Динамика травяно-кустарничкового яруса в течение четырёх лет после гибели древостоя ели в очаге поражения короедом-типографом

Автор: Каплевский А.А., Уланова Н.Г.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 4-1 т.27, 2018 года.
Бесплатный доступ
Изучена динамика травяно-кустарничкового яруса в ельнике с сохранённым погибшим древостоем и на вырубке в сравнении с исходным лесом за три года. Исследования встречаемости и ординация растительности показали сходство ельника с погибшим древостоем и исходного леса и отличие этих фитоценозов от вырубки.
Ельник с погибшим древостоем ели, короедник, динамика фитоценозов, мониторинг, короед-типограф
Короткий адрес: https://sciup.org/148313619
IDR: 148313619 | УДК: 581.524.33 | DOI: 10.24411/2073-1035-2018-10104
Текст краткого сообщения Динамика травяно-кустарничкового яруса в течение четырёх лет после гибели древостоя ели в очаге поражения короедом-типографом
Выдвинута гипотеза: сохранение сухостоя ели в очагах усыхания древостоя ели после вспышки численности короеда-типографа позволяет сохранить лесное сообщество, близкое к исходному. В образовавшемся сообществе динамика фитоценоза будет направлена на восстановление лесного сообщества уже в первые годы после гибели древостоя.
Цель проведенных исследований – выявление особенностей изменения структуры травяно-кустарничкового яруса и мохового покрова ельника после гибели древостоя в сравнении с фитоценозами после вырубки сухостоя и исходным лесом.
ОБЪЕКТЫ И МЕТОДЫ
На территории Звенигородской биостанции МГУ (Московская область) в 2013 г. рядом заложены три постоянные пробные площади (ППП) одинакового размера (800 м2) в ельнике зеленчуковом: с погибшим в 2012 г. древостоем ели (короедник), на сплошной вырубке сухостоя ели зимой 2012-2013 гг. и с живым древостоем ели (контроль). Исследования проведены в августе 2014, 2015, 2016 и 2017 гг. по единой методике. На ППП заложены по три трансекты длиной 40 м и шириной 40 cм. На каждом мет- ре трансект изучена корневая встречаемость видов травяно-кустарничкового яруса (ТКЯ). Для её определения использована жесткая рамка размером 40×100 см, разделённая съёмными нитями на квадраты 20×20 см.
Проведена ординация описаний (встречаемости всех видов ТКЯ и мхов) площадок размером 0,4 м2 методом DCA по трем ППП для трех трансект с 2014 по 2017 г. Ординация проведена с помощью программы PCord. Сравнительный анализ ТКЯ и мхов с контролем и между площадями двух технологий уборки сухостоя проведен с использований статистических методов.
Идентификация осей ординации проведена при помощи индикационных шкал Г. Эллен-берга с использованием балловых характеристик 36 видов по 4 факторам среды (Уланова, Жмылев, 2014) согласно принятой методики (Уланова, 1995).
Для общей оценки сходства травянокустарничкового яруса и мохового покрова исследуемых фитоценозов в 2014-2017 году был вычислен коэффициент сходства Жаккара
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Доля видов, характерных для исходного ельника зеленчукового в 2014 году, были в ко-роеднике в 2 раза больше, чем на вырубке. На площадке, где древостой был вырублен и вывезен, больше половины присутствующих видов появились после вырубки древостоя, видовой состав травяно-кустарничкового яруса существенно отличался от такового в контроле.
Для короедника и контрольной площадки в 2014 году значение коэффициента Жаккара было максимальным (табл. 1). В 2015 - 2017 году произошло уменьшение сходства травянокустарничкового яруса и мохового покрова этих пробных площадей. При этом их сходство каждый год было больше, чем у вырубки и контроля, короедника и вырубки.
Коэффициент сходства вырубки и контроля в течение всего периода исследований оставался стабильным, сходство вырубки и контроля было меньше, чем сходство вырубки и короед-ника, короедника и контроля.
