Динамика зообентоса Северного Каспия в период подъёма уровня моря

Автор: Малиновская Любовь Васильевна, Зинченко Татьяна Дмитриевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 5-1 т.14, 2012 года.
Бесплатный доступ
Дается анализ данных многолетних исследований изменения численности и биомассы макрозообентоса Северного Каспия, отражающих основные тенденции количественного развития донных сообществ в период трансгрессии моря.
Макрозообентос, численность, биомасса, северный каспий, трансгрессия каспийского моря
Короткий адрес: https://sciup.org/148201442
IDR: 148201442 | УДК: 594.124
Zoobenthos dynamics of the North Caspian Sea at the sea level rise
In the article the analysis of long-term researches of change of macrozoobenthos number and biomass in the North Caspian Sea reflecting the basic tendencies of quantitative development of bottom-dwelling communities during the sea transgression is given.
Текст научной статьи Динамика зообентоса Северного Каспия в период подъёма уровня моря
П од влиянием различных факторов среды жизненные арены гидроэкосистем постоянно изменяются. Меняются экологические условия, а вместе с ними видовой и численный составы биоты планктонных и донных сообществ. И, чем масштабнее а, главное продолжительнее по срокам фактор изменчивости воздействует на природные экосистемы, тем бóльшие изменения он вызывает [2].
Так, исторически сложившиеся повышения и понижения уровня Каспия, контролируемые динамикой глобальных гидроклиматических процессов [17] приводят к перераспределению местообитаний гидробионтов разной толерантности по отношению к различным факторам воздействия. Результатом одной из смен фаз климата – сухой-теплой на влажную-прохладную явилась резкая трансгрессия Каспийского моря [43]. Динамичные преобразования гидролого-гидрохимических факторов, изменение биотопической структуры и биотических параметров характерны для различных районов Каспийского моря, но в значительной степени проявляются в его Северной части, в связи с чем этот район моря можно рассматривать как классическую модель для характеристики природопреобразовательных процессов. На этом основании Северный Каспий представляет собой уникальный полигон для исследований локальных и глобальных процессов динамики численности донных сообществ, изучение которых в многолетнем аспекте дает возможность показать динамику трофических условий и прогнозирования состояния промысловых рыб Каспия.
Период падения уровня Каспийского моря (1930-1977 гг.) сменился его подъёмом, который начался с 1978 г. Абсолютная отметка уровня Каспия достигла – 27,03 м. к 2002 г. Трансгрессия моря сказалась в первую очередь на экосистеме Северного Каспия, что явилось причиной значительного увеличения объема воды, возрастания, за счет за-
топления новых территорий, площади этой части моря, понижения солености, изменения гидрологического режима и степени накопления осадков [11, 12, 18, 49]. Указанные факторы в различной степени оказали влияние на процессы структурных преобразований донных сообществ, отдельных беспозвоночных, составляющих ядро кормовой базы рыб-бентофагов [27]. Вышесказанное позволяет констатировать, что анализ многолетних изменений количественного развития сообществ макрозообентоса в современный период трансгрессии экосистемы Каспия является актуальным и практически значимым. В публикациях ряда авторов показано, что под влиянием естественных и антропогенных причин, происходят значительные изменения (в большинстве случаев негативные) в развитии донных сообществ [21, 24, 29, 32, 35, 52, 53], а также популяций отдельных видов, в том числе и инвазионных [26, 37].
Целью работы является характеристика динамики развития зообентоса Северного Каспия в период трансгрессии моря на основе анализа многолетних исследований донных сообществ.
МАТЕРИАЛ И МЕТОДИКА
В работе использованы материалы мониторинговых съёмок (июнь) в северной части Каспийского моря в период с 1978 по 2002 гг., проводимых в Каспийском научно-исследовательском институте. Исследовали литоральную зону и глубоководную часть акватории северного Каспия с глубинами от 1,2 до 30 м.
Пробы бентоса отбирали дночерпателем «Океан-50», с площадью захвата 0,1 м². Содержимое дночерпателя промывали через капроновое сито из газа № 14 и фиксировали 4% - ным раствором формальдегида. Обработку материала в лаборатории проводили по общепринятой счетно-весовой методике (Романова, 1983).
В связи с тем, что в 1998 г. в Северном Каспии исследовалась исключительно литоральная зона (глубины менее 6,0 м), а в 1999 г. - акватория только западной части моря, то при осреднении количественных показателей зообентоса Северного Кас- пия, указанные годы не были включены в анализ данных.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Выделяются три периода повышения уровня Каспийского моря: 1978-1988 гг. (равномерный подъём уровня моря до средней отметки - 28,1 м БС); 1989 – 1995 гг. (резкий подъём уровня при средней отметке – 27,0 м БС); 1996 – 2002 гг.(стабилизация уровня моря, при средней отметке - 26,95 м БС), в течении которых анализируются изменения численности и биомассы макрозообентоса Северного Каспия.
