Динамика углевод - ферментного комплекса проростков ржи при адаптации к ионам свинца после теплового закаливания и без него

Автор: Чудинова Лариса Алексеевна, Конова Дина Радиковна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2020 года.
Бесплатный доступ
Изучали динамику содержания глюкозы, активности амилазы и инвертазы, а также интенсивность фотосинтеза в процессе адаптации проростков ржи к свинцовому стрессу (1 мМ раствор ацетата свинца) в условиях наличия или отсутствия теплового закаливания (+40°С, 3 ч.). Установили значительное снижение уровня обоих фондов глюкозы (компартментированного и свободного), вызываемое как негативным воздействием свинца на интенсивность фотосинтеза, так и усилением оттока сахаров из листьев в корни в результате снижения активности инвертазы. Другой возможной причиной можно считать высокую степень метаболизации глюкозы, на что косвенно указывает рост активности амилазы. Предполагается, что в реализации механизма теплового закаливания участвуют оба фонда глюкозы (свободный и компартментированный), что обеспечивает сочетание повышенного уровня жизнедеятельности и развитие металлоустойчивости растений.
Проростки ржи, ацетат свинца, тепловое закаливание, глюкоза, интенсивность фотосинтеза, активность амилазы и инвертазы
Короткий адрес: https://sciup.org/147229638
IDR: 147229638 | УДК: 581.1:632.122.1 | DOI: 10.17072/1994-9952-2020-1-63-69
Dynamics of the carbohydrate-enzyme complex of rye seedlings when adapting to lead ions after heat hardening and without it
We studied the dynamics of glucose content, the activity of amylase and invertase, as well as the intensity of photosynthesis in the process of adaptation of rye seedlings to lead stress (1 mM solution of lead acetate) in the presence or absence of heat hardening (+ 40 ° С, 3 h). We found a significant decrease in the level of both glucose funds (compartmentalized and free), caused both by the negative effect of lead on the intensity of photosynthesis and by an increase in the outflow of sugars from leaves to the roots as a result of a decrease in invertase activity. Another possible reason is a high degree of glucose metabolism, as indirectly indicated by an increase in amylase activity. It is assumed that both glucose funds are involved in the implementation of the thermal hardening mechanism, which provides a combination of an increased level of vital activity and the development of metal resistance of plants
Текст научной статьи Динамика углевод - ферментного комплекса проростков ржи при адаптации к ионам свинца после теплового закаливания и без него
Тяжелые металлы (в первую очередь ртуть, кадмий и свинец) относятся к числу наиболее опасных химических загрязняющих веществ, которые даже в небольших количествах способны оказывать токсическое влияние на различные организмы, в том числе на растения. В силу этого ведутся активные исследования металлоустойчивости растений, однако полученные сведения зачастую носят противоречивый характер. Кроме того, такие работы в основном касаются дикорастущих видов растений, тогда как культурные виды изучены в гораздо меньшей степени [Yang et al., 2005; Титов, Казнина, Таланова, 2014].
Недостаточно сведений и о закаливающем дей- ствии повышенной температуры на развитие ме-таллоустойчивости растений, хотя во многих исследованиях отмечается протекторный эффект кратковременного теплового шока к последующему тепловому, холодовому, солевому воздействию и водному дефициту [Титов, Таланова, 2009].
Важная роль углеводного обмена для процессов адаптации растений к различным стрессорам в настоящее время не вызывает сомнения. Однако направленность превращений углеводноферментного комплекса в условиях стресса до настоящего времени остается предметом дискуссии. Одни авторы считают, что это результат деструктивных процессов, связанных с повреждением клеток, другие – одна из составляющих ком-
плекса защитных реакций на стресс [Сабельникова, 2007; Трунова, 2007]. Следует отметить, что наибольшее количество экспериментальных данных по углеводному обмену получено при изучении гипотермии. В то же время исследованию роли данного обмена в формировании металлоустой-чивости уделялось значительно меньше внимания.