Таблица 1 Коэффициенты сходства (Жаккара) растительности короедника, вырубки и контроля в 2014-2017 гг.
|
Фитоценозы |
2014 |
2015 |
2016 |
2017 |
|
Короедник-контроль |
0,67 |
0,53 |
0,48 |
0,47 |
|
Вырубка-контроль |
0,38 |
0,35 |
0,35 |
0,36 |
|
Короедник-вырубка |
0,46 |
0,51 |
0,37 |
0,35 |
Коэффициенты сходства между изученными фитоценозами уменьшались от максимума в начале исследования к минимуму на 4 год наблюдений. Сходство вырубки и контроля оказалось стабильно низким все годы, так как на вырубке образовалось луговое сообщество, принципиально отличное от лесного. Некоторое увеличение сходства короедника и вырубки в 2015 г. можно связать с переходом в валёж погибших в результате вспышки короеда-типографа елей, а уменьшение – с последующим разрастанием лещины. В 2017 году коэффициенты сходства вырубки и короедника и вырубки и контроля стали близки.
Ценотический спектр видов травянокустарничкового яруса в короеднике в 2017 г. схож со спектром контрольной площади, изменяются только соотношения групп (рис. 1). На обеих площадях преобладают лесные виды ( Asarum europaeum , Oxalis acetosella, Galeobdolon luteum , Pulmonaria obscura , Stellaria holostea и др.). Кроме собственно лесных, на площадках в контроле и короеднике были отмечены растения из групп: лесолуговые ( Ajuga reptans , Equisetum pratense , Luzula pallescens ), сорные ( Geum urbanum , Impatiens parviflora .), сорно-лесные ( Rubus idaeus ). Произошло увеличение доли сорно-лесных ( Urtica dioica ) и лесолуговых (Galium mollugo ) видов. Увеличение вклада сорных видов связано с исчезновением части лесных видов, в частности Athyrium filix-femina . В 2015 году в короеднике появился луговой вид (Campanula patula ). Появление различий короедника и контроля по соотношению ценотических групп можно объяснить повышением освещённости в короедни-ке.
Если по составу эколого-ценотических групп видов короедник и контроль сходны, то вырубка резко отличается от них числом и составом эколого-ценотических групп. Значительные изменения флористического состава вырубки по сравнению с контролем произошли в результате гибели ТКЯ, нарушений мохового и почвенного покрова при вывозе древесины, сжигании рубочных остатков и последующего вселения новых видов. На вырубке происходит вселение луговых (например, Centaurea scabiosa) и сорно-луговых (Calamagrostis epigeios, Taraxacum officinale) видов, в два раза увеличивается доля сорных видов за счёт Galeopsis bifida, Cirsium vulgare, Erigeron an-nuus, Sonchus vulgaris и других. Суммарная доля собственно лесных видов уменьшается. При этом появляются виды, не встречавшиеся в контроле: лесные (Paris quadrifolia) и сорно- лесные виды (Chamerion angustifolium), Moehringia trinervia, Solidago virgaurea).
Проведено исследование встречаемости видов ТКЯ на площадках 1x0,4 м. Различия средней встречаемости наиболее распространённых видов значительны (рис. 2). Анализ проведен для всех видов, чья встречаемость была более 1 % на каждой постоянной пробной площади.
В 2017 году в короеднике значимо большую встречаемость, чем в контроле, имеют Aegopodium podagraria и Stellaria holostea, а значимо меньше встречаемость Asarum euro-paeum и Oxalis acetosella . Наибольшая встречаемость из трех площадок в короеднике отмечена для Stellaria holostea и Pulmonaria obscura.
ТКЯ короедника богаче видами, чем контрольная площадь. После гибели древостоя появились светолюбивые виды: Chrysosplenium alternifolium, Galium mollugo, Luzula pilosa и другие. Отмечена значительная локальная вариабельность встречаемости видов, выражена мозаичность, связанная с гетерогенностью погибшего и выжившего древостоя и разрастанием кустов лещины. Пятна мозаик с высокой встречаемостью образуют Galeobdolon luteum, Asarum europaeum , Aegopodium podagraria , Sta-chys sylvatica, Stellaria holostea. Присутствие сорных видов говорит о значительной рекреационной нагрузке на этот участок.