Период начала трансгрессии моря (1978-1988 гг.) характеризовался возрастанием объёма весеннего половодья в среднем до 102,1 км³, уменьшением солености (средняя соленость составила 7,95 ‰) и увеличением площадей опресненных районов Северного Каспия. По данным [14], содержание в воде биогенных элементов и органических веществ в многолетнем ряду наблюдений было повышенным. Так, содержание минерального фосфора в водном стоке составило 7,51 тыс. т, что соответствовало уровню наиболее продуктивных лет периода естественной водности реки Волга, фосфора органического – 48,26 тыс. т, кремнекислоты – 458,9 тыс. т, органического вещества – 8,19 млн. т. Газовый и температурный режим Северного Каспия был благоприятным для развития донных животных. Было установлено, что средняя биомасса макрозообентоса составила 82,2 г/м², превышая в 1,5 раза биомассу бентоса, отмеченную в период «низкого уровня» моря. В отдельные годы этого периода биомасса бентоса варьировала от 54,8 (1980 г.) до 116,3 г/м² (1987 г.); средняя численность животных макрозообентоса изменялась от 23,3 тыс. экз./м² до 31,9 тыс. экз./м² (рис. 1). Валовая биомасса кормовых организмов бентоса составила 6,6 млн. т.
Высокие биомасса бентоса были обусловлены развитием моллюсков, доля которых составила до 80% от общей биомассы бентоса (рис. 2). Среднемноголетняя биомасса двустворчатых моллюсков в указанный период составила 63,9 г/м², изменяясь в разные годы от 36,0 (1980 г.) до 91,1 г/м² (1986 г.). Доминирующими среди моллюсков были представители «средиземноморского комплекса» Mytilaster lineatus (Gmelin) [7, 28], биомасса которых составила до 40% от общей биомассы бентоса и запаса моллюсков в Северном Каспии. В 1980 и 1982 гг. преобладающими по биомассе в бентосе становятся интродуцированные в море моллюски Abra ovata (Philippi). Наряду с массовым развитием представителей морской фауны, было установлено увеличение биомассы моллюсков, представителей солоноватоводного комплекса, таких как Didacna trigonoides (Pallas) , Adacna angusticostata (Borcea), составляющих до 30% от общей биомассы моллюсков в бентосе Северного Каспия.
Помимо моллюсков в составе так называемого «мягкого бентоса», биомасса олигохет и полихет (Annelidae) составила от 5,9 г/м² (1978 г.) до 14,2 г/м² (1987 г.) (рис. 2). Среди них доля олигохет была до 55% от общей биомассы ценоза, за исключением 1983 г., когда биомасса полихет Hediste diversicolor (Müller) и олигохет были сопоставимы.
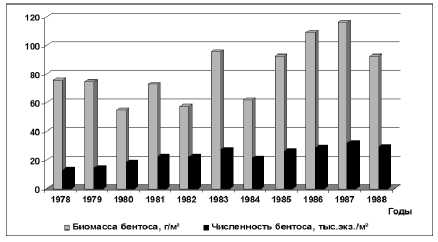
Рис. 1. Изменение численности и биомассы макрозообентоса Северного Каспия в 1978-1988 гг. (июнь)
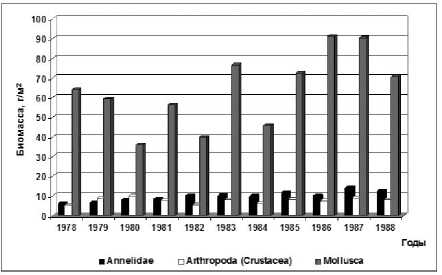
Рис. 2. Динамика биомассы массовых групп бентоса в донных сообществах Северного Каспия (1978-1988 гг., июнь)
Среднемноголетняя биомасса ракообразных (рис. 2) была достаточно высока – 7,6 г/м², изменяясь от 5,4 г/м² (1978 г.) до 9,9 г/м² (1980 г.). Коро-фииды составляли 60%, гаммариды – 30%, а доля кумовых раxков не превышала 10% от биомассы ракообразных. Отмечено, что в 1982 году биомасса кумовых могла достигать в отдельных районах до 47%.
Период с 1989 по 1995 гг. характеризовался резким подъёмом уровня моря. В 1995 г. средняя отметка уровня (-26,54 м БС) была близка к таковой в 1934 г., что привело к активизации динамических процессов в Северном Каспии, обусловив усиление переноса волжских вод в его восточную часть. Зарегистрировано увеличение водного стока во время половодья в среднем до 129,5 км³. Два экстремально многоводных года (1990, 1991 гг.) и ряд средневодных лет способствовали устойчивому снижению солености вод Северного Каспия в среднем до 6,09 ‰ [15], особенно в восточном районе и до 3,88 ‰ – в отдельных мелководных районах. В Северный Каспий с речным стоком поступило 6,32 млн. т органического вещества, что в 1,7 раза выше, чем в период «низкого» уровня моря [14]. Регистрировался возросший компенсационный подток средне- каспийских вод с низким содержанием кислорода, что стало одной из основных причин увеличения площадей Северного Каспия с дефицитом кислорода (начиная с 1989 г.) в придонном горизонте воды. В 1991-1995 гг. площади с насыщением воды кислородом менее 80% в Северном Каспии составляли 14,4 тыс. км², из которых 70% акватории приходилось на западную часть района [16]. В этих условиях среднемноголетняя биомасса зообентоса составила 75,6 г/м², что, примерно, соответствовало величинам развития бентоса в период равномерного подъема уровня воды.