Сформировалось мнение, что усиление гидролиза полимерных форм углеводов, в том числе и низкомолекулярных олигосахаридов, является неспецифической ответной реакцией растений на неблагоприятные воздействия различной природы [Карпец, Колупаев, 2009]. Но четкие экспериментальные доказательства связи активности гидролитических ферментов с формированием металло-устойчивости растений на сегодняшний день отсутствуют.
Следует также отметить, что в большинстве работ изучалась динамика концентрации углеводов в растительных клетках в состоянии стресса, но не учитывалась их компартментация (или разделение на отдельные фонды). Необходимость компарт-ментирования низкомолекулярных сахаров определяется их полифункциональностью на уровне клетки. Проявляется компарментация в том, что в определенных физиологических или биохимических процессах участвует не вся масса внутриклеточного вещества, а только некоторая его часть. Иными словами – в разной доступности разных фондов для определенных функций, что обеспечивается наличием внутриклеточных мембран. Не исключена, хотя и имеет меньшее значение, ком-партментация, основанная на различии свойств отдельных зон цитозоля в пределах клетки [Холодова, 1978]. Проведенные нами ранее исследования позволили сделать предположение, что изменение компартментации глюкозы и сахарозы, является условием реализации защитных функций этих моносахаридов в условиях стресса [Чудинова, Чернышева, 2017].
Все вышесказанное даёт основание для изучения динамики свинцовой устойчивости проростков ржи, содержания в них свободного (СФ) и ком-партментированного (КФ) фондов глюкозы и активности ключевых гидролитических ферментов углеводного обмена (амилазы и инвертазы) в условиях наличия или отсутствия предобработки повышенной температурой, что и является целью данной работы.
Объект и методы исследований
Исследования проводили на 10-дневных проростках ржи посевной (озимая форма, сорт Вятка 2), которые выращивали на вермикулите в климатической камере ШН-М при температуре 20°C, влажности 75% и освещенности люминесцентными лампами 16 ч. в сутки (1-й вариант – контроль).
Затем проростки подвергали стрессовым воздействиям по следующей схеме: 2 вариант – экспозиция на 1 мМ растворе ацетата свинца; 3 вариант – предобработка 3 ч. при +40°C, затем экспозиция на 1 мМ растворе ацетата свинца.
Анализировали интенсивность роста по длине побега и его сырой массе на 3-и и 8-е сут. свинцового стресса (повторность 20-кратная); содержание СФ глюкозы (экстракция из гомогената 10 мин. при 80°С) и КФ глюкозы (экстракция из гомогената 50 мин. при 80°С) определяли по методу Вознесенского [Большой практикум …, 2012] (повторность 3-кратная); интенсивность фотосинтеза определяли фотоколориметрическим методом по содержанию углерода в листьях [Аликов, 1983] (повторность 3-кратная).
Результаты обрабатывали статистически, с использованием однофакторного дисперсионного анализа в программе Stadia. Определяли достоверность различий между вариантами по наименьшей существенной разнице (d > НСР 0.05 ).
Результаты и их обсуждение
Рост является интегральным показателем, отражающим степень адаптации растения к окружающей среде, а торможение ростовых процессов – это один из ранних признаков повреждающего действия неблагоприятных факторов на растения. Нами были проанализированы два параметра, характеризующих интенсивность роста: линейные размеры и сырая масса побегов ржи (рис. 1).
Полученные результаты показали заметное ингибирование накопления сырой биомассы побегов ржи после 3-суточного роста на 1мМ растворе ацетата свинца и последующую остановку роста проростков на 8-е сут. в этих же условиях по сравнению с контролем. Предварительное тепловое воздействие (+40оС, 3 ч.) заметно уменьшило рост ингибирующий эффект свинца (рис.1, А). Сходные тенденции (но с меньшей амплитудой) наблюдали и в отношении динамики длины побегов (рис. 1, Б).
Токсическое действие тяжелых металлов на растения основано на следующих процессах: вмешательство в функционирование ферментных систем; нарушение баланса элементов питания; изменение мембран, приводящее к нарушению ближнего и дальнего транспорта [Титов, Казнина, Таланова, 2014].
