЦР (температурный профиль: 30 с при 95 °C, 30 с при 50 °C, 30 с при 72 °C; всего 30 циклов) с использованием ДНК-полимеразы En-cyclo («Евроген», Россия) и универсальных праймеров к вариабельному участку v4 гена 16S рРНК: F515 (GTGCCAGCMGCCGCGGTAA) и R806 (GGACTACVSGGGTATCTAAT). В праймеры вводили олигонуклеотидные идентификаторы для каждого образца и служебные последовательности, необходимые для пиросеквенирования по протоколу «Roche» (Швейцария). Подготовку проб и секвенирование выполняли на приборе GS Junior («Roche», Швейцария) в соответствии с рекомендациями производителя. Демультиплексирование, проверку качества последовательностей 16S рРНК, выделение ОТЕ (операционных единиц анализа, аналог вида), нормализацию образцов, таксономическую идентификацию ОТЕ, подсчет индексов а- и p-разнообразия осуществляли в пакете ПО QIIME с использованием параметров, установленных по умолчанию (39, 40). Библиотеки фрагментов гена 16S рРНК были нормализованы по числу последовательностей в библиотеке наименьшего размера.
Различия в частотах микробных таксонов между исследуемыми образцами определяли посредством проведения точного теста Фишера с поправкой на множественные сравнения по процедуре Бенджамини-Хох-берга на 5 % уровне значимости. Для того чтобы объяснить изменчивость видового состава микробных сообществ под влиянием фактора среды, применяли метод главных компонент (PCoA). Вычисления проводили в программах Statistica 7 («StatSoft, Inc.», США) и Microsoft Excel.
Результаты . С 1970 года в известняковом карьере Печурки начались работы по рекультивации, в процессе которых на некоторых участках в разное время высаживали саженцы сосны. При этом большую часть карьера оставили под самозарастание. Карьер характеризовался сложным внутренним рельефом, что способствовало формированию относительно высокого разнообразия почв. Большинство факторов среды были оптимальными для развития растительности (табл.), за исключением высокой плотности и сильной каменистости на скальных днищах карьера. Для всех участков было характерно крайне неоднородное распределение фракций по профилю и значительное количество скелетного крупнообломочного ма560
териала при относительно низком содержании мелкозема. В целом, результаты мезоморфологических исследований свидетельствовали о довольно высоких темпах педогенного преобразования субстрата. Основные процессы преобразования минеральной части почв — химическое, биохимическое и физическое выветривание карбонатных пород. Интенсивному выветриванию обломков известняков способствовало высокое содержание мелкозема (исключением были лишь каменистые днища карьера). Это, в свою очередь, повышало влагоемкость эмбриоземов и их плодородие, тем самым обеспечивалось активное развитие растительных сообществ, продуцирующих органическое вещество. Интенсивная декарбонатизация выражалась в значительном снижении рН гумусовых горизонтов по сравнению с породой.
Характеристика почв, формирующихся в известняковом карьере Печурки (Слан-цевский р-н, Ленинградская обл., 2016 год)
|
А |
] Б |
1 В 1 |
Г |
1 Д 1 |
Е 1 |
Ж 1 |
З 1 |
И |
1 К |
1 Л |
1 М |
|
1 |
O |
0-4 |
5,02 |
0,00 |
8,16 |
0,85 |
0,07 |
0,07 |
3,225 |
0,021 |
13,1 |
|
AY |
4-33 |
4,40 |
0,06 |
6,79 |
0,03 |
0,03 |
1,695 |
0,018 |
|||
|
C |
33-48 |
5,72 |
0,10 |
1,19 |
0,02 |
0,02 |
1,186 |
0,017 |
|||
|
[C] |
48+ |
7,45 |
0,17 |
0,86 |
0,03 |
0,03 |
1,491 |
0,017 |
|||
|
2 |
O |
0-3 |
5,26 |
0,00 |
1,69 |
0,76 |
0,11 |
0,12 |
5,060 |
0,022 |
30,5 |
|
BF |
3-13 |
6,35 |
0,24 |
5,73 |
0,04 |
0,04 |
2,103 |
0,019 |
|||
|
C |
13+ |
6,55 |
0,08 |
2,05 |
0,04 |
0,04 |
1,797 |
0,020 |
|||
|
3 |
O |
0-7 |
6,36 |
0,10 |
3,27 |
0,78 |
0,04 |
0,05 |
2,205 |
0,018 |
6,9 |
|
AY |
7-15 |
5,86 |
0,10 |
7,85 |
0,02 |
0,03 |
1,491 |
0,015 |
|||
|
G |
15-36 |
