Динамика растительности верховых болот

Автор: Кузнецов Олег Леонидович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
В статье рассмотрены основные закономерности динамики растительного покрова верховых болот Европейской части России в голоцене и их особенности в различных геоморфологических условиях.
Верховые болота, растительность, динамика, геоморфология, голоцен
Короткий адрес: https://sciup.org/148200807
IDR: 148200807 | УДК: 581.524.32:551.438.222(470.22)
Dynamics of raised bogs vegetation
This paper observes the main dynamical trends for raised bogs plant cover in European part of Russia during Holocene under varying geomorphological conditions.
Текст научной статьи Динамика растительности верховых болот
Болотные экосистемы это важный и специфический компонент большинства ландшафтов таежной зоны. Обладая уникальным свойством торфонакоп-ления, они являются аккумуляторами углерода, а также «летописями» природных процессов, происходивших на протяжении тысячелетий. Анализ озерно-болотных отложений с использованием комплекса палеоботанических и палеогеографических методов позволяет выполнить реконструкции динамики растительности и климатических условий как на обширных территориях [5, 7, 15], так и конкретных болотах, что активно применяют болотоведы в своих исследованиях [1, 3, 11]. В основе реконструкции сукцессий растительности болот лежит ботанический состав торфов, по преобладающим остаткам растений в которых достаточно точно можно установить материнское торфоотлагающее сообщество и режим его увлажнения с учетом данных по степени сохранности разных видов растений в торфах [1, 4]. Необходимо также учитывать внедрение корней сосудистых растений в ранее отложенные слои торфа.
Развитие болот сопровождается накоплением торфяных отложений. Это, с од ной стороны перераспределяет все прямодействующие факторы среды, а с другой – обусловливает обеднение субстрата питательными веществами, доступными для растительности. Прогрессирующее обеднение торфяных отложений приводит к закономерным сменам растительного покрова, которые идут по типу эн-доэкогенетических (автогенных) сукцессий. В процессе развития болотных массивов во вмещающих их ландшафтах могут происходить и достаточно сильные изменения природных условий (климат, понижения базиса эрозии, уровней водоемов), что приводит к резким сменам в растительном покрове и функционировании болот, такие сукцессии уже являются аллогенными. В динамике растительного покрова отдельных болотных массивов в течение голоцена имеют место оба типа сукцессий, преобладание того или иного из них обусловливается условиями залегания массива.
В пределах лесной зоны Европейской части России представлен широкий спектр типов болотных массивов, выделяемых по растительному покрову [16],
среди которых наибольшие площади занимают болота двух классов: сфагновые (13 типов, из них 11 верховых, включающих 9 типов с грядовомочажинными и грядово-озерковыми комплексами, 2 - сосново-кустарничково-сфагновые) и травяно-сфагново-гипновые (аапа – 3 типа). Болотные массивы этих классов являются резко контрастными как по современной структуре растительного покрова, так и своей динамике в голоцене. Динамика болот аапа типа подробно рассмотрена в ряде работ [8, 9]. В данной статье анализируются особенности развития верховых грядово-мочажинных болот Европейской части России.
Верховые сфагновые грядово-мочажинные болотные массивы широко распространены во всех подзонах тайги, значительно реже встречаются и южнее. В пределах двух физико-географических стран (Фенноскандия и Русская равнина), а также их природных подзон, верховые болота различаются как по растительности, так и условиям залегания в рельефе, что нашло отражение в их динамике.
На Русской равнине крупные верховые болота и их системы (с площадями от нескольких тысяч до десятков тысяч га) приурочены к низменностям различного генезиса, часто залегают также на плоских водоразделах и надпойменных террасах [14, 16]. Образование большинства из них начиналось путем зарастания остаточных мелководных водоемов травяными и травяно-гипновыми евтрофными сообществами. На многих болотах евтрофная фаза развития была непродолжительной. После заполнения первичных депрессий и достижением болотом плоской формы поверхности наступала кратковременная мезотрофная фаза, которая представлена в торфяных залежах тонкими слоями переходных торфов. Затем болото начинало приобретать слабовыпуклую форму, происходил его переход на атмосферное питание и распространение омбро-трофных сфагновых сообществ, которые и отложили основные толщи торфяных залежей этих болот. Омбротрофная (верховая) фаза развития на многих болотах в подзонах средней и южной тайги началась в конце бореального периода, уже в атлантический период в центральных частях массивов стали формироваться грядово-мочажинные комплексы [6], часть мочажин которых затем превратилась во вторичные озерки.