Полученные нами данные вполне согласуются с имеющимися в литературе сведениями о повышении металлоустойчивости растений после непродолжительной тепловой предобработки [Титов, Таланова, 2009; Чудинова, Чернышева, 2017]. Можно предположить, что это является результатом теплового закаливания, которое, как правило, сопровождается изменениями термостабильности белков и определенных функций клеток, в резуль- тате чего активируются защитно-приспособительные процессы. Многие авторы положительное влияние теплового закаливания на последующую устойчивость растений к разным стрессорам связывают с индукцией синтеза белков теплового шо- ка и с их активным участием в защите клетки от токсичного действия тяжелых металлов [Suzuki, Koizumi, Sano, 2001; Карпец, Колупаев, 2009; Титов, Казнина, Таланова, 2014].
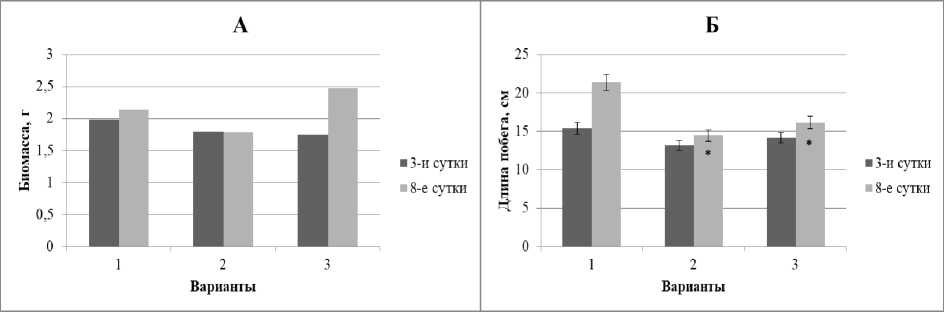
Рис. 1 . Динамика биомассы (А) и длины побегов (Б) проростков ржи при свинцовом стрессе в условиях предобработки повышенной температурой и без нее
Здесь и далее: 1 – контроль; 2 – экспозиция на1 мМ растворе ацетата свинца; 3 – экспозиция 3 ч. при 40°C, затем на 1 мМ растворе ацетата свинца. * - различие с 1-м вариантом достоверно; # - различие со 2-м вариантом достоверно (d > НСР 0.05 )
Не вызывает сомнения, что адаптация растений к действию тяжелых металлов представляет собой сложный многокомпонентный процесс, включающий различные физиолого-биохимические механизмы, которые, дополняя друг друга, обеспечивают определенный уровень металлоустойчивости, зависящий от генотипа [Титов, Таланова, 2009].
Полученные нами ранее результаты показали участие обоих фондов (СФ и КФ) глюкозы и саха- розы в формировании устойчивости овса к кадмиевому стрессу [Чудинова, Чернышева, 2017]. Было сделано предположение, что СФ растворимых сахаров участвует в активном метаболизме, а КФ – в формировании кадмиевой устойчивости.
Как видно из рис. 2, у контрольных проростков ржи количество обоих фондов глюкозы было примерно одинаковым и достаточно стабильным в течение всего эксперимента.
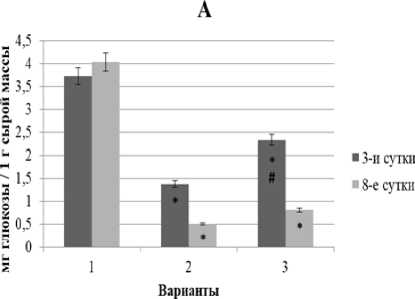
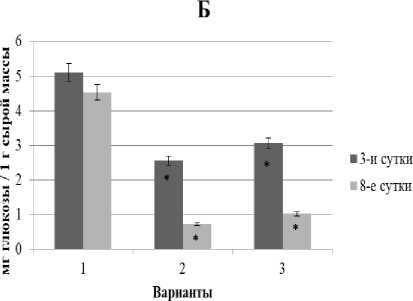
Рис. 2. Динамика содержания СФ (А) и КФ (Б) глюкозы в проростках ржи при свинцовом стрессе в условиях предобработки повышенной температурой и без нее
В условиях монодействия свинца (рис. 2, 2-й вар.) наблюдалось существенное уменьшение уровня глюкозы на 3-и сут. воздействия до 37% (СФ) и 27% (КФ) от контроля. По мере увеличения времени экспозиции до 8 сут. содержание обоих фондов глюкозы падало до 13‒16% от контроля.
