Динамика содержания карбоновых кислот в проростках огурца при гипертермии в зависимости от теплового и холодового закаливания

Автор: Чудинова Л.А., Кайгородов Р.В., Пархоменко В.В., Ощепкова А.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
Представлены результаты изучения содержания щавелевой, лимонной и фумаровой кислот в проростках огурца, подвергшихся температурному воздействию (тепловое и холодовое закаливание и гипертермия). Установлено участие этих органических кислот в механизме теплового и холодового закаливания растений к действию последующей гипертермии.
Карбоновые кислоты, холодовое и тепловое закаливание, гипертермия, перекрестная адаптация, огурец
Короткий адрес: https://sciup.org/147204700
IDR: 147204700 | УДК: 581.1:632.122.1
Dynamic of carboxylic acids in cucumber seedlings under hyperthermia depending on heat and cold hardening
The results of studying the content of oxalic, citric and fumaric acids seedlings of cucumber. Established part of the organic acids in the mechanism of heat and cold hardening plants to action subsequent hyperther-mia.
Текст научной статьи Динамика содержания карбоновых кислот в проростках огурца при гипертермии в зависимости от теплового и холодового закаливания
В настоящее время довольно хорошо изучена физиология и биохимия теплового шока, который рассматривается как неспецифический стрессор, способный модифицировать метаболизм любого живого организма. Показано, что последствия будут зависеть от режима гипертермии, генетических особенностей организма, стадии онтогенеза, а также от теплового или холодового закаливания [Таланова и др., 1993; Кузнецов, Дмитриева, 2006; Чудинова, Суворов, 2011]. Последнее рассматривается как неспецифическое воздействие, запускающее адаптивные процессы [Ба-лагурова, Акимова, Титов, 2001; Трунова, 2007; Титов, Таланова, 2009; Чудинова, Суворов, 2011]. Однако молекулярно-биохимические механизмы закаливания остаются до конца не выясненными. Недостаточно исследованы системы устойчивости растений к одновременному действию пониженной и высокой температуры. Анализ литературы демонстрирует их многообразие и вовлечение в них низкомолекулярных метаболитов, в том числе пролина, полиаминов, веществ фенольной природы, а также каротиноидов. Вклад каждого из этих соединений в защитный ответ организма неравнозначен и, возможно, приурочен к различным стадиям адаптационного процесса или является видоспецифичным [Чиркова, 2002; Титов, Таланова, 2009].
В литературе имеются указания на участие органических кислот в механизмах адаптации. Так, некоторые авторы отметили изменение содержания некоторых органических кислот у галофитов и ряда гликофитов при засолении [Матухин, 1963]. Изучение обмена органических кислот у растений в условиях стресса представляет интерес с точки зрения связующего звена между углеводным и азотным обменом. Однако оценка значимости процессов обмена органических кислот для формирования устойчивости к температурным воздействиям фактически не проводилась.
В связи с вышеизложенным, целью нашей работы было изучение влияния теплового и холодового закаливания на содержание некоторых карбоновых кислот в проростках теплолюбивых растений (огурец) в процессе их адаптации к последующей гипертермии.
Объекты и методы исследования
Проростки огурца посевного ( Cucumis sativus L.) сорта «Родничок» в течение 15 сут. выращивали в климатической камере ШН-М на вермикулите при температуре 22 ° С, влажности 80-85% и освещенности люминесцентными лампами 18 ч. в сутки (1 вариант – контроль). Затем часть проростков (опытные варианты) подвергали температурному воздействию по следующей схеме: 2-й вариант - гипертермия (40 ° С, 7 сут.); 3-й вариант - тепловое закаливание (30 ° С, 1 сут.), затем гипертермия (40 ° С, 6 сут.); 4-й вариант - холодовое закаливание (10 ° С, 1 сут.), затем гипертермия (40 ° С, 6 сут.).
Режимы тепловой и холодовой обработки были выбраны на основании данных литературы [Чуди-
нова, Суворов, Механошина, 2001; Титов, Таланова, 2009; Чудинова, Суворов, 2011].
Для анализа брали растения через 12 ч. (стадия стресс-реакции адаптации) и 72 ч (стадия адаптации) после окончания всех обработок.
Анализировали прирост проростков за 3 сут. В побегах, фиксированных паром (5 мин.) и досушенных до воздушно-сухого состояния, определяли содержание щавелевой, лимонной, фумаровой кислот методом высокоэффективной жидкостной хроматографии (ВЭЖХ) на приборе Ultimate 3000 (Dionex, Германия) при 210 нм. Тип хроматографической колонки – Acclaim® C18; 3 мкм; 120 Ǻ 2.1×210 мм. В качестве подвижной фазы использовались элюэнты: ацетонитрил (ОСЧ для ВЭЖХ и фосфатный буфер (25 мМ КН2РО4 с рН 3.6). Градуировка прибора проводилась по государственным стандартным образцам (ГСО) карбоновых кислот с чистотой не менее 98% [Руководство по методам…., 2004]. Повторность биохимических анализов – 3-кратная.