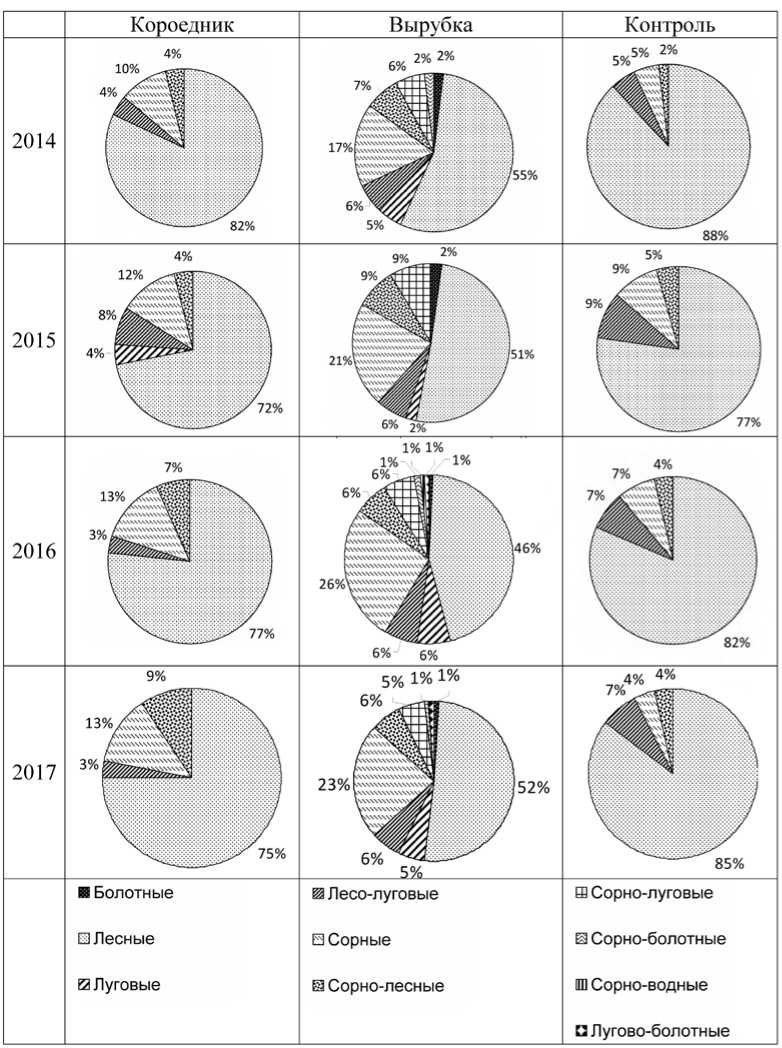
Рис. 1. Ценотические спектры видов травяно-кустарничкового яруса постоянных пробных площадях в 2014-2017 гг.