При комбинировании теплового закаливания и последующей экспозиции в растворе свинца (рис. 2, 3-й вар.) содержание обоих фондов глюкозы также падало, однако амплитуда снижения была значительно меньше (до 63% от контроля при 3-суточной экспозиции и до 23% от контроля при 8- суточной экспозиции), чем в варианте с монодей- ляются составной частью механизма теплового за- ствием свинца.
На основании полученных результатов можно предположить, что рост, ингибирующий эффект свинца, связан с существенным снижением уровня СФ и КФ глюкозы, особенно в период специализированной адаптации (8-суточный стресс). Возможно, это определяется интенсивным расходом глю- козы на поддержание структурного и энергетического метаболизма в условиях свинцового стресса. Так, уменьшение содержания растворимых сахаров некоторые авторы объясняют расходованием глюкозы на перехват активных форм кислорода и на включение альтернативных путей дыхания [Дерябин и др., 2007].
Поскольку тепловая предобработка примерно в каливания, направленного на развитие адаптационных процессов, обеспечивающих повышение металлоустойчивости растений. В большей степени положительный эффект закаливания проявился при 3-суточном свинцовом стрессе, тогда как при более длительном воздействии свинца (8 сут.) – был существенно слабее.
Внутриклеточная концентрация растворимых сахаров у растений определяется главным образом интенсивностью фотосинтеза, гидролизом запасных полисахаридов, а также скоростью их оттока из листьев в нефотосинтезирующие органы.
В связи с этим одной из наших задач было изучение интенсивности фотосинтеза проростков. Полученные результаты (рис. 3) показали существен- одинаковой степени влияла на СФ и КФ глюкозы, а именно обеспечивала более высокий их уровень ное снижение этого показателя в условиях свинцо- вого стресса (особенно на 8-е сут. воздействия), в условиях последующего свинцового стресса, то которое, однако, было достоверно меньше в вари- можно предположить, что оба фонда глюкозы яв- анте с предварительным тепловым закаливанием.
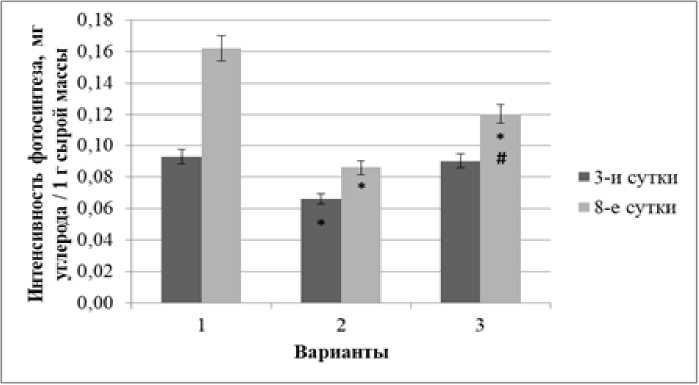
Рис. 3 . Динамика интенсивности фотосинтеза в проростках ржи при свинцовом стрессе в условиях предобработки повышенной температурой и без обработки
Негативное действие тяжелых металлов на фотосинтез проявляется в нарушении ультраструктуры хлоропластов, уменьшении содержания фотосинтетических пигментов, ингибировании активности ферментов цикла Кальвина, а также в замедлении скорости электронного транспорта и поступления в клетки СО 2 . Причем степень торможения фотосинтеза во многом зависит от концентрации металла в окружающей среде [Титов, Каз-нина, Таланова, 2014].