6,43 |
0,07 |
0,65 |
0,06 |
0,07 |
3,021 |
0,020 |
|||
|
G ox |
36-45 |
6,12 |
0,08 |
1,94 |
0,04 |
0,04 |
2,001 |
0,019 |
|||
|
C |
45 |
6,50 |
0,09 |
5,45 |
0,02 |
0,02 |
0,982 |
0,016 |
|||
|
4 |
AY |
0-26 |
5,22 |
0,21 |
3,94 |
0,91 |
0,04 |
0,05 |
2,205 |
0,018 |
29,5 |
|
AC |
26+ |
6,34 |
0,16 |
6,32 |
0,02 |
0,02 |
0,982 |
0,016 |
|||
|
5 |
O |
0-13 |
5,64 |
0,00 |
14,4 |
0,65 |
0,04 |
0,05 |
2,205 |
0,020 |
28,0 |
|
AY |
13-25 |
6,05 |
0,35 |
2,13 |
0,06 |
0,07 |
3,021 |
0,021 |
|||
|
C1 |
25-37 |
5,76 |
0,08 |
0,67 |
0,02 |
0,03 |
1,390 |
0,018 |
|||
|
C2 ox |
37+ |
6,10 |
0,52 |
3,33 |
0,03 |
0,04 |
1,797 |
0,017 |
|||
|
6 |
O |
0-4 |
6,57 |
0,00 |
24,81 |
0,06 |
0,08 |
2,817 |
0,022 |
13,8 |
|
|
AY |
4-28 |
6,53 |
0,13 |
13,25 |
0,76 |
0,04 |
0,04 |
2,001 |
0,018 |
||
|
C |
28+ |
5,40 |
0,27 |
4,47 |
0,04 |
0,04 |
1,797 |
0,020 |
|||
|
7 |
O |
0-7 |
5,98 |
0,00 |
8,24 |
0,07 |
0,08 |
3,429 |
0,019 |
4,5 |
|
|
AY |
7-9 |
6,90 |
0,00 |
9,46 |
0,68 |
0,03 |
0,04 |
1,899 |
0,017 |
||
|
C |
9+ |
7,35 |
0,09 |
3,89 |
0,04 |
0,04 |
1,899 |
0,019 |
|||
|
8 |
C |
0-5 |
6,70 |
0,52 |
3,24 |
0,05 |
0,05 |
2,307 |
0,020 |
80,0 |
|
|
9 |
O |
0-3 |
5,96 |
0,00 |
8,80 |
0,57 |
0,07 |
0,07 |
3,327 |
0,021 |
22,4 |
|
AC |
3-14 |
6,70 |
0,36 |
3,75 |
0,04 |
0,04 |
2,103 |
0,019 |
|||
|
10 |
AY |
0-18 |
6,50 |
0,04 |
12,64 |
0,62 |
0,02 |
0,11 |
4,669 |
0,005 |
22,4 |
|
C |
18+ |
6,50 |
0,07 |
12,50 |
0,04 |
0,08 |
3,726 |
0,010 |
|||
|
11 |
AC |
0-3 |
6,00 |
0,91 |
16,00 |
0,03 |
0,08 |
3,621 |
0,009 |
80,0 |
|
|
C |
3+ |
6,30 |
1,03 |
15,58 |
0,02 |
0,09 |
3,831 |
0,005 |
|||
|
12 |
AY |
0-25 |
6,15 |
0,23 |
17,68 |
0,8 |
0,02 |
0,10 |
4,250 |
0,005 |
30,0 |
|
C |
25+ |
6,11 |
0,22 |
10,00 |
0,02 |
0,07 |
3,097 |
0,006 |
|||
|
П р и м е ч а н и е. А |
— участок; Б |
— горизонт; В |
мощность, |
см; Г — |
pH вод; Д — |
CO 2 , %; Е — |
Cорг, %; |
||||
Ж — Сгк/С ф к; З — базальное дыхание, мкг СO2-С/г•ч - 1; И — СИД (субстрат-индуцированное дыхание), мкг СO2-С/г•ч - 1; К — Смик, мкг С/г почвы; Л — qCO2, мкг СO2-С/мг Смик•ч - 1; М — каменистость, %;
Сорг — содержание общего органического углерода, Сгк — содержание углерода гуминовых кислот, С ф к — содержание углерода фульвокислот, Смик — содержание углерода микробной биомассы, qCO2 — метаболический коэффициент. Прочерки означают, что анализ не проводился.
Места для закладки пробных площадок выбирали таким образом, чтобы избежать краевого эффекта (на стыках биоценозов увеличивается плотность и число видов, что может привести к искаженным результатам), то есть предпочтительно в центральной части фитоценоза (в случаях его наличия) или центральной части экотопа (при отсутствии сомкнутого растительного покрова). Расположение пробных площадок представлено на рисунке 1. Ключевыми педогенными процессами на всех участках становились накопление органического вещества и окисление субстрата. Они приводили к формированию А-горизонта с максимальной мощностью и наибольшим содержанием органического вещества в аккумулятивных эко-топах с оптимальными условиями увлажнения и физическими параметрами субстрата. Различия между верхними органогенными горизонтами и ниж-
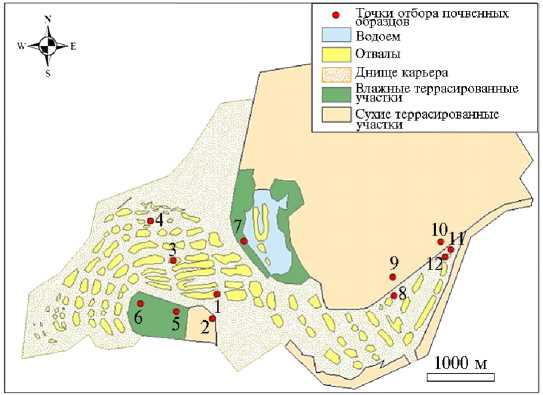
Рис. 1. Схема отбора почвенных образцов в известняковом карьере Печурки: 1 — отвал, редкая растительность (возраст 28 лет), 2 — сухая терраса, мелкий сосняк (16 лет), 3 — отвал, редкая растительность (30 лет), 4 — отвал, редкая растительность (8 лет), 5 — влажная терраса, мелколиственный лес (35 лет), 6 — влажная терраса, мелколиственный лес (35 лет), 7 — влажная терраса, зеленомошный сосняк (35 лет), 8, 11, 12 — основание отвала, редкая растительность (29 лет), 9, 10 — сухая терраса, редкая растительность (29 лет) (Сланцев-ский р-н, Ленинградская обл., 2016 год).