Galeobdolon luteum
Galeobdolon luteum
Equisetum pretense
Stellaria holostea
Equisetum protense
Stellaria holostea
Impatiens parviflora
Stellaria nemorum
Impatiens parviflora
Stellaria nemorum
Stachys sylvatica
Urticadioica
Stachys sylvatica
Urticadioica
Asorumeuropaeum
Aegopodium podagrorio Oxolisacetosella
Asorumeuropaeum
Aegopodium podagrorio
Oxalis acetosello
Ajuga reptons Dryopteris carthusiana Pulmonarioobscuro
Ajuga reptans Dryopteris carthusiana Pulmonarioobscuro
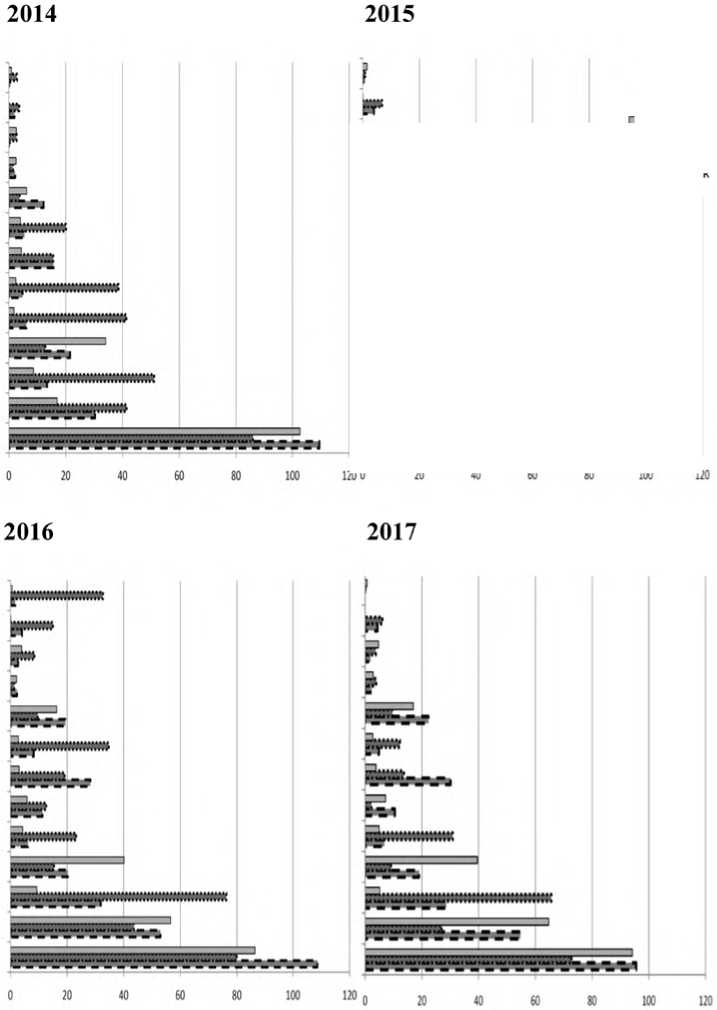
Рис. 2. Встречаемость наиболее распространённых видов ТКЯ на площадках 0,4 м2 (встречаемость более 1% на каждой пробной площади) в короеднике, на вырубке и в контроле в 2014-2017 гг.
Схожие изменения во встречаемости основных видов травяно-кустарничкового яруса после гибели древостоя ели отмечены также для лесов Чехии (Jonášová, Prach, 2008). Вместе с Oxalis acetosella происходило распространение Trientalis europaea и Homogyne alpina. При исследовании горных ельников зеленчуковых в Швейцарии выявлено разрастание Chamerion angustifolium, Dryopteris dilatata , D. carthusiana, Athyrium filix-femina (Kupferschmid et al., 2005) .
Отличия травяно-кустарнчкового яруса вырубки от леса гораздо более значительны чем в короеднике. Резкое изменение видового состава и экологических условий привело к росту числа видов почти в два раза. Произошло увеличение числа сорных видов. Велика гетерогенность экологических условий, связанная с разной степенью нарушенности почвенного покрова, куртинами подроста и подлеска, кострищами, приствольными повышениями у пней, западин, завалов порубочных остатков и мелких веток. Поэтому явно выражена мозаичность ТКЯ с пятнами 100% встречаемости одного из видов: Aegopodium podagraria, Stellaria nemorum, Stellaria holostea, Rubus idaeus, Galeobdolon luteum, Impatiens parviflora. Светолюбивые и нитрофильные виды имеют наибольшую встречаемость на вырубке (Aegopodium podagraria, Impatiens parviflora, Equisetum pratense). Однако и теневыносливый Oxalis acetosella успешно разрастается на вырубке под пологом из Stellaria nemorum и Galeobdolon luteum. Для этого вида в Чехии была отмечена другая тенденция – сокращение распространения на вырубках (Jonášová, Prach, 2008).