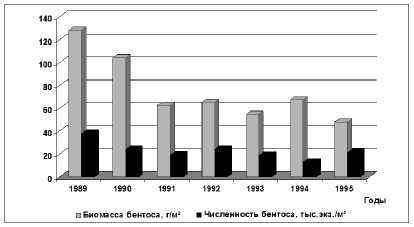
Рис. 3. Численность и биомасса макрозообентоса в Северном Каспии в 1989-1995 гг. (июнь)
Валовая биомасса донных животных составила 7,1 млн. т, увеличившись на 464,6 тыс. т. за счет развития олигохет, для которых было зарегистрировано расширение площадей обитания. Максимальная биомасса (128,2 г/м²) отмечена в маловодном 1989 г. (объем половодья – 97,0 км³), который характеризовался замедленным водообменом волжских и морских вод, увеличением продолжительности весеннего половодья. Соленость вод Северного Каспия в среднем была равна 7,27 ‰ [11]. Было отмечено массовое развитие средиземноморских моллюсков, которые обусловили высокую биомассу бентоса. Так, биомасса моллюска Mytilas-ter lineatus в донных сообществах Северного Каспия составила 41,9 г/м², а моллюска Abra ovata – 21,1 г/м².
Низкая биомасса (47,8 г/м²) донных беспозвоночных зарегистрирована в многоводном 1995 году (рис. 3) в условиях достаточно высокого весеннего половодья (объем водных масс – 136,8 км³) и снижения солености воды до 5,55 ‰ [11]. В сравнении с 1989 г. количественные показатели средиземноморских моллюсков Северного Каспия сократились в 10-16 раз, что и обусловило снижение общей биомассы бентоса.
Анализ состояния донных сообществ в период 1989-1995 гг. показал, что произошло увеличение биомассы организмов слабосолоноватоводного и солоноватоводного комплексов, среди которых преобладали моллюски Dreissena polymorpha poly-morpha (Pallas), D. p. andrusovi (Andrusovi) и Adacna angusticostata. Характерно, что биомасса моллюска Adacna angusticostata достигла в 1992 г. максимальной за весь период исследований величины – 17,8 г/м². В тоже время, начиная с 1991 г.
резко снизились численность и биомасса морской фауны, а именно моллюсков – средиземноморских вселенцев Mytilaster lineatus и Cerastoderma la-marcki (Reeve), что обусловлено изменением режима весеннего половодья. Свободные трофические ниши Северного Каспия были заняты моллюском Abra ovata , имеющим конкурентные преимущества, связанные с устойчивостью моллюсков к дефициту кислорода.
Общая биомасса моллюсков колебалась от 31,8 (1995 г.) до 105,6 г/м² (1989 г.), составив в среднем за период 57,3 г/м² (рис. 4).
В 1988-1995 гг. при начавшемся процессе эв-трофирования, характерного для зоны устьевого волжского пространства Северного Каспия, в донных сообществах заиленных серых и черных илов стали преобладать Oligochaeta [16]. Численность олигохет на заиленных биотопах в отдельные годы достигала 15,5 тыс. экз./м².
Количественные показатели ракообразных, среди которых, по-прежнему, преобладали корофии-ды, были на 20% ниже, чем в период равномерного подъёма уровня моря (рис. 4).
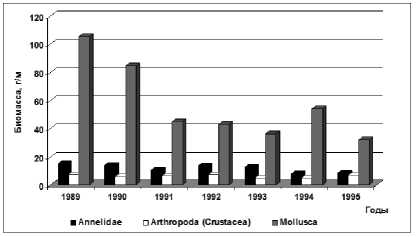
Рис. 4. Динамика биомассы отдельных таксонов зообентоса в Северном Каспии в 1989-1995 гг.