Большинство грядово-мочажинных верховых болот Русской равнины являются концентрическими, в южных районах превышение их центральных частей над окрайками достигает 6-8 метров, в подзоне северной тайги они менее выпуклые – 2-3 метра. Стратиграфия их торфяных залежей довольно однообразна. На многих имеются слои пушицевого и древесно-пушицевого торфов с высокой степенью разложения и различной мощности, которые ранее трактовались как «пограничный горизонт», отложившийся в более сухой суббореальный период. Однако они имеют различный возраст и часто, наряду с общеклиматическими колебаниями, их отложение обусловлено изменением локальных гидрологических условий на болоте. Установлено, что в грядово-мочажинных комплексах, существующих на протяжении нескольких тысячелетий, происходит пульсация границ гряд и мочажин и даже смена их местами [13]. В субатлантический период, начавшийся около 2500 лет назад, характеризующийся более влажным и прохладным климатом по сравнению с суббореалом, произошло увеличение площади грядово-мочажинных комплексов на верховых болотах, а в их структуре возросла доля мочажин и озерков. В этот период также происходила активная экспансия болот на прилегающие леса в равнинных ландшафтах и урочищах.
В восточной Фенноскандии (Карелия и Мурманская область) большинство верховых грядовомочажинных болот имеют эксцентрическую слабо выпуклую поверхность. Это обусловлено их развитием на склонах и у подножий склонов холмов и гряд, а также на наклонных террасах Беломорского побережья. Болотные массивы в основном небольшие (от десятков до нескольких сотен гектаров), только на террасах Прибеломорской низменности дистрофные вересково-лишайниковые грядово-озерковые массивы и их системы достигают десятков тысяч гектаров.
Установлено, переход болотных массивов и их отдельных участков в верховую фазу развития в Карелии имеет большой временной интервал – от 1 до 9,3 тыс. л. н. Грядово-мочажинные комплексы, имеющие возраст не менее 5-6 тыс. л., в том числе и с первичными мочажинами, выявлены в разных районах Карелии [10, 11]. Мощность верховых торфов на грядово-мочажинных болотах вне При-беломорской низменности варьирует от 1,0 до 4,0 м, а на обширных прибеломорских грядовоозерковых болотах достигает 6,5 м. На маленьких сфагновых болотах (1-3 га) в бессточных межкамо-вых котловинах встречаются залежи, включающие до 6,0 м верхового торфа. В отличие от Русской равнины, в Фенноскандии много приозерных верховых болот, на которых сохранились остаточные первичные озера, оказывающие влияние на их динамику. Небольшие размеры болот, а также достаточно расчлененный рельеф обусловливают высокую лабильность их гидрологического режима при изменениях климата и базиса эрозии на водосбо- рах, что четко отражается на динамике болот. Отсюда выявлен широкий спектр сукцессионных рядов в развитии верховых болот в Карелии. Переход из мезотрофной фазы развития болот в омбротроф-ную происходил разными путями, как с резким изменением условий увлажнения и минерального питания и со сменой топяных переходных торфов (осоковый, шейхцериево-сфагновый) верховыми кочковыми (фускум, магелланикум), так и постепенно, практически без изменения условий увлажнения. При этом шло выпадение из растительного покрова минеротрофных видов (Carex rostrata, C. lasiocarpa, Menyanthes trifoliata) и их замена на Scheuchzeria palustris, Carex limosa [11].