На вырубке значимо меньше, чем в контроле встречаемость Oxalis acetosella, Asarum europaeum и Pulmonaria obscura . Другие виды на вырубке имеют значимо большую встречаемость, чем в короеднике и контроле. Наибольшая встречаемость из трёх площадок в этом фитоценозе отмечена для Aegopodium podagraria , Impatiens parviflora , Equisetum pratense, Oxalis acetosella , Stellaria nemorum , Urtica dioica и Stachys sylvatica . Значительные отличия при сравнении ТКЯ вырубки и короед-ника выявлены также для ельников Чехии (Jonášová, Prach, 2008). Активно распространяются луговые и сорно-луговые виды Centaurea scabiosa, Calamagrostis epigeios , Taraxacum officinale и другие.
В травяно-кустрничквовм ярусе ненарушенного леса доминируют Galeobdolon luteum, Asarum europaeum и Oxalis acetosella . Покрытие площади видами ТКЯ относительно однородно, не отмечено выраженной мозаичности. Однако исследованный фитоценоз испытывает значительную рекреационную нагрузку, о чём говорит присутствие сорных видов.
Сравнение встречаемости видов в 2014-2017 году (рис. 1) показало, что в короеднике значимых изменений немного. Из-за небольшого повышения освещенности и влажности, повысилась встречаемость Stellaria holostea и Aegopo-dium podagraria .
На вырубке в 2016 и 2017 году в сравнении с 2015 снижается встречаемость Aegopodium po-dagraria, Stellaria nemorum и Impatiens parviflo-ra. Изменения такого рода связаны с разрастанием на вырубке малины и сохранившихся кустов лещины.
Встречаемость Stachys sylvaticа в этом фитоценозе в 2016 году выросла в 12 раз, но изменения оказались не значимыми, т.к. куртины были приурочены лишь к отдельным площадкам. В 2017 году встречаемость Stachys sylvaticа вновь значительно снизилась.
На контрольной площадке происходит увеличение встречаемости Oxalis acetosella и уменьшение Galeobdolon luteum .
Гибель древостоя не привела к существенным изменениям мохового покрова короедни-ка. Уменьшение встречаемости большинства видов мхов в короеднике связано с уменьшением освещённости за счёт зарастания окон лещиной и разрастания ТКЯ. В ельниках Чехии в короедниках не происходит вселения новых видов (Jonášová, Prach, 2008).
На вырубке отмечено в полтора раза больше видов мхов, по сравнению с контролем и коро-едником. Именно разнообразие микросайтов с экологически разными условиями создает условия для появления многих видов мхов. Центрами биоразнообразия оказались кострища, образованные при сжигании рубочных остатков. Однако, встречаемость видов мохового покрова на вырубке ниже, чем в короеднике. Это связано с повышенной конкуренцией со стороны растений травяно-кустарничкового яруса на вырубке. Исключением из этого правила являются Eurhynchium angustirete и Plagiomnium affine , для которых наибольшая встречаемость отмечена на вырубке. Эти виды приурочены к нарушенным микросайтам, медленно зарастающих травой. Для Швейцарии отмечено увеличение на вырубке встречаемости Hylocomnium splendens , Thuidium tatarisci-num, Dicranium scoparium (Kupferschmid et al., 2005). Нами отмечены новые пионерные виды мхов Ceratodon purpureus, Funaria hygrometrica, Polytrichum juniperinum на кострищах и участках с нарушенной почвой. Присутствие этих видов характерно и для вырубок на месте ельников Чехии (Jonášová, Prach, 2008).
При сравнении ординационных диаграмм за четыре последовательных года исследований можно отметить, что зона наибольшей плотности площадок ельника с погибшим древостоем ели в 2016-17 году размывается, но не совпадает ни вырубкой, ни с контрольной площадью (рис. 3). Такое изменение положения площадок говорит о самобытности развития фитоценоза ельника после гибели ели.