В период стабилизации уровня моря (1996-2002 гг.) водный сток за период половодья составил 108 км³, изменяясь от 61,6 км³ (1996 г.) до 133,7 км³ (2001 г.). Начиная с 1998 г. площади акватории моря с низким содержанием кислорода (менее 80%), в результате изменения гидродинамики водных масс, сократились. В 2001-2002 гг. в северной части Каспийского моря регистрировался благоприятный кислородный режим для обитания гидробионтов [16, 10]. Было установлено возрастание транзитного поступления биогенных веществ и постепенная стабилизация биогенного режима и продуцирования органического вещества [11, 16]. В западной части северного Каспия на обширной его акватории возросло содержание органического углерода; преобладающим субстратом стал заиленный песок [49], что обусловило качественную перестройку таксономической структуры бентоса [25]. Начиная с 1998 г., при увеличении температуры воды до 29-30ºС, регистрируется антропогенное загрязнение вод поллютантами различной природы, источником которых являются бытовые и промышленные стоки, а также техногенное загрязнение, связанное с разведкой и добычей углеводородного сырья [9,
-
14, 20, 36, 38, 39]. Средняя биомасса макрозообентоса в этот период снизилась более чем в 2 раза по отношению к периоду резкого подъёма уровня моря и составила 35,5 г/м² с колебаниями в различные годы от 27,4 г/м² (1996 г. – экстремально маловодный) до 50,0 г/м² (2000 г. – средневодный). Численность донных животных сократилась примерно в два раза (рис. 5). Анализируя межгодовую динамику бентоса, можно отметить, что невысокие величины численности и биомассы бентоса в 1996 г., обусловлены, вероятно, экстремально низкой водностью (объем весеннего половодья составил 61,6 км³, в сравнении с 136,8 км³ в 1995 г.) и возрастанием солености, которая, например в июне 1996 г. была 7,78 ‰, тогда как в 1995 году не превышала 5,55 ‰. [15]. Хотя основу биомассы зообентоса формировали по-прежнему моллюски (до 70%), их биомасса, в сравнении с данными за 1989-1995 гг. сократилась в 2 раза, а численность – более чем в 5 раз (рис. 6). Доминирующей по биомассе в течение всего периода была каспийская монодакна ( Adacna angusticostata ), за исключением 2000 г., когда в составе бентоса преобладали моллюски M. lineatus .
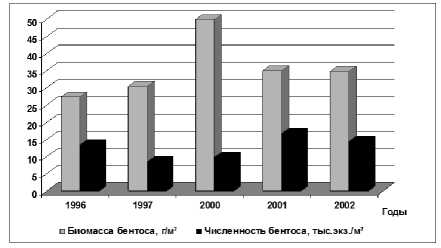
Рис. 5. Численность и биомасса зообентоса Северного Каспия в 1996-2002 гг. (июнь)
Помимо снижения количественных показателей двустворчатых моллюсков, произошло перераспределение их размерного состава. В зообентосе, начиная с 2001 г. стали преобладать моллюски крупных размеров (более 15 мм) и значительно сократилась доля молоди, что особенно характерно для галофильных видов. Так, у солоноватоводных моллюсков – A. angusticostata особи менее 5 мм в период с 1990 по 2000 гг. составляли 69,6% от общей численности популяции, тогда как в 2002 г. молодь моллюсков составила 9,4 %. У других видов моллюсков, например, D. trigonoides отмечено сходное соотношение крупных и мелких особей (19 и 7,2%, соответственно), у M. lineatus – 81 и 50,3%, у A. ovata – 58 и 25.8% соответственно.
Наши данные соответствуют результатам изменения структуры зоопланктонных сообществ. Так, в июне 2001 и 2002 гг. численность велигеров моллюсков в планктоне Северного Каспия сократилось более чем в 4,5 раза [1]. По литературным сведениям, одной из основных причин снижения доли молоди двустворчатых моллюсков в составе донных сообществ, является расселение с 2000 г. в водах Каспийского моря гребневика – Mnemiopsis leidyi
Agassiz , вселение которого нарушило функционирование всех звеньев трофической сети экосистемы Каспия [50, 54]. Известно, что у большинства моллюсков личинки встречаются в водоеме с апреля по октябрь. Появление гребневика в водах Северного Каспия регистрируется в начале июля, после чего его численность достигает максимума в конце лета [8], в период пика размножения галофильных моллюсков M. lineatus , A. ovata , C. lamarcki [5, 31]. В спектр питания гребневика, наряду с зоопланктоном, входят личинки брюхоногих и двустворчатых моллюсков [44], что в конечном итоге приводит к сокращению популяций моллюсков, в основном, представителей морского комплекса и, как следствие, к снижению доли оседающих на субстрат личинок.
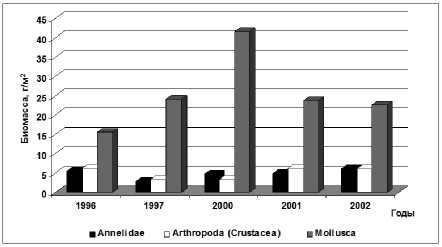
Рис. 6. Изменение биомассы отдельных групп зообентоса Северного Каспия в годы исследования периода с 1996 по 2002 гг.
Подтверждением влияния гребневика на изменение возрастной структуры галофильных моллюсков является и то, что в составе, например, слабосолоноватоводных моллюсков в 2001-2002 гг., несмотря на общее снижение численности популяций, по-прежнему, преобладала молодь. Так, в составе популяции моллюска Adacna glabra ( A. vitrea ) особи менее 3 мм занимали в 1990-2000 гг. 89,1%, в 2002 г. – 97,0%. Этому способствует и тот факт, что размножение моллюска происходит весной, то есть в период отсутствия гребневика в Северном Каспии [46]. Кроме этого, вселившийся гребневик Mnemiopsis leidyi относится к представителям морского комплекса и в Северном Каспии имеет область распространения в водах с соленостью превышающей 4‰. Слабосолоноватоводные гидробионты, обитающие за пределами этой границы, оказываются более устойчивыми к воздействию гребневика и по сути находятся вне зоны конкурентных взаимоотношений с M. leidyi , нежели морские виды моллюсков и других гидробионтов.