Важным показателем динамики болотных экосистем является процесс торфонакопления, оцениваемый в первую очередь по вертикальному приросту торфа. На верховых болотах Карелии средний прирост торфа в разные периоды голоцена колеблется от 0,18 до 0, 85 мм/год, а в среднем составляет – 0,52 мм/год, при этом прирост отдельных слоев на некоторых болотах достигает 1,5-2,0 мм/год [17]. Это свидетельствует, что мощность залежей не является надежным индикатором их возраста. Эксцентрические массивы имеют обычно залежи меньшей глубины (до 3-4 м) и более низкие показатели прироста торфа, чем болота в бессточных и приозерных котловинах, а также на террасах Белого моря. Следует отметить, что даже идентичные по составу слои торфа в конкретном разрезе иногда резко различаются по скорости прироста, поэтому только сочетание ботанического, споровопыльцевого и радиоуглеродного анализов позволяют выполнять достаточно точные и детальные реконструкции растительности болот. Это хорошо иллюстрирует разрез с болота Вирменское (рис.), расположенного на второй террасе Белого моря. В скважине, пробуренной в центре болота, выполнены 11 радиоуглеродных датировок [11]. Датировки в сочетании с данными по стратиграфии залежи позволили детально реконструировать динамику центральной части болота. Его формирование на этом участке началось с омбротрофной фазы около 8700 л.н. (калиброванный возраст) с сосново-пушицево-сфагновых сообществ (I), через 700 лет сменившихся пушицево-сфагновыми (II) и просуществовавших здесь на протяжении 1600 лет. Тор-фонакопление в течение этих стадий было невысоким – 0,23 -0,56 мм/год. Затем на их месте возникла мочажина, сначала шейхцериево-сфагновая (III), сменившаяся затем пушицево-сфагновой (IV), которая сохранялась здесь на протяжении более трех тысяч лет. В период существования мочажины прирост торфа вначале был очень низким – 0,15 мм/год, затем резко возрос до 0,79 мм/год. Около 3 тыс. л.н. мочажинное сообщество сменилось грядовым со Sphagnum fuscum (V), прирост торфа за время его существования сильно варьировал – 0,28 – 1,0 мм/год. В последние 200-300 лет произошла сукцессия сфагнового сообщества в вересково-во-
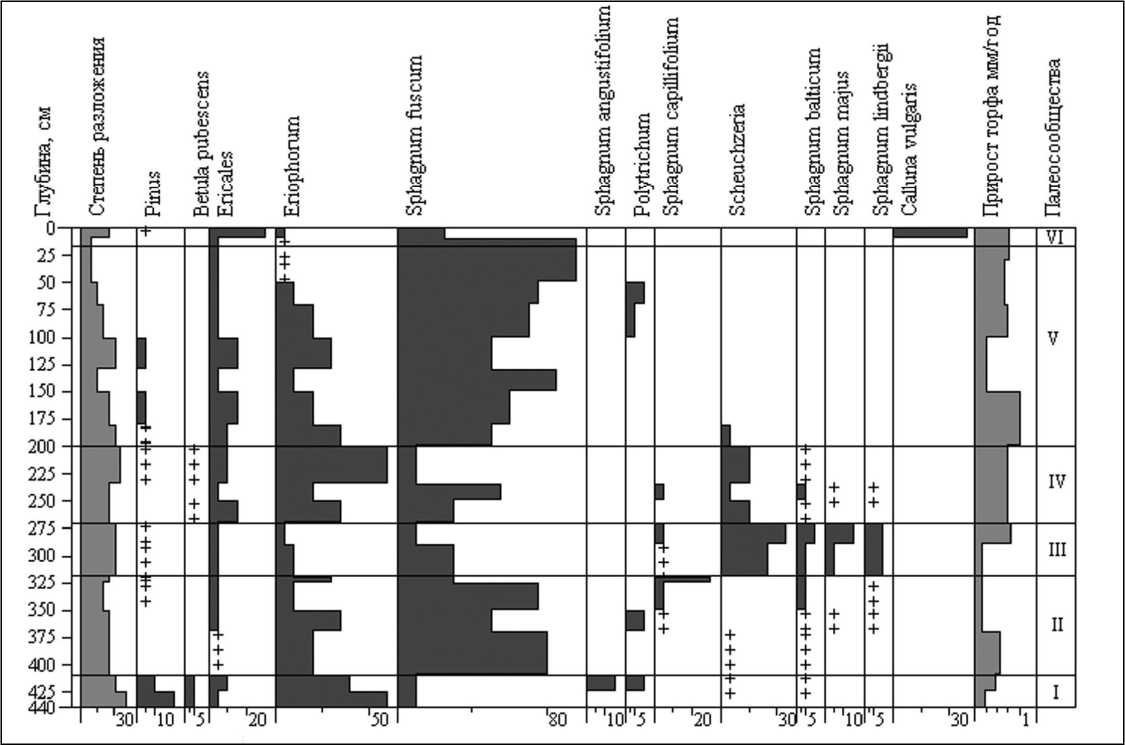
Рис. Сукцессии растительности и вертикальный прирост торфа на болоте Вирменское
Палеосообщества: I – Pinus sylvestris–Eriophorum vaginatum–Sphagnum fuscum+S. angustifolium, II – Eriophorum vaginatum–Sphagnum fuscum , III – Scheuchzeria palustris+ Eriophorum vaginatum–Sphagnum sect. Cuspidata, IV – Ericales– Eriophorum vaginatum+ Scheuchzeria palustris– Sphagnum fuscum , V – Ericales–Eriophorum vaginatum– S. fuscum, VI – Calluna vulgaris+Empetrum nigrum– S. fuscum+Cladina ронично-лишайниковое (VI), отлагающее сейчас кустарничковый торф, который еще находится в процессе торфогенеза и уплотнения. Прирост торфа в этом разрезе в течение голоцена сильно варьировал – 0,15-1,00 мм/год, причем даже слои со сходным ботаническим составом имеют значительные различия в приросте. Это обусловлено конкретными условиями увлажнения и стока с болотного массива, так как формирование мочажин связано с задержкой излишков воды в центре болота.