Площадки короедника в 2014 году образуют компактное скопление (рис. 3), с ходом развития фитоценоза разброс площадок на ордина-ционной диаграмме увеличивается, что говорит о повышении гетерогенности микроусловий. Это повышение связанно с ветроломом уничтоженных короедом елей, приводящим к повреждению подлеска и образованием микросайтов расположенных под нависающими на небольшой высоте над землёй упавшими стволами и местам опадения ветвей, образующих в некоторых местах сплошной покров. После зарастания образовавшихся при повреждении подлеска окон образуется новая мозаика освещенности, связанная с разрастанием кустов лещины.
Площадки вырубки в 2014 году образуют на ординационной плоскости отдельное крупное скопление, не пересекаются со скоплениями ельника с погибшим древостоем ели и нетронутого леса. Растительность вырубки более разнообразна, образует широкую область с большим числом отдельно расположенных участков. На вырубке выражена мозаичность ТКЯ, связанная с гетерогенностью экологических условий и разной степенью нарушенности почвы. Сходные исследования динамики ТКЯ и мохового покрова в лесах Чехии также выявили различия в динамике фитоценозов вырубки и короедника (Jonášová, Prach, 2008).
Короеднк Вырубка Контроль
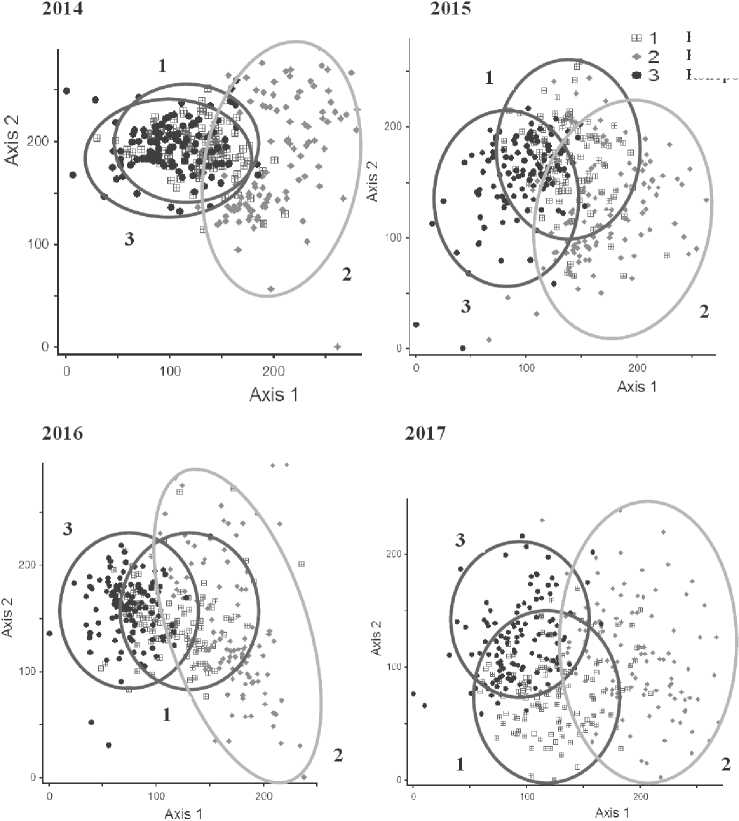
Рис. 3. Ординация растительности (травяно-кустарничкового яруса и мохового покрова) площадок трех пробных площадей (контроль, короедник и вырубка) в 2014-2017 гг.
В ходе развития сообщества вырубки в 2014-2017 году разнообразие микросайтов сохраняется и увеличивается. При этом, частично происходит смена видов (место Impatiens parvi-flora занимает Rubus idaeus и Aegopodium po-dagraria , место Stellaria nemorum - густой подрост осины и ели), образующих пятна мозаики, но их разнообразие сохраняется.
Зона локализации площадок контроля на протяжении четырёх лет исследования сохраняется стабильной и значительно не размывается. Стабильность положения площадок на ор-динационной диаграмме свидетельствует отсутствие динамических процессов в фитоценозе ельника зеленчукового.