Результаты обрабатывали статистически с использованием однофакторного дисперсионного анализа. Рассчитывали среднюю арифметическую величину, ошибку средней арифметической, достоверность различий между вариантами по критерию Стьюдента (р ≤ 0.05).
Результаты и их обсуждение
Важным показателем устойчивости растений к действию неблагоприятных факторов является изменение их роста.
Нами была проанализирована величина прироста проростков за 7 сут. эксперимента (рис. 1). Показано, что прирост в условиях гипертермии (40 ° С, 7 сут.) существенно уменьшался. Предварительное тепловое закаливание снизило рост ингибирующий эффект гипертермии. Холодовое закаливание не оказало заметного влияния на последующий рост проростков в условиях гипертермии. Таким образом, наши результаты свидетельствуют об ограниченных адаптивных возможностях теплолюбивых проростков огурца при холодовом закаливании.
Известно, что при закаливании реализуются разные физиологические программы [Титов, Таланова, 2009]. При физиологически нормальных температурах осуществляется ростовая и онтогенетическая программа. Низкие и высокие закаливающие температуры вызывают своеобразное «генетическое перепрограммирование», при этом реализация ростовой и онтогенетической программ тормозится или полностью блокируется, а жизнеспособность растения в этих условиях, прежде всего, связана с запуском адаптивной программы, направленной на повышение их устойчивости. Повреждающие высокие температуры приводят к структурным и функциональным повреждениям, в результате которых происходит гибель клеток.
Возможно, что закаливание, как краткосрочное повреждающее воздействие, вызывает появление в клетках соединений, которые являются результатом определенных (обратимых) нарушений и одновременно выполняют функции сигнала, запускающего механизмы формирования повышенной резистентности. В отсутствие такого сигнала (при действии более низких температур) адаптивный потенциал растений, очевидно, проявляется не в полной степени.
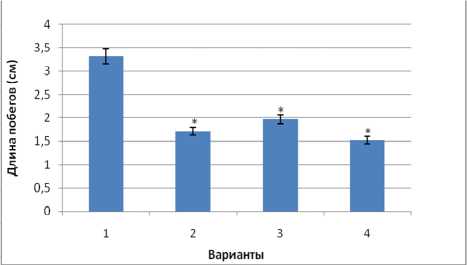
Рис. 1 . Динамика прироста проростков огурца в контрольном и опытных вариантах за 7 сут.: 1 - вариант - контроль (23 ° С, 7 сут.); 2 - вариант - гипертермия (40 ° С, 7 сут.); 3 - вариант - тепловое закаливание (последовательно 30 ° С, 1 сут. и 40 ° С, 6 сут.); 4 – вариант – холодовое закаливание
(последовательно 10 ° С, 1 сут. и 40 ° С, 6 сут.); * различия между вариантами достоверны по критерию Стьюдента (р ≤ 0,05)
В литературе высказывается мнение, что синтез и компартментализация органических кислот в клетке четко регулируются, однако точная оценка величины и состава пулов этих соединений у растений практически отсутствует [Осмоловская, Попова, Кучаева, 2003].
Полагают, что в цитозоле этот пул метаболически активен, относительно стабилен и вследствие нейтральных значений рН представлен не свободными кислотами, а преимущественно их диссоциированными анионами или водорастворимыми калиевыми солями. Вакуолярный пул является временным, динамичным, быстро утилизируемым в темноте для метаболизма в цитозоле и представляет комплексы с катионами кальция и магния [Осмоловская, 2007].
Отсутствие данных по влиянию гипертермии на обмен органических кислот побудило нас провести исследования такого плана.
Из большого разнообразия органических кислот для анализа мы выбрали следующие карбоновые кислоты: щавелевую, лимонную и фумаровую.
В литературе имеются сведения о том, что щавелевая кислота вовлечена в поддержание катионанионного баланса и рН-гомеостаза растений [Осмоловская, 2007]. Известно также, что из-за уве- личения проницаемости мембран при гипертермии катион-анионный гомеостаз нарушается. У растений за эту регуляцию отвечают два механизма: биофизический – электрогенная АТФ-зависимая протонная помпа, с помощью которой ионы водорода выводятся через мембраны наружу против электрохимического градиента, и биохимический – рН-чувствительные процессы карбоксилирования и декарбоксилирования органических кислот, в ходе которых продуцируется или потребляется протон.