Подобные сукцессии выявлены и на других болотах такого типа в Прибеломорье [2], при этом средние для всего голоцена показатели прироста торфа довольно близкие, однако на конкретных массивах в отдельные периоды голоцена колебания прироста значительные, в том числе и в одноименных видах торфа [12].
Только комплексные исследования стратиграфии торфяных залежей с сериями радиоуглеродных датировок позволяют выполнять достаточно точные реконструкции динамики болотных экосистем. Мощность торфяной залежи не является хорошим показателем возраста болота.
Работа выполнена в рамках ФЦП (контракт № 02.740.11.0700).
Список литературы Динамика растительности верховых болот
- Елина Г.А. Реконструкция растительности болот по ботаническому и спорово-пыльцевому анализам//Общие методы изучения истории современных экосистем. М.: Наука, 1979. С. 62-79.
- Елина Г.А. Принципы и методы реконструкции и картирования растительности голоцена. Л.: Наука, 1981. 159 с.
- Елина Г.А., Антипин В.К. Эндои экзогенные сукцессии растительности болот бассейна Онежского озера в голоцене//Ботан. журн. 1992. Т. 77. №3. С. 16-29.
- Елина Г.А., Юрковская Т.К. Методы определения палеогидрологического режима как основа объективизации причин сукцессий растительности болот//Ботан. журн. 1992. Т. 77. № 7. С. 120-124.
- Елина Г. А., Лукашов А. Д., Юрковская Т.К. Позднеледниковье и голоцен восточной Фенноскандии (палерастительность и палеогеография). Петрозаводск: Карельский научный центр РАН, 2000. 242 с.
- Иванов К.Е., Кузьмин Г.Ф. Строение торфяной залежи под грядово-мочажинными комплексами верховых болот//Вестн. ЛГУ. 1982. № 12. С. 70-81.
- Климанов В.А. К методике восстановления количественных характеристик климата прошлого//Вест. МГУ, сер.геогр., 1976. № 2. С. 92-98.
- Кузнецов О.Л. О развитии аапа болот северной Карелии//Болота Европейского Севера СССР. Петрозаводск: Карельский филиал АН СССР, 1980. С. 92-113.
- Кузнецов О.Л. Структура и динамика грядово-мочажинных комплексов аапа болот//Ботан. журн. 1982. Т. 67. № 10. С. 1394-1400.
- Кузнецов О.Л. Структура и динамика растительного покрова болотных экосистем Карелии: Автореф. дис. д-р биол. наук. Петрозаводск, 2006. 53 с.
- Кузнецов О.Л. Особенности торфонакопления и динамики некоторых типов болотных массивов Карелии//Направления исследований в современном болотоведении России. С.-Пб. -Тула: РБО, 2010. С. 96-112.
- Кузнецов О.Л., Филимонова Л.В. Стратиграфия и прирост торфяных залежей верховых болот побережья Белого моря в голоцене//Геология морей и океанов. Матер. XIX Межд. школы по морской геологии. (Москва, 14-18 ноября 2011 г.). М.: Геос, 2011. C. 192-196. 13.
- Кузьмин Г.Ф. Развитие олиготрофных болотных систем и перспективы их использования: Автореф. дис. канд. геогр. наук. Л., 1980. 21 с.
- Тюремнов С.Н. Торфяные месторождения. М.: Недра, 1976. 487 с.
- Хотинский Н.А. Голоцен северной Евразии. М.: Наука, 1977.199 c.
- Юрковская Т.К. География и картография растительности болот европейской части России и сопредельных территорий. СПб.: БИН РАН, 1992. 256 с.
- Kuznetsov O., Mäkilä M. Peat increment and carbon accumulation on karelian mires in Holocene time//Резервуары и потоки углерода в лесных и болотных экосистемах бореальной зоны. Матер. Межд. конф. Сыктывкар, 26-30 сентября 2011. Сыктывкар: КНЦ УрО РАН, 2011. С. 7-8.