ними росли с увеличением срока зарастания. Первичный субстрат и нижележащие горизонты характеризовались более высокими значениями кислотности (с рН от 6 до 7,45) вследствие большого количества первичных минералов и отсутствия органического вещества. Содержание органического углерода увеличивалось со временем зарастания, тогда как значение рН уменьшалось до 5,02. Наиболее кислая реакция наблюдалась на положительных элементах рельефа (участки 1, 2, 4). Основными доминантами здесь были Populus tremula и Betula pendula .
Преобладали почвы с фульватно-гуматным типом гумуса, что было характерно для района. Повышенная доля гуминовых кислот объясняется карбонатным субстратом почвообразующей породы. Для включения в модель методом прямого отбора выделили пять факторов окружающей среды, оказывающих наибольшее влияние на распределение растительности: гранулометрический состав почвы, каменистость, содержание физической глины, рН водной вы- тяжки и влажность. Эти данные согласуются с исследованиями, проведенными в Канаде (41). Все выделенные нами факторы значительно влияют на распределение растительного сообщества, однако в других источниках, по- мимо них, отмечают огромное значение возраста отвала (41-43).
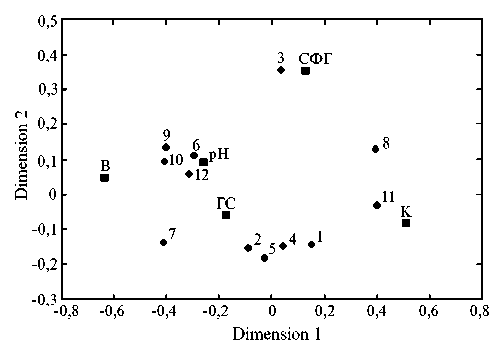
Рис. 2. Результаты канонического анализа соответствий факторов среды и растительных сообществ (неметрические мультиразмерные шкалы). Кружками обозначены растительные сообщества, СФГ — содержание физической глины, В — влажность, ГС — гранулометричекий состав, К — каменистость (анализ выполнен в программе Statistica 7; карьер Печурки, Сланцевский р-н, Ленинградская обл., 2016 год).
Результаты канонического анализа соответствий приведены на рисунке 2. Первая ось объясняет 65 % изменчивости, вторая ось — 23 %. Участки с максимальной каменистостью колонизировались в основном разреженными растительными группировками с преобладанием Ce-ratodon purpureus и Bryum pseudotriquetrum. Во многих исследованиях подтверждено, что именно мхи и лишайники первыми колонизируют заброшенные карьеры (17, 44, 45). Предположительно это вызвано тем, что они лучше адаптированы к неблагоприятным условиям среды, а также способны подготавливать субстрат для поселения других рас- тений (46-48). Участки с большой долей глинистой фракции колонизировали ельники травяные. Дренированные супесчаные участки были заняты различными типами сосняков. На площадках со значительным количеством скелетного материала и относительно низким содержанием мелкозема пре-обладалимелколиственные леса.
Всего в карьере обнаружили 136 видов высших растений, относящихся к 106 родам, 49 семействам, 45 порядкам, 5 классам и 4 отделам. Преобладающими по числу видов оказались семейства Fabaceae и Poaceae , включавшие по 13 видов (почти по 10 % выявленного разнообразия), а также Asteraceae и Rosaceae — по 11 видов (по 8 %). Большое число представителей семейства Fabaceae характерно для нарушенных местообитаний, Asteraceae и Rosaceae — лидирующие семейства флоры Ленинградской области. Насчитывалось 24 одновидовых семейств. Стоит отметить довольно высокую видовую насыщенность семейства Orchidaceae — 9 видов (6 % от всего списка видов), что связано с карбонатным субстратом карьера. Также было зафиксировано 22 вида мхов из 10 семейств и 11 эпилит-ных лишайников из 7 семейств, 75 видов водорослей и цианобактерий из 38 родов и 29 семейств. В карьере найдены представители всех жизненных форм растений, распространенных в Ленинградской области, согласно Н.Н. Цвелеву (49). По требовательности к богатству почв были представлены все экологические группы: от олиготрофов до эвтрофов и типичных нитрофилов, а также широко распространены виды-кальцефилы. По отношению к водному режиму почв выявлены все группы — от ксерофитов до гигрофитов. Число видов высших сосудистых растений на участках варьировало от 14 до 39 в зависимости от экотопа. В разных экотопах обнаружили 14 охраняемых видов. Максимальное сходство видового состава наблюдалось на скальных днищах карьера (коэффициент Съеренсена-Чекановского — 92 %). Здесь создавались наиболее суровые условия для развития растительности, вследствие чего сообщества были представлены ограниченным набором видов. Наименьшее число общих видов имели участки, занимающие разные позиции в рельефе.