Установлена способность многих растений накапливать значительные количества оксалата. Од- нако исследования, направленные на оценку степени его вовлечения в формирование ионного гомеостаза растений, крайне малочисленны, и процессы синтеза щавелевой кислоты в них, как пра- вило, трактуются как вторичные по отношению к минеральному обмену растительного организма [Осмоловская и др., 2002].
Полученные нами результаты показали у кон- трольных растений достаточно высокий уровень свободного оксалата по сравнению с содержанием цитрата или фумарата (рис. 2).
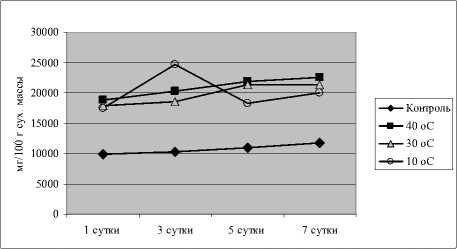
Рис. 2 . Динамика содержания щавелевой кислоты у проростков огурца при действии закаливающих и повреждающих температур
Гипертермия без закаливания вызвала сильное увеличение количества оксалата (в 1.7 раза по сравнению с контролем), которое оставалось стабильно высоким на протяжении всего эксперимента (7 сут.).
Динамика оксалата в вариантах с холодовым и тепловым закаливанием у проростков огурца была разной. Так, тепловое закаливание вызвало резкий подъем количества этой кислоты, а холодовое – понижение по сравнению с контролем.
Таким образом, полученные нами результаты свидетельствуют о вовлечении оксалата в формирование ионного гомеостаза растений в процессе адаптации к гипертермии. Некоторые авторы также приходят к выводу, что контролируемая аккумуляция калиевых солей щавелевой кислоты является доминирующим фактором, определяющим формирование катион-анионного баланса и задающим осмотический потенциал клеток листа [Ruiz, Mansfield, 1994; Franceschi, 2001; Осмоловская, 2007].
Результаты определения цитрата показали, что в норме его уровень в проростках огурца был стабильно высоким в течение 7 сут. эксперимента (рис. 3).
Гипертермия без закаливания, а также гипертермия после теплового закаливания вызвали небольшое увеличение уровня цитрата по сравнению с контролем. При гипертермии после холодового закаливания количество цитрата вначале достоверно снизилось по отношению к контролю, а затем имело тенденцию к возрастанию до уровня контроля.
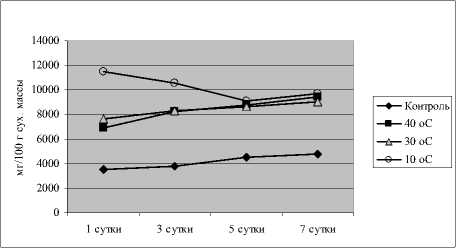
Рис. 3 . Динамика содержания лимонной кислоты у проростков огурца при действии закаливающих и повреждающих температур
Содержание фумарата в норме у исследуемых растений было значительно ниже, чем оксалата и цитрата, что соответствует данным литературы [Землянухин, Землянухин, 1995] (рис. 4).

Рис. 4 . Динамика содержания фумаровой кислоты у проростков огурца при действии
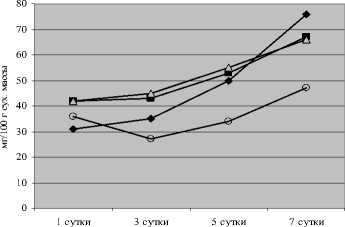
закаливающих и повреждающих температур
Во всех опытных вариантах количество фумарата имело явную тенденцию к снижению по сравнению с контролем, особенно при действии гипертермии после холодового закаливания.
Различия в содержании в растениях отдельных карбоновых кислот следует рассматривать как следствие различий скоростей ферментативных реакций, лежащих в основе их образования и превращения. В этом смысле первостепенную роль играют ферментативные реакции ЦТК (цикл трикарбоновых кислот).
Накопление цитрата и фумарата свидетельствует о превращениях, идущих по ЦТК, причем ги- пертермия оказывает непосредственное влияние на ход таких превращений. Известно, что лимитирующей реакцией всего ЦТК является реакция синтеза цитрата (фермент цитратсинтетаза). В наших исследованиях содержание цитрата при гипертермии значительно повышалось, а фумарата – было на уровне контроля. Последнее может является показателем стабильной скорости ЦТК, которая в норме должна соответствовать скорости гликолиза.
На основании этого можно предположить, что при гипертермии усиливается другая важная функция ЦТК – анаболическая, т. е. промежуточные соединения ЦТК включаются в процессы биосинтеза и, в первую очередь, аминокислот. Последнее, с одной стороны, обеспечивает усиленный белковый синтез (БТШ), а с другой – нейтрализует избыток аммиака.