ЗАКЛЮЧЕНИЕ
В травяно-кустарничковом ярусе короедни-ка виды сохранили свое доминирование после гибели древостоя ели. Появились новые светолюбивые виды с небольшой встречаемостью. На вырубке сухостоя ели произошли кардинальные изменения травяно-кустарничкового яруса, количество видов увеличилось в 2 раза. Доминирование перешло к другим видам. Значительные нарушения напочвенного покрова привели к высокой мозаичности травянокустарничкового яруса.
Спектры ценотических групп растений травяно-кустарничкового яруса в короеднике и исходном фитоценозе близки, преобладают лесные виды. Присутствие сорных и сорнолесных видов связано с большой рекреационной нагрузкой в лесу. На вырубке доля лесных видов значительно сокращена, возросла доля луговых и сорно-луговых. Фитоценоз вырубки можно отнести к лесо-луговому типу.
Ординация всех описаний площадок методом DCA выявила близость растительности ко-роедника и ненарушенного ельника. Площадки вырубок удалены от скоплений площадок коро-едника и контроля. За три года после вырубки леса возник другой луговой фитоценоз с выраженной мозаичностью.
Общая динамика развития фитоценоза коро-едника направлена на восстановление сообщества, близкого к исходному. Большинство происходящих изменений в фитоценозе с сохранённым сухостоем ели носят количественный, а не качественный характер. Естественный ход восстановления сообщества ведет к формированию устойчивого широколиственного леса уже в первые годы после гибели елей. Зарастание вырубки приводит к образованию мозаичного сообщества, значительно отличающегося от исходного.
Список литературы Динамика травяно-кустарничкового яруса в течение четырёх лет после гибели древостоя ели в очаге поражения короедом-типографом
- Ермаков А.Л., Маслов А.А. Породный состав естественного возобновления в очагах усыхания ели от короеда типографа в Московской области // Изв. Самар. НЦ РАН. 2012. Т. 14, № 1(5). С. 1236-1238.
- Малахова Е.Г., Лямцев Н.И. Распространение и структура очагов усыхания еловых лесов Подмосковья в 2010-2012 годах // Изв. Санкт-Петербургской лесотехнической академии. 2014. Т. 207. С. 193-201.
- Маслов А.Д. Комарова, И.А., Котов А.С. Динамика размножения короеда-типографа в Центральной России в 2010-2013 гг. и прогноз на 2014 г. // Лесохоз. информация. 2014. Т. 1. С. 38-46.
- Маслов А.Д., Комарова И.А., Котов А.С. Состояние и динамика очагов размножения короеда-типографа в Центральной России в 2010 и первой половине 2011 г. // Лесохоз. информация. 2011. Т. 1. С. 39-46.
- Уланова Н.Г. Математические методы в геоботанике. М.: Изд-во МГУ, 1995. 109 с.
- Уланова Н. Г. Восстановительная динамика растительности сплошных вырубок и массовых ветровалов в ельниках южной тайги (на примере европейской части России): автореф. дис.. докт. биол. наук. М., 2006. 46 с.
- Уланова Н.Г., Жмылёв П.Ю. Эколого-ценотический анализ растительных сообществ. Учебное пособие. М.: МАКС Пресс, 2014. 80 с.
- Уланова Н.Г., Маслов А.А., Синичкина Д.С. Лесовосстановление на шестой год после усыхания ели в ельнике-кисличнике // Тр. Звенигор. биол. станции. 2011. Т. 5. С. 152-157.
- Jonášová M., Prach K. The influence of bark beetles outbreak vs. salvage logging on ground layer vegetation in Central European mountain spruce forests // Biological conservation. 2008. V. 141. P. 1525-1535.
- Kupferschmid A.D., Bugmann H. Predicting decay and ground vegetation development in Picea abies snag stands // Plant Ecology. 2005. V. 179. P. 247-268.