Согласно всем рассчитанным индексам, наименьшее разнообразие соответствовало участку, где на плоской поверхности (на мягкой вскрыше) в 1970 году были высажены саженцы сосны. Такое биоразнообразие типично для сосновых лесов Ленинградской области. Наибольшее разнообразия, согласно индексу Шеннона, соответствовало недавно рекультивированному участку, где кроны деревьев еще не сомкнулись, а опушечнолуговые виды еще не вытеснены типичными лесными видами. Согласно индексу Симпсона, наибольшим разнообразием выделялся отвал карьера, где более 30 лет шла спонтанная сукцессия без вмешательства человека. На плоских участках под биологической рекультивацией биоразнообразие сокращалось. На площадках, где развитие протекало по типу спонтанной сукцессии, а именно на отвалах, биоразнообразие со временем увеличивалось. Интересно, что на сильно каменистых участках (отвалы из крупных обломков и скальные днища) развитие происходило так медленно, что за 46 лет существенных изменений не случилось. По-видимому, этого времени слишком мало для естественного преобразования субстрата водорослями, лишайниками и мхами. Наши результаты подтверждаются многочисленными данными литературы, утверждающими, что с точки зрения разнообразия растительных сообществ спонтанная сукцессия — оптимальный подход при частично управляемой рекультивации земель (50-53).
Микробиом служит связующим звеном в системе почва—растение, обеспечивая экологическую адаптацию растения к стрессовым условиям 563
нарушенных почв. Кроме того, микроорганизмы способны утилизировать многие химические соединения, участвуя в биоремедиации нарушенных территорий.
Содержание микробной биомассы в исследуемых образцах варьировало от 0,98 до 4,6 мкг С/г почвы, при этом значения увеличивались со сроком зарастания и наблюдалась тенденция к уменьшению этого показателя вниз по почвенному профилю. Согласно данным литературы (55, 56), количество микробной биомассы можно рассматривать как один из индикаторов накопления органического вещества и его минерализации. Поскольку базальное дыхание и содержание микробной биомассы сильно зависят от таких параметров, как влажность и температура (57), был рассчитан микробный метаболический коэффициент, который относится к интегральным показателям биологического состояния почв. Его значения варьировали от 0,004 до 0,022 мкг СO2-С/мг Смик•ч - 1. Полученные результаты свидетельствуют о пониженной устойчивости микробных сообществ и неэффективном использовании органического субстрата, особенно на первых этапах зарастания карьера. По данным J. Frouz с соавт. (58), активно изучавших микробиологическое состояние карьеров в Чехии, показатель почвенного дыхания на единицу микробной биомассы снижается с увеличением срока зарастания. Однако по нашим данным сложно выявить тенденцию изменения микробиологических показателей с возрастом. Также авторы отмечают, что под 30-40-летними сообществами большинство микробиологических показателей были такими же, как в ненарушенных сообществах.
В нашей работе таксономический анализ микробиомов показал, что во всех образцах доминирующим филотипом был Proteobacteria (55,7 %), за ним следовали Actinobacteria (17,0 %), Bacteroidetes (10,3 %), Acidobacteria (6,4 %) и Chloroflexi (3,8 %). Полученные результаты согласуются с данными литературы: перечисленные филы становятся доминирующими в почвах нарушенных местообитаний (25, 59, 60).
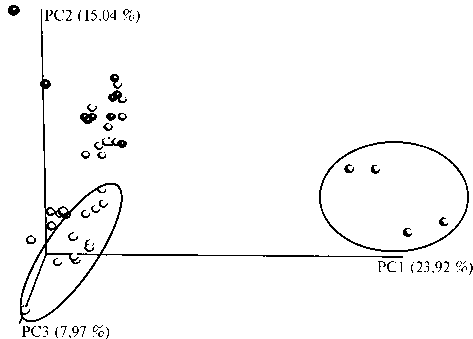
Рис. 3. Результаты анализа главных компонент (PCoA): белые кружки — старовозрастные отвалы (35 лет), серые кружки — средневозрастные отвалы (28-30 лет), черные кружки — молодые отвалы (8-16 лет). Выделенные области обозначают группы отвалов определенного возраста, объединяющиеся в клаcтеры (анализ выполнен в программе QIIME; карьер Печурки, Слан-цевский р-н Ленинградской обл., 2016 год).