Имеются данные о том, что некоторые ферменты ЦТК (в том числе цитратсинтетаза) имеют разную компартментализацию в растительной клетке и являются множествеиными молекулярными формами (изоферментами, т.е. кодируются различными генами). Установлено также, что долговременное влияние теплового закаливания может сопровождаться изменениями изоферментных спектров [Martins et al., 1999]. Ферменты трикарбоновых кислот, участвующие в анаболических реакциях, пространственно разобщены от ферментов ЦТК, находятся в цитоплазме и участвуют в синтезе аминокислот. Ферменты же ЦТК, выполняющие энергетические функции, локализованы в митохондриях. Именно участие изоферментов в различных процессах и обусловливает разницу их свойств [Землянухин, Землянухин, 1995].
Оксикислоты могут образовываться не только в ЦТК, но и в результате протеолиза и дезаминирования аминокислот. Например, глутамат может последовательно превращаться в α-кетоглутарат, изоцитрат, цитрат и малат. Повышение содержания большинства кетокислот может вызываться также как снижением активности процессов их прямого аминирования, так и задержкой гликолитического распада сахаров.
На основании полученных нами результатов можно заключить, что тепловое закаливание, как правило, вызывает аккумуляцию щавелевой, лимонной и фумаровой кислот, тогда как холодовое закаливание приводит к падению их уровня. Это подтверждает участие органических кислот в механизме теплового и холодового закаливания растений к действию последующей гипертермии. Вклад каждой из изученных нами органических кислот в процесс закаливания и защитный ответ на действие повреждающей температуры неравнозначен и, возможно, приурочен к различным стадиям адаптационного процесса или является видоспецифичным.
Список литературы Динамика содержания карбоновых кислот в проростках огурца при гипертермии в зависимости от теплового и холодового закаливания
- Балагурова Н.И., Акимова Т.В., Титов А.Ф. Влияние локального охлаждения проростков огурца и пшеницы на различные виды устойчивости листа и корня//Физиология растений. 2001. Т. 48, № 1. С. 113-118.
- Землянухин А.Л., Землянухин Л.А. Метаболизм органических кислот растений: учеб. пособие. Воронеж, 1995. 150 с.
- Кузнецов В.В., Дмитриева Г.А. Физиология растений: учебник. М.: Высш. шк., 2006. 742 с.
- Матухин Г.Р. Физиология приспособления культурных растений к засолению почв. Ростов н/Д: Изд-во Ростов. ун-та, 1963. 340 с.
- Осмоловская Н.Г. Влияние условий минерального питания на формирование пулов оксалата в листьях Amaranthus cruentus L.//Вестник СПб. гос. ун-та. Сер. 3. 2007. Вып. 2, № 11. С. 41.
- Осмоловская Н.Г. и др. Регуляция обмена оксалата в листьях Amaranthus cruentus при разной обеспеченности азотом и катионами//Вестник СПб. гос. ун-та. Сер. 3. 2002. Вып. 2, № 11. С. 41-47.
- Осмоловская Н.Г., Попова Н.Ф., Кунаева Л.Н. Регуляция пулов оксалата в листьях амаранта условиями минерального питания//V съезд общества физиологов растений России: тез. докл. Пенза, 2003. С. 151.
- Руководство по методам контроля качества и безопасности биологически активных добавок к пище. М.: Федеральный центр госсанэпиднадзора Минздрава России, 2004. 240 с.
- Таланова В.В. и др. Раздельное и комбинированное действие засоления и закаливающих температур на растения//Физиология растений. 1993. Т. 40, вып. 4. С. 584-593.
- Титов А.Ф., Таланова В.В. Устойчивость растений и фитогормоны. Петрозаводск, 2009. 206 с.
- Трунова Т.И. Растение и низкотемпературный стресс. М.: Наука, 2007. 55 с.
- Чиркова Т.В. Физиологические основы устойчивости растений. СПб: Изд-во СПб ун-та, 2002. 244 с.
- Чудинова Л.А., Суворов В.И. Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений//Вестник Пермского университета. Сер. Биология. 2011. Вып. 1. С. 17-20.
- Чудинова Л.А., Суворов В.И., Механошина Л.И. Влияние гипертермии на устойчивость кукурузы к последующему засолению//Вестник Пермского университета. 2001. Вып. 4. Биология. С. 85-89.
- Franceschi V.R. Calcium oxalate in plants//Plant Science. 2001. Vol. 6, № 7. P. 331.
- Martins L.L. et al. Electrophoretic changes by heat of isoperoxidase from green bean//Agron. Lusit. 1999. Vol. 47. P. 317-325.
- Ruiz L.P., Mansfield T.A. A postulated role for calcium oxalate in the regulation of calcium ions in the vicinity of stomatal guard cells//New Phytol. 1994. Vol. 127. P. 473-481.