Следующей причиной сокращения количественных показателей моллюсков, вероятно, следует считать возросшее антропогенное воздействие. По данным [9], основным загрязняющим компонентом вод Северного Каспия в 1998 г. являлись нефтеуг-леводороды, содержание которых в среднем увеличилось в 4,3 раза в сравнении с 1997 г. и было в 1,2 раза выше, чем в речной воде, а в районах, распо- ложенных в северо-западной части Северного Каспия и прилегающих к взморью р. Урал превышало 20-25 ПДК. Уровень содержания фенолов в воде Северного Каспия также возрос и в среднем составил 3,6 ПДК. Установлено, что появление в воде токсических веществ самой разнообразной природы, даже при сравнительно небольших концентрациях, вызывает у донных животных нарушения физиологических и биохимических процессов, в том числе процесса воспроизводства [3, 6]. Есть сведения о том, что в результате длительного воздействия токсикантов или при их высоких концентрациях некоторые виды моллюсков элиминируют, другие мигрируют, третьи приспосабливаются к условиям загрязнения [42].
Колебания солености и температурного режима Северного Каспия также могут негативно влиять на процесс размножения моллюсков. Так, распресне-ние водоема у одних видов моллюсков приводит к увеличению пелагических личинок, у других - к подавлению гаметогенеза или задержке сроков нереста [5]. Опасность внезапного срыва в воспроизводстве заключает в себе и изменение (повышение) специфичной для каждого вида оптимальной температуры воды, определяющей сроки нереста [4].
Что касается развития ракообразных, то в период стабилизации уровня моря их биомасса на превышала 5,5 г/м2 (рис. 6). Минимальная биомасса ракообразных (2,2 г/м²) зарегистрирована в западном районе Северного Каспия в 1999 г. и является самой низкой за 46-летний период (1953-2002 гг.) исследований бентоса в Северном Каспии. Одним из факторов, влияющих на динамику ракообразных, возможно, следует считать возросшее с 1998 г. загрязнение Северного Каспия. Ракообразные, в отличие от моллюсков, быстрее реагируют на все процессы, протекающие в водоеме. Особенно уязвимыми придонные ракообразные становятся в момент линьки [40]. Вероятно, реакцией на изменение состояние экосистемы Северного Каспия в 1998 г., стало резкое снижение численности и биомассы ракообразных уже в 1999 г. В 2001 г. биомасса Crustacea увеличилась (рис. 6) за счет доминирования в различные годы гаммарид и корофиид, которые стали повсеместно составлять основу численности зообентоса, достигая на отдельных участках до 90 тыс. экз./м². Помимо изменяющихся условий среды, фактором, обуславливающим колебания численности и биомассы ракообразных, является их выедание рыбами бентофагами [19]. Биомасса ракообразных в период стабилизации уровня моря, в сравнении с предыдущим периодом, сократилась примерно в два раза.
Биомасса олигохет и полихет составила 4,8 г/м², что в 2,5 раза ниже средней многолетней величины и биомассы периода резкого подъема уровня моря. Доминирующей группой среди них были малощетинковые черви, на долю которых приходилось 52%. Отмечается, что в 1997 и 2002 гг. биомассы Hediste diversicolor и олигохет были сопоставимы.
В анализируемый период преобладала численность полихеты Hypaniola kowalewskii . Вместе с тем, снижение количественных показателей червей в донных сообществах Северного Каспия, среди которых численность олигохет уменьшилась в три раза, связано, по-видимому, с возможным недоучетом их численности вследствие сокращения в последние годы исследований мелководных участков (с глубиной менее 3,0 м) Северного Каспия, где малощетинковые черви образуют основные поселения.
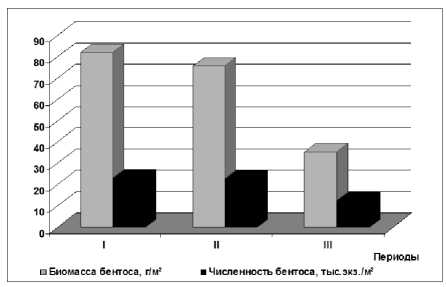
Рис. 7. Изменение численности и биомассы макрозообентоса Северного Каспия в различные периоды: I – 1978 - 1988 гг. – равномерный подъем уровня моря; II – 1989 - 1995 гг. – резкий подъем уровня моря; III – 1996 - 2002 гг. – стабилизация уровня моря (представлены данные за июнь)
Таким образом, динамичные преобразования экосистемы Каспийского моря в периоды его трансгрессии привели к коренным изменениям структурных характеристик пелагических и донных сообществ, некоторые результаты которых представлены в данной работе и ранее освещались в публикациях [25, 35, 52, 53] и др. Можно констатировать, что донные сообщества интенсивней развивались в период повышения уровня моря, нежели в годы с низким его уровнем. Подводя итог выше сказанному, на основании анализа динамики численности и биомассы макрозообентоса в различные периоды изменения уровня моря, установлено снижение численности и биомассы лидирующих ценозов донных организмов в период стабилизации уровня моря, в сравнении с периодом равномерного подъема уровня Каспийского моря (рис. 7).