ного покрова. Доминирующими таксонами
Как показал анализ главных компонент (PСoA), самым сильным фактором был возраст отвалов (19,9 % объясненной вариабельности). Микробиомы старых (35 лет и более) отвалов отчетливо обособлялись, сообщества микроорганизмов молодых (8-16 лет) и средневозрастных (28-30 лет) отвалов проявляли тенденцию к группировке в отдельные кластеры (рис. 3). Микробиомы образцов днища карьера также формировали обособленную группу, что стало следствием особых экологических условий из-за каменистости субстрата и разреженного раститель-в молодых отвалах были
Acinetobacter (8,8 % всего сообщества), Micrococcaceae (8 %) и Pseudomonas
(6 %), при этом количество этих бактерий статистически значимо уменьшалось (p < 0,05) в ряду молодые > средние > старые отвалы. Виды Acinetobacter обладают сравнительно широкой экологической нишей и могут быть найдены в местах обитания, загрязненных углеводородами, активированном иле, сточных водах, а также в отвалах вскрышных пород (61). Высокая доля псевдомонад была зафиксирована при анализе 6-летних техноземов, сформированных на отвалах в результате угледобычи (62). В средневозрастных отвалах преобладали представители Micrococcaceae (4,5 %) и Sphingomonadaceae (1,4 %). По мере увеличения возраста отвалов в структуре микробиомов повышалась доля α-протеобактерий порядка Rhi-zobiale s, главным образом за счет семейств Bradyrhizobiaceae и Hyphomicro-biaceae . Статистически значимые максимумы доли этих групп отмечали в 35-летних отвалах — соответственно 5,0 и 2,5 % сиквенсов. Известно, что бактерии Hyphomicrobiaсeae и Bradyrhizobiaceae играют важную роль в процессах превращений углерода и азота в почве (63), многие из них часто встречаются в ризосфере травянистых растений и входят в группу plantgrowth-promoting (PGPR) бактерий. По сравнению с молодыми и средневозрастными отвалами старые характеризовались значительным (в 6-8 раз) увеличением доли представителей Micromonosporaceae и Sinobacteraceae , также отмечалось существенное повышение доли бактерий Chitinophagaceae и Cytophagaceae в структуре микробиомов отвалов 35-летнего возраста.
Сообщества влажных террас тоже несколько отличались от микробиомов сухих террас и оснований отвалов, однако достоверных различий выявить не удалось. Кислотность (pH), которая служит одним из главных почвенных факторов и основным предиктором состава микробного сообщества (64-66), также не оказывала достоверного влияния на состав исследуемых микробиомов. По-видимому, формирование определенной структуры микробных сообществ становилось результатом совокупного действия факторов с их наиболее благоприятным сочетанием в 35-летних отвалах.
Результаты таксономического анализа состава микробиомов отвалов разного возраста выявили отчетливую смену видового состава микроорганизмов по мере увеличения возраста отвалов. В ранневозрастных отвалах преобладали микроорганизмы, которые можно отнести к экологической группе копиотрофов и наличие которых в почве маркирует ранние этапы сукцессии. Согласно данным Y. Li с соавт. (25), наиболее важная стадия восстановления микробиома проходит между 15-м и 20-м годами после начала рекультивации. В наших исследованиях, начиная со средневозрастных отвалов (старше 16 лет), в сообществе становились преобладающими олиготрофные группы бактерий, в том числе бактерии, участвующие в разложении сложных биополимеров. Доминирование этой группы микроорганизмов в составе сообщества может характеризовать переход микробоценоза в климаксную стадию.
Таким образом, неблагоприятный гранулометрический состав становится лимитирующим фактором развития растительности вследствие высокой каменистости на скальных днищах. Наличие в карьере участков с неодинаковыми физическими параметрами субстрата, находящихся на разных стадиях зарастания, способствовало развитию высокого разнообразия растительных сообществ. Была обнаружена смена видового состава микроорганизмов в процессе сукцессии, при этом на ранних стадиях преобладали группы копиотрофов, на более поздних — олиготрофы. Установлено, что для разработки оптимального плана рекультивации территорий и ее дальнейшего сельскохозяйственного использования необходим мониторинг каждого участка карьера, поскольку эдафические условия мо-565
гут различаться в зависимости от положения в рельефе и способа воздействия на земли. Восстановление территорий в качестве земель сельскохозяйственного назначения требует дорогостоящих мероприятий по выпо-лаживанию и нивелированию высокой гетерогенности субстрата, что экономически затратно. С точки зрения вопросов биоразнообразия лучшим методом для рекультивации карбонатных карьеров может считаться создание благоприятных физических условий субстрата и оставление участка под самозарастание.
Список литературы Динамика растительного сообщества и микробиома хроносерий посттехногенных почв в известняковом карьере в условиях рекультивации
- Williamson N.A., Johnson M.S., Bradshaw A.D. Mine wastes reclamation. Mining Journal Books, Ltd., London, United Kingdom, 1982.
- Abakumov E., Trubetskoj O., Demin D., Celi L., Cerli C., Trubetskaya O. Humic acid characteristics in podzol soil chronosequence. Chem. Ecol., 2010, 26: 59-66 ( ) DOI: 10.1080/02757540.2010.497758
- Abakumov E.V., Maksimova E.I., Lagoda A.V., Koptseva E.M. Soil formation in the quarries for limestone and clay production in the Ukhta region. Eurasian Soil Science, 2011, 44(4): 380-385 ( ) DOI: 10.1134/S1064229311040028
- Исаченко Г.А., Резников И.А. Ландшафты Санкт-Петербурга: эволюция, динамика, разнообразие. Биосфера, 2014, 2: 196-200.