Содержание растворимых сахаров в условиях воздействия стрессоров различной природы, по мнению некоторых авторов [Трунова, 2007], в значительной степени определяется активностью гидролитических ферментов – амилазы и инвертазы. Амилолитическая активность определяет уровень пластидного крахмала, а также крахмала амилопластов, выполняющих роль своеобразного депо глюкозы. Инвертаза катализирует необратимое расщепление сахарозы на глюкозу и фруктозу, которые затем используются в процессе метаболизма.
В растениях было зафиксировано как увеличение, так и падение активности гидролитических ферментов в ответ на стрессы различной природы, интенсивности и продолжительности (гипертермию, засоление, обезвоживание, раневые воздействия, заражение патогеном). При этом, как отмечают авторы [Сабельникова, 2007; Синкевич и др., 2008], изменение активности амилазы и инвертазы может происходить быстро за счет регуляции каталитической активности уже существующих молекул фермента в результате взаимодействия с гетерогенными белками-ингибиторами, изменения изоферментного состава, а также вследствие взаимного перехода растворимой и связанной форм. Нами была исследована динамика активности амилазы и инвертазы в проростках ржи при монодействии ацетата свинца в условиях от- (рис. 4). сутствия или наличия тепловой предобработки
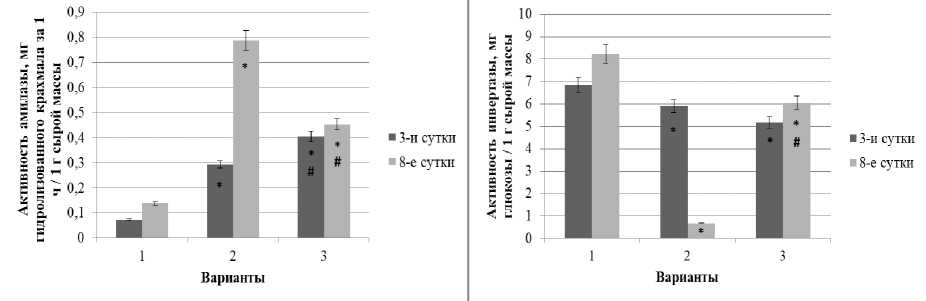
Рис. 4 . Динамика активности амилазы (А) и инвертазы (Б) в побегах ржи при свинцовом стрессе в условиях предобработки повышенной температурой и без нее
Полученные результаты показали рост активности амилазы в условиях свинцового стресса, усиливающийся по мере длительности воздействия (рис. 4, А). При комбинировании теплового закаливания и ацетата свинца повышенный уровень активности амилазы сохранился, но был достоверно ниже, чем в варианте с монодействием свинца.
Поскольку наблюдаемое нами увеличение активности амилазы при свинцовом стрессе не приводило к росту содержания глюкозы, то можно предположить изменение в этих условиях соотношения между изоформами амилазы в сторону увеличения доли β-амилаз, которые относят к стресс-индуцируемым белкам. Результатом гидролиза крахмала под действием β-амилаз является накопление мальтозы, а не глюкозы [Даффус, Даффус, 1987].
Было установлено снижение активности инвертазы в побегах опытных вариантов, причем более сильное в условиях монодействия свинца по сравнению с контролем, и менее сильное в варианте с тепловым закаливанием (рис. 4, Б). Можно предположить, что пониженная активность инвертазы, которая сопровождалась снижением уровня глюкозы, указывает на возможную загрузку флоэмы и транспорт сахарозы в корни в условиях свинцового стресса, что, вероятно, имеет адаптивное значение. В литературе уже указывалось, что процессы оттока сахаров из листьев в корни играют значительную роль в адаптации теплолюбивых растений к низким положительным температурам [Попов, 2002].