К основным факторам, обусловившим структурные изменения донных сообществ, следует отнести величину объема половодья, динамику уровня моря, режима солености, трофические условия и антропогенное воздействие, в том числе техногенное и биогенное загрязнение моря, вызванное инвазией гребневика Mnemiopsis leidyi. Межгодовые колебания биомассы зообентоса, наблюдаемые в отдельные периоды, в значительной степени были вызваны изменением численности и биомассы ви-дов-вселенцев – Abra ovata, Hediste diversicolor и Mytilaster lineatus, которые в отдельные годы со- ставляли до 70% от общей биомассы донных беспозвоночных, что подтверждается исследованиями ряда авторов [22-24, 30, 33, 34, 37, 45, 47, 48, 51].
Для выяснения современных тенденций и многолетней динамики количественных характеристик бентосных сообществ мы ограничились анализом экосистемы Северного Каспия, где сконцентрированы комплексы разнотипных по своему происхождению гидробионтов и отчетливо видны их изменения в период трансгрессии моря. Это позволило выявить некоторые закономерности их развития в условиях коренной перестройки гидрологогидрохимического режима водоема. Полученные данные могут быть использованы при прогнозировании состояния кормовой базы в условиях разнотипных воздействий на экосистему моря.
Список литературы Динамика зообентоса Северного Каспия в период подъёма уровня моря
- Ардабьева А.А., Тарасова Л.И., Малиновская Л.В., Смирнова Л.В. Кормовая база Северного Каспия в 2003 г.//Рыбохозяйственные исследования на Каспии. Результаты НИР за 2003 г. Астрахань, 2004. С. 112-123.
- Вилков Е.В. Тренды численности регулярно мигрирующих куликов на западном побережье Среднего Каспия//Кулики Северной Евразии: экология миграции и охрана. Тез. докл. VIII Междунар. науч. конф. Ростов-на-Дону: Изд-во ЮНЦ РАН, 2009. С. 29-30.
- Галина М.С., Петров С.А. Моллюски в загрязненном море//Химия и жизнь. 1998. № 6. С. 52-53.
- Гальперина Г.Е. К вопросу о воспроизводстве кормовой базы рыб Северного Каспия (моллюски)//Тезисы отчетной сессии ЦНИОРХа, Астрахань. 1972. С. 37-38.
- Гальперина Г.Е. Размножение двустворчатых моллюсков (Bivalvia) Северного Каспия: Автореф. дис… канд. биол. наук. М.: ИОАН, 1976. 29 с.
- Гераскин П.П., Металлов Г.Ф., Аксенов В.П., Дубовская А.В., Файзулина Д.Р., Галактионова М.Л. Использование биологических индикаторов для оценки загрязненности морской среды в районах расположения ликвидированных поисково-разведочных скважин//Защита окружающей среды в нефтегазовом комплексе. 2009. № 9. С. 55-58.
- Зенкевич Л.А. Фауна и биологическая продуктивность моря. М.: Советская наука, 1951. Т. 1. 506 с.
- Камакин А.М., Студеникина Ю.Б. Распространение вселенца Mnemiopsis leidyi в Каспийском море//Рыбохозяйственная наука на Каспии: задачи и перспективы. Астрахань: Изд-во КаспНИРХ, 2003. С. 81-84.
- Катунин Д.Н., Гераскин П.П. Рыба и нефть Каспийского моря//Рыбное хозяйство. № 4. 1996. С. 46-47.
- Катунин Д.Н., Хрипунов И.А. Многолетняя динамика формирования зон гипоксии в Северном Каспии в районах обустройства нефтегазовых месторождений//Проблемы сохранения экосистемы Каспия в условиях освоения нефтегазовых месторождений. Астрахань: Изд-во КаспНИРХ, 2005. С. 98-100.
- Катунин Д.Н., Беспорточный Н.П., Хрипунов И.А. Особенности гидролого-гидрохимического режима Каспийского моря//Научные основы устойчивого рыболовства и регионального распределения промысловых объектов Каспийского моря. М.: Изд-во ВНИРО, 1998. С. 9-25.
- Катунин Д.Н., Курочкина Т.Ф., Насибулина Б.М., Попова О.В., Карьгина Н.В. Эколого-токсикологическая характеристика Волго-Каспийского бассейна в современных условиях//Рыбохозяйственные исследования на Каспии: Результаты НИР за 1998. Астрахань: КаспНИРХ, 1999. С. 26-29.
- Катунин Д.Н., Галушкина Н.В., Железцова Е.Г., Радованов Г.В. Многолетние (1936-1999) изменения биогенного стока р. Волги//Рыбохозяйственные исследования на Каспии: Результаты НИР за 1999 г. Астрахань: КаспНИРХ, 2000. С. 362-368.