- Abakumov E.V., Cajthaml T., Brus J., Frouz J. Humus accumulation, humification, and humic acid composition in soils of two post-mining chronosequences after coal mining. J. Soils Sediments, 2013, 13(3): 491-500 ( ) DOI: 10.1007/s11368-012-0579-9
- Johnson N., Payton P., Spalding A. The conservation value of metalliferous mine sites in Cornwall. Cornwall Archaeological Unit and Institute of Cornish Studies, Truro, 1996.
- Key R. Bare ground and the conservation of invertebrates. British Wildlife, 2000, 11: 183-191.
- Андроханов В.А., Овсянникова С.В., Курачев В.М. Техноземы: свойства, режимы, функционирование. Новосибирск, 2000.
- Арчегова И.Б. Закономерности формирования биоразнообразия растительных сообществ в восстанавливающихся и преобразующихся экосистемах в разных типах техногенных объектов на Северо-Востоке Европейской части России. Вестник института биологии Коми научного центра Уральского отделения РАН, 2013, 3(179): 24-27.
- Kuráž V., Frouz J., Kuráž M., Mako A., Shustr V., Cejpek J., Romanov O.V., Abakumov E.V. Changes in some physical properties of soils in the chronosequence of self-overgrown dumps of the Sokolov quarry-dump complex, Czechia. Eurasian Soil Science, 2012, 45(3): 266-272 ( ) DOI: 10.1134/S1064229312030076
- Андронов Е.Е., Петрова С.Н., Пинаев А.Г., Першина Е.В., Рахимгалиева С.Ж., Ахмеденов К.М., Горобец А.В., Сергалиев Н.Х. Изучение структуры микробного сообщества почв разной степени засоления с использованием T-RFLP и ПЦР с детекцией в реальном времени. Почвоведение, 2012, 2: 173.
- Копцева Е.М. Естественное восстановление растительности на техногенных местообитаниях Крайнего Севера: Ямальский сектор Арктики. СПб, 2005.
- Нешатаев В.Ю., Карапухин Н.С., Ефремов Д.Ф., Кноль В.В., Нешатаев М.В., Штак К.Д. Практическое пособие по восстановлению растительного покрова на землях, нарушенных открытыми горными разработками при освоении месторождений полезных ископаемых в условиях Камчатского края. СПб, 2012.
- Сумина О.И. Формирование растительности на техногенных местообитаниях Крайнего Севера России. Автореф. докт. дис. СПб, 2011.
- Абакумов Е.В., Гагарина Э.И. Почвообразование в посттехногенных экосистемах карьеров на Северо-Западе Русской равнины. СПб, 2006.
- Yarwood S., Wick A., Williams M., Daniels W.L. Parent material and vegetation influence soil microbial community structure following 30-years of rock weathering and pedogenesis. Microb. Ecol., 2015, 69(2): 383-394 ( ) DOI: 10.1007/s00248-014-0523-1
- Labbé L., Lanctôt B., Fortin, J.A. Étalement des îlots d'aulnes et enrichis sement dessolsau complexe La Grande. Rapport pour la Vice-présidence Environnement, Hydro-Québec. Institut de recherche en biologie végétale de Montréal, Montréal, Qué., 1995.
- Laitinen J., Rehell S., Oksanen J. Community and species responses to water level fluctuations with reference to soil layers in different habitats of mid-boreal mire complexes. Plant Ecol., 2008, 194(1): 17-36 ( ) DOI: 10.1007/s11258-007-9271-3
- Gretarsdottir J., Aradottir A.L., Vandvik V., Heegaard E., Birks H.J.B. Long-term effects of reclamation treatments on plant succession in Iceland. Restor. Ecol., 2004, 12(2): 268-278 ( ) DOI: 10.1111/j.1061-2971.2004.00371.x
- Tormo J., Bochet E., Garcia-Fayos P. Roadfill revegetation in semiarid Mediterranean environments. Part II: Topsoiling, species selection, and hydroseeding. Restor. Ecol., 2007, 15(1): 97-102 ( ) DOI: 10.1111/j.1526-100X.2006.00194.x
- Стифеев А.И., Головастикова А.В., Бессонова Е.А. Изменение состава и структуры микробного сообщества в условиях техногенного ландшафта отвалов михайловского ГОКа КМА. Вестник Курской государственной сельскохозяйственной академии, 2011, 4(4): 40-41.