Резюмируя полученные нами данные, следует отметить, что у проростков ржи примерно в одинаковом соотношении формируются два фонда глюкозы – свободный, предположительно участвующий в активном метаболизме, и компартментиро-ванный, предположительно участвующий в формировании устойчивости. Адаптация проростков к свинцовому стрессу сопровождается значительным снижением уровня обоих фондов глюкозы. Основной причиной этого следует рассматривать показанное нами отрицательное воздействие свинца на интенсивность фотосинтеза. Другой возможной причиной можно считать высокую степень метабо-лизации глюкозы, ее непосредственное участие в формировании устойчивости. Об этом косвенно свидетельствует повышенная активность амилазы у проростков, подвергнутых воздействию свинца. Снижение активности инвертазы в этой ситуации, вероятно, должно усиливать отток сахаров из листьев в корни, что также является адаптивной реакцией.
Предварительное тепловое закаливание ослабляет негативный эффект свинца на все изучаемые нами показатели обмена глюкозы. Можно предположить, что в реализации механизма теплового закаливания участвуют оба фонда глюкозы (СФ и КФ), что обеспечивает сочетание повышенного уровня жизнедеятельности и развитие высокой ме-таллоустойчивости растений.
Список литературы Динамика углевод - ферментного комплекса проростков ржи при адаптации к ионам свинца после теплового закаливания и без него
- Аликов Х.К. Фотоколориметрический метод определения содержания углерода в листьях мокрым сжиганием в хромовой смеси // Методы комплексного изучения фотосинтеза. Л., 1983. Вып. 2. С. 6-14.
- Большой практикум «Биохимия»: лабораторные работы / сост. М.Г. Кусакина, В.И. Суворов, Л.А. Чудинова. Пермь, 2012. С. 26-35.
- Даффус К., Даффус Дж. Углеводный обмен растений. М.: Агропромиздат, 1987. 150 с.
- Дерябин А.Н. и др. Влияние сахаров на развитие окислительного стресса, вызванного гипотермией (на примере растений картофеля, экспрес-сирующих ген инвертазы дрожжей) // Физиология растений. 2007. Т. 54, №1. С. 39^6.
- Карпец Ю.В., Колупаев Ю.Е. Ответ растений на гипертермию: молекулярно-клеточные аспекты // Вестник Харьковского национального аграрного университета. Сер. Биология. 2009. Вып. 1 (16). С. 19-38.
- Попов В.Н. Сравнительное исследование низкотемпературной адаптации томата и огурца в связи с их углеводным метаболизмом: дис. ... канд. биол. наук. М., 2002. 115 с.
- Сабельникова Е.П. Роль углеводного метаболизма в устойчивости растений к гипотермии на примере картофеля, трансформированного геном дрожжевой инвертазы: дис. ... канд. биол. наук. М., 2007. 137 с.
- Синькевич М.С. и др. Динамика активности инвер-таз и содержания сахаров при адаптации растений картофеля к гипотермии // Физиология растений. 2008. Т. 55, № 4. С. 501506.
- Титов А.Ф., Казнина Н.М., Таланова В.В. Тяжелые металлы и растения. Петрозаводск, 2014. 194 с.
- Титов А.Ф., Таланова В.В. Устойчивость растений и фитогормоны. Петрозаводск, 2009. 206 с.
- Трунова Т. И. Растение и низкотемпературный стресс. М.: Наука, 2007. 54 с.
- Холодова В.П. Компартментация сахаров в тканях растений // Рост растений. Первичные механизмы: сб. статей. М.: Наука, 1978. 289 с.
- Чудинова Л.А., Чернышева Ю.Д. Динамика содержания растворимых сахаров в проростках овса в условиях раздельного и комбинированного действия повышенных температур и ацетата кадмия // История и методология физиолого-биохимических и почвенных исследований: сб. науч. тр. Пермь, 2017. С. 48-51.
- Suzuki N., Koizumi N., Sano H. Screening in cadmium-responsive genes in Arabidopsis thaliana // Plant Cell Environ. 2001. Vol. 24, № 11. P. 1177-1188.
- Yang X. et al. Molecular mechanisms of heavy metal hyperaccumulation and phytoremediation // J. Trace Elem. Med. Biol. 2005. Vol. 18. P. 339353.