- Катунин Д.Н., Хрипунов И.А., Беспорточный Н.П., Галушкина Н.В., Никотина Л.Н., Кравченко Е.А., Родованов Г.В., Дулимов А.Б. Гидролого-гидрохимический режим дельты Волги и Каспийского моря в 2000 г.//Рыбохозяйственные исследования на Каспии: Результаты НИР за 2000 г. Астрахань: КаспНИРХ, 2001. С. 52-69.
- Катунин Д.Н., Егоров С.Н., Кашин Д.В., Хрипунов И.А., Галушкина Н.В., Кравченко Е.А., Гуляев В.Ю., Дектярева О.А. Основные черты гидролого-гидрохимического режима нижнего течения р. Волги и Каспийского моря//Вопросы промысловой океанологии. М.: Изд-во ВНИРО, 2004. Вып. 1. С. 69-96.
- Катунин Д.Н., Егоров С.Н., Хрипунов И.А., Кашин Д.В., Галушкина Н.В., Кравченко Е.А. Основные особенности гидролого-гидрохимического режима р. Волга и Каспийского моря в трансгрессивный период//Рыбное хозяйство, 2007. № 3. С. 75-79.
- Кривенко В.Г. Водоплавающие птицы и их охрана. М.: Агропромиздат, 1991. 273 с.
- Кураев А.В. Особенности гидрологических условий Северного Каспия в период современного повышения уровня моря: Автореф. дис… канд. биол. наук. Москва, 1998. 20 с.
- Кушнаренко А.И. Распределение и численность полупроходных рыб в Северном Каспии//Рыбохозяйственные исследования на Каспии: результаты НИР за 2001 г. Астрахань: КаспНИРХ, 2002. С. 247-263.
- Макарова Е.Н., Монахов С.К., Гаврилова Е.В., Петреченкова В.Г., Холина О.И. Роль Волго-Каспийского канала в транзите загрязняющих веществ из Волги в Северный Каспий//Защита окружающей среды в нефтегазовом комплексе. 2009. № 9. С. 55-58.
- Малиновская Л.В. Особенности развития зообентоса Северного Каспия в условиях продолжающегося подъема уровня моря//Тез. докл. Х Междунар. конф. по промысловой океанологии. М.: ВНИРО, 1997. С. 82-83.
- Малиновская Л.В. Роль вселенцев в формировании донной фауны Северного Каспия//Виды -вселенцы в Европейских морях России. Мурманск, 2000. С. 57-59.
- Малиновская Л.В. Развитие средиземноморских вселенцев Abra ovata и Mytilaster lineatus в Северном Каспии в современный период//Эволюция морских экосистем под влиянием вселенцев и искусственной смертности фауны. Ростов-на-Дону, 2003. С. 106.
- Малиновская Л.В. Зообентос Северного Каспия в период подъема уровня моря: Атореф. дис.. канд. биол. наук. Тольятти, 2007. 20 с.
- Малиновская Л.В. Состояние макрозообентоса на лицензионном участке ООО «КНК» в летний период 2004-2006 гг.//Материалы II Междунар. научно-практич. конф. «Проблемы сохранения Экосистемы Каспийского моря в условиях освоения нефтегазовых месторождений». Астрахань: Изд-во КаспНИРХ, 2007 б. С. 71-75.
- Малиновская Л.В., Зинченко Т.Д. Mytilaster lineatus (Gmelin): многолетняя динамика, распределение инвазионного моллюска в Северном Каспии//Российский журнал биологических инвазий (ISSN 1996-1499. www. sevin.ru). 2009, №1. С. 7-17.
- Малиновская Л.В., Зинченко Т.Д. Многолетняя динамика сообществ макрозообентоса Северного Каспия//Изв. СамНЦ РАН. 2010. Т.12, №.1. С. 179-184.
- Модухай-Болтовской Ф.Д. Каспийская фауна в Азово-Черноморском бассейне. М. Л.: Изд-во АН СССР, 1960. 298 с.
- Осадчих В.Ф. Бентос Северного Каспия в условиях зарегулирования стока р. Волги//Зоол. журн. Т. 42, вып. 2. 1963а. С. 184-196.
- Осадчих В.Ф. Роль вселенцев в бентосе Северного Каспия//Зоол. журн. 1963 б. Т. 42. Вып. 7. С. 90 -104.
- Осадчих В.Ф. Сезонная динамика северокаспийских двустворчатых моллюсков//Тр. КаспНИРХ, 1967. Т. 23. С. 80-90.
- Осадчих В.Ф. Изменение биомассы бентоса В Северном Каспии за последнее пятилетие//Тр. КаспНИРХ, 1968. Т. 24. С. 100-112.
- Осадчих В.Ф. Роль отдельных видов в биомассе бентоса Северного и Среднего Каспия//Зоол. журн. 1978. Т. 57. Вып. 1. С. 26-31.
- Осадчих В.Ф., Романова Н.Н. Современное состояние зообентоса Каспийского моря//Вопросы гидробиологии. I съезд ВГБО. Тез. докладов. М.: Наука, 1965. С. 323-324.