- Dangi S.R., Stahl P.D., Wick A.F., Ingram L.J., Buyer J.S. Soil microbial community recovery in reclaimed soils on a surface coal mine site. Soil Sci. Soc. Am. J., 2012, 76(3): 915-924 ( ) DOI: 10.2136/sssaj2011.0288
- Acosta-Martinez V., Dowd S.E., Sun Y., Wester D., Allen V. Pyrosequencing analysis for characterization of soil bacterial populations as affected by an integrated livestock-cotton production system. Appl. Soil Ecol., 2010, 45(1): 13-25 ( ) DOI: 10.1016/j.apsoil.2010.01.005
- Liu S., Liu W., Yang M., Zhou L., Liang H. The genetic diversity of soil bacteria affected by phytoremediation in a typical barren rare earth mined site of South China. SpringerPlus, 2016, 5(1): 1131 ( ) DOI: 10.1186/s40064-016-2814-0
- Li Y., Wen H., Chen L., Yin T. Succession of bacterial community structure and diversity in soil along a chronosequence of reclamation and re-vegetation on coal mine spoils in China. PLoS ONE, 2014, 9(12): e115024 ( ) DOI: 10.1371/journal.pone.0115024
- Luna L., Pastorelli R., Bastida F. The combination of quarry restoration strategies in semiarid climate induces different responses in biochemical and microbiological soil properties. Appl. Soil Ecol., 2016, 107: 33-47 ( ) DOI: 10.1016/j.apsoil.2016.05.006
- Полянская Л.М. Микробная сукцессия в почве. Автореф. докт. дис. М., 1998.
- Fierer N., Leff J.W., Adams B.J., Nielsen U.N., Bates S.T., Lauber C.L., Owens S., Gilberte J.A., Wall D.H., Caporaso J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. PNAS USA, 2012, 109(52): 21390-21395 ( ) DOI: 10.1073/pnas.1215210110
- Zhang C., Xue S., Liu G.B., Zhang C.S. Effects of slope aspect on soil chemical and microbial properties during natural recovery on abandoned cropland in the Loess plateau, China. Progress in Environmental Science and Engineering, 2011, 356-360: 2422-2429 (doi: 10.4028/www.scientific.net/AMR.356-360.2422).
- Шапошников А.И., Белимов А.А., Кравченко Л.В., Виванко Д.М. Взаимодействие ризосферных бактерий с растениями: механизмы образования и факторы эффективности ассоциативных симбиозов. Сельскохозяйственная биология, 2011, 3: 16-22.
- Burns R.G., DeForest J.L, Marxsen J., Sinsabaugh R.L., Stromberger M.E., Wallenstein M.D., Weintraub M.N., Zoppini A. Soil enzymes in a changing environment: current knowledge and future directions. Soil Biol. Biochem., 2013, 58: 216-234 ( ) DOI: 10.1016/j.soilbio.2012.11.009
- Ananyeva N.D., Susyan E.A., Chernova O.V., Wirth S. Microbial respiration activities of soils from different climatic regions of European Russia. Eur. J. Soil Biol., 2008, 2: 147-157.
- Цитович И.К. Курс аналитической химии. М., 1994.
- Растворова О.Г. Физика почв (практическое руководство). Л., 1983.
- Пономарева В.В., Плотникова Т.А. Миграционная и седиментационная способность черных и бурых гуминовых кислот и их соединений с кальцием. В кн.: Проблемы почвоведения/Под ред. М.А. Глазовской, И.А. Соколова. М., 1978: 65-72.
- Anderson J.P.E., Domsch K.H.A. Physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem., 1978, 10(3): 215-221 ( ) DOI: 10.1016/0038-0717(78)90099-8
- Blanchet F.G., Legendre P., Borcard D. Forward selection of explanatory variables. Ecology, 2008, 89(9): 2623-2632 ( ) DOI: 10.1890/07-0986.1
- Ter Braak C.J.F. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecology, 1986, 67(5): 1167-1179 ( ) DOI: 10.2307/1938672
- Edgar R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics, 2010, 26(19): 2460-2461 ( ) DOI: 10.1093/bioinformatics/btq461
- Huber J.A., Butterfield D.A., Baross J.A. Temporal changes in archaeal diversity and chemistry in a mid-ocean ridge subsea floor habitat. Appl. Environ. Microb., 2002, 68: 1585 ( ) DOI: 10.1128/AEM.68.4.1585-1594.2002
- Hugron S., Andersen R., Poulin M., Rochefort L. Natural plant colonization of borrow pits in boreal forest highlands of eastern Canada. Botany, 2011, 89: 451-465 ( ) DOI: 10.1139/B11-036
- Gilardelli F., Sgorbati S., Armiraglio S., Citterio S., Gentili R. Assigning plant communities to a successional phase: time trends in abandoned limestone quarries. Plant Biosyst., 2016, 150(4): 799-808 ( ) DOI: 10.1080/11263504.2015.1011722
- Řehounková K., Prach K. Spontaneous vegetation succession in gravel-sand pits: a potential for restoration. Restor. Ecol., 2008, 2(16): 305-312 ( ) DOI: 10.1111/j.1526-100X.2007.00316.x
- Karim M.N., Mallik A.U. Roadside revegetation by native plants: I. Roadside microhabitats, floristic zonation and species traits. Ecol. Eng., 2008, 32(3): 222-237 ( ) DOI: 10.1016/j.ecoleng.2007.11.003
- Kershaw G.P., Kershaw L.J. Successful plant colonizers on disturbances in tundra areas of northwestern Canada. Arctic Alpine Res., 1987, 19(4): 451-460 ( ) DOI: 10.2307/1551411
- Alpert P. The discovery, scope, and puzzle of desiccation tolerance in plants. Plant Ecol., 2000, 151(1): 5-17 ( ) DOI: 10.1023/A:1026513800380
- Beckett R.P., Kranner I., Minibayeva F.V. Stress physiology and the symbiosis. In: Lichen biology. 2nd ed./T.H. Nash, III (ed.). Cambridge University Press, Cambridge, UK, 2008.
- Sancho L.G., de la Torre R., Horneck G., Ascaso C., de los Rios A., Pintado A., Wierzchos J., Schuster M. Lichens survive in space: results from the 2005 LICHENS experiment. Astrobiology, 2007, 7(3): 443-454 ( ) DOI: 10.1089/ast.2006.0046
- Цвелев Н.Н. Определитель сосудистых растений Северо-Западной России (Ленинградская, Псковская и Новгородская области). СПб, 2000.
- Hodačová D, Prach K. Spoil heaps from brown coal mining: technical reclamation vs spontaneous revegetation. Restor. Ecol., 2003, 11: 385-391 ( ) DOI: 10.1046/j.1526-100X.2003.00202.x
- Mudrák O., Frouz J., Velichová V. Underestory vegetation in reclaimed and unreclaimed post-mining forest stands. Ecol. Eng., 2010, 36: 783-790 ( ) DOI: 10.1016/j.ecoleng.2010.02.003
- Šebelíková L., Řehounková K., Prach K. Spontaneous regeneration vs. forestry reclamation in post-mining sand pits. Environ. Sci. Pollut. Res., 2016, 23: 13598-13605.
- Tropek R., Černá I., Straka J., Čížek O., Konvička M. Is coal combustion the last chance for vanishing insects of inland drift sand dunes in Europe? Biol. Conserv., 2013, 162: 60-64 ( ) DOI: 10.1016/j.biocon.2013.03.027
- Арчегова И.Б. О гумусе в связи с нетрадиционным пониманием почвы. Почвоведение, 1992, 1: 58-64.
- Ананьева Н.Д. Микробиологические аспекты самоочищения и устойчивости почв. М., 2003.
- Jenkinson D.S., Ladd J.N. Microbial biomass in soil: measurement and turnover. Soil Biochemistry, 1981, 5: 415-471.
- Приходько В.Е., Сиземская М.Л. Базальное дыхание и состав микробной биомассы целинных, агро-и лесомелиорированных полупустынных почв Северного Прикаспия. Почвоведение, 2015, 8: 974.
- Frouz J., Nováková A. Development of soil microbial properties in topsoil layer during spontaneous succession in heaps after brown coal mining in relation to humus microstructure development. Geoderma, 2005, 129: 54-64 ( ) DOI: 10.1016/j.geoderma.2004.12.033
- Mummey D.L., Stahl P.D., Buyer J.S. Soil microbiological properties 20 years after surface mine reclamation: spatial analysis or reclaimed and undisturbed sites. Soil Biol. Biochem., 2002, 34: 1717-1725 ( ) DOI: 10.1016/S0038-0717(02)00158-X
- Megharaj M., Naidu R., Ramadass K., Samkumar R.A., Satheesh V., Subashchandrabose S.R., Thavamani P., Venkateswarlu K. Microbes from mined sites: Harnessing their potential for reclamation of derelict mine sites. Environ. Pollut., 2017, 230: 495-505 ( ) DOI: 10.1016/j.envpol.2017.06.056
- Doughari H.J., Ndakidemi P.A., Human I.S., Benade S. The ecology, biology and pathogenesis of Acinetobacter spp.: an overview. Microbes Environ., 2011, 26: 101-112 ( ) DOI: 10.1264/jsme2.ME10179
- Urbanova M., Kopecky J., Valaskova V., Mareckova M.S., Elhottova D., Kyselkova M., Loccoz Y.M., Baldrian P. Development of bacterial community during spontaneous succession on spoil heaps after brown coal mining. FEMS Microbiol. Ecol., 2011, 78: 59-69 ( ) DOI: 10.1111/j.1574-6941.2011.01164.x
- Андронов Е.Е., Иванова Е.А., Першина Е.В., Орлова О.В., Круглов Ю.В., Белимов А.А., Тихонович И.А. Анализ показателей почвенного микробиома в процессах, связанных с почвообразованием, трансформацией органического вещества и тонкой регуляции вегетационных процессов. Бюллетень Почвенного института им. В.В. Докучаева, 2015, 80: 83-94.
- Lauber C.L., Hamady M., Knight R., Fierer N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microb., 2009, 75: 5111-5120 ( ) DOI: 10.1128/AEM.00335-09
- Kaiser K., Wemheuer B., Korolkow V., Wemheuer F., Nacke H., Schöning I. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Scientific Reports, 2016, 6: 33696 ( ) DOI: 10.1038/srep33696
- Rousk J., Bååth E., Brookes P.C., Lauber C.L., Lozupone C., Caporaso J.G. Soil bacterial and fungal communities across a pH gradient in an arable soil. The ISME Journal, 2010, 4: 1340-1351 ( ) DOI: 10.1038/ismej.2010.58