- Осадчих В.Ф., Ардабьева А.Г., Белова Л.Н., Елизаренко М.М., Курашова Е.К., Малиновская Л.В., Попова М.К., Степанова Т.Г., Тиненкова Д.Х., Татаринцева Т.А. Особенности развития и использования кормовой базы рыбами в условиях повышения уровня Каспийского моря//Комплексные рыбохозяйственные исследования на Каспии. М.: Изд-во ВНИРО, 1989. С. 119-137.
- Панин Г.Н., Мамедов Р.М., Митрофанов И.В. Современное состояние Каспийского моря/отв. ред. М.Г. Хубларян//Ин-т Водных проблем РАН. М.: Наука, 2005. 356 с.
- Полянинова А.А. Виды вселенцы в Каспии и их роль в экосистеме моря. Астрахань: КаспНИРХ, 2007. 104 с.
- Попова О.В., Попова Э.С. Многолетняя динамика выноса фенолов в составе волжского стока и распределение по акватории Северного Каспия // Современные проблемы водной токсикологии // Тез. Междунар. конф. Борок: ИБВВ РАН. 2005. С. 109.
- Попова О.В., Чуйко Е.В. Многолетнее изменение содержания тяжелых металлов в поверхностных водах Северного Каспия и факторы их определяющие//Тез. Междунар. конф. Борок: ИБВВ РАН. С. 110.
- Романова Н.Н. Экология и количественное распределение автохтонных гаммарид Каспийского моря//Тр. ВНИРО. 1973. Т. 80. Вып. 3. С. 73-103.
- Романова Н.Н. Методические указания к изучению бентоса южных морей СССР. М.: ВНИРО, 1983. 13 с.
- Рылова Н.Е., Райская Г.Ю. О роли гидробионтов в экологическом мониторинге поверхностных и техногенных вод территории Северного Каспия//Материалы Междунар. конф., посвященной 90-летнему юбилею АзНИИРХ. Современные проблемы биологических ресурсов Каспийского моря. Астрахань: Изд-во КаспНИРх, 2003. С. 75-78.
- Свиточ А.А. Геоэкологическая катастрофа в приморских городах Дагестана//Природа. 1998. Т. 5 (993). С. 16-17.
- Сергеева Н.Г., Буркацкий О.Н. Макрозообентос восточной части Азовского моря в осенний период//Экология моря. Севастополь. 2002. Вып. 61. С. 29-35.
- Смирнова Л.В. Состояние зообентоса Северного Каспия в конце 80-х годов//Морские гидробиологические исследования. М.: ВНИРО, 2000. С. 103-110.
- Спасский Н.Н. Жизненный цикл моллюска Adacna vitrea и сезонная динамика его биомассы и количества//Тр. КаспНИРХ. 1967. Т. 23, С. 91-94.
- Стыгар В.М., Мутышева Г.К. Зообентос восточной части Северного Каспия в зоне предполагаемой разработки месторождений углеводородного сырья в период стабилизации уровня моря//Морские гидроб. исследования: Сб. науч. трудов. М.: ВНИРО, 2000. С. 111-126.
- Татаринцева Т.А., Малиновская Л.В., Терлецкая О.В., Ардабьева А.Г. Средиземноморские вселенцы в планктоне и донной фауне Каспийского моря//Виды-вселенцы в Европейских морях России, Апатиты, 2000. С. 169-184.
- Хрипунов И.А., Азаренко А.В. Органическое вещество донных осадков Северного Каспия//Рыбохозяйственные исследования на Каспии: результаты НИР за 2004 г. Астрахань. 2005. С. 66-69.
- Шиганова Т.А., Камакин А.М., Жукова О.П., Ушивцев В.Б., Дулимов А.Б., Мусаева Э.И. Вселенец в Каспийское море и первые результаты его воздействия на пелагическую систему//Океанология. 2001. Т. 41. Вып. 4. С. 242-249.
- Яблонская Е.А., Осадчих В.Ф. Изменение кормовой базы бентосоядных рыб Северного Каспия//Тр. ВНИРО. Т. 80. 1973. С. 48-73
- Яблонская Е.А., Осадчих В.Ф. Влияние океанологических условий на формирование бентоса//Гидрометеорология и гидрохимия морей. Каспийское море. СПб.: «Гидрометеоиздат». 1996. Т. VI. Вып. 2. С. 263-278.
- Malinovskaja L.V., Filippov A.A., Osadchikh V.F., Aladin N.V. Benthic macroinvertebrates of the Northern Caspian Sea during recent rises in water -level//Intern. Journ. of Salt Lake Research. Vol. 7. 1998. P. 211-233.
- Shiganova T.A., Dumont H.J., Sokolsky A.F., Kamakin A.M. et al. Pobulation dynamics of Mnemiopsis leidy in the Caspian and Mediterranean Seas // Eds. Subramanian B.K., Ramadhan V., Venugopolan V.L. Nannoplankton production in Vellar Estuary // Curr. Sci (India), 2004.Vol. 46, № 7. P. 212-215.
